Alternative Oxidase: A Mitochondrial Respiratory Pathway to Maintain Metabolic and Signaling Homeostasis during Abiotic and Biotic Stress in Plants

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
1.1. Plant Respiration and the Plant Mitochondrial Electron Transport Chain
1.2. Respiratory Metabolism and Plant Stress Biology
1.3. Mitochondrial Reactive Oxygen and Reactive Nitrogen Species
1.4. The Plant Mitochondrial Stress Response
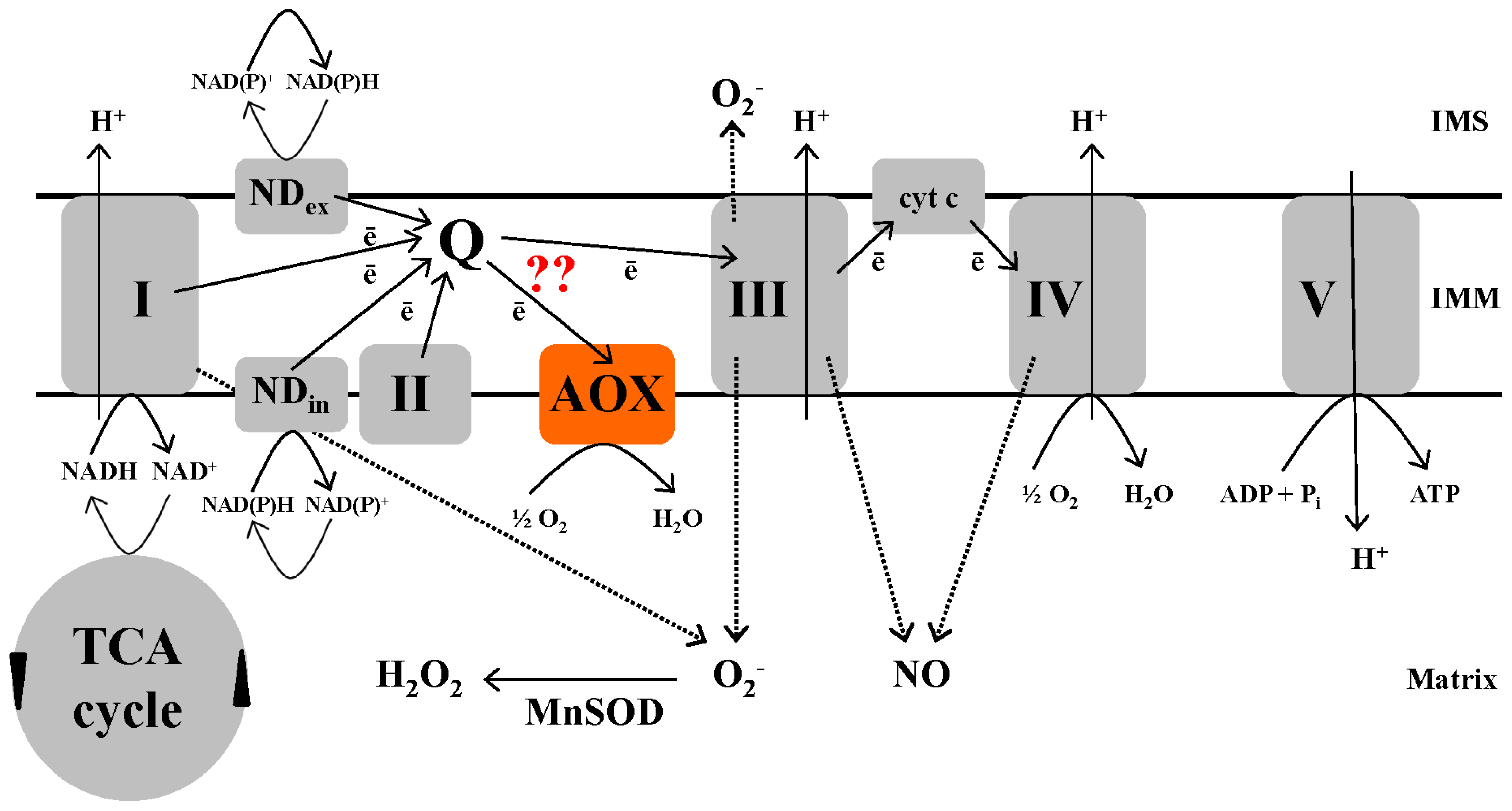
2. Alternative Oxidase
2.1. Brief Background
2.2. AOX Capacity and Activity
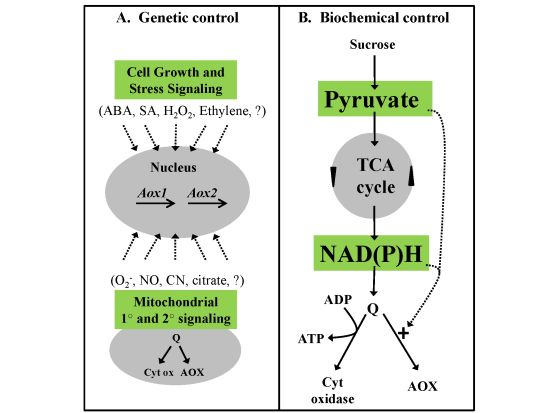
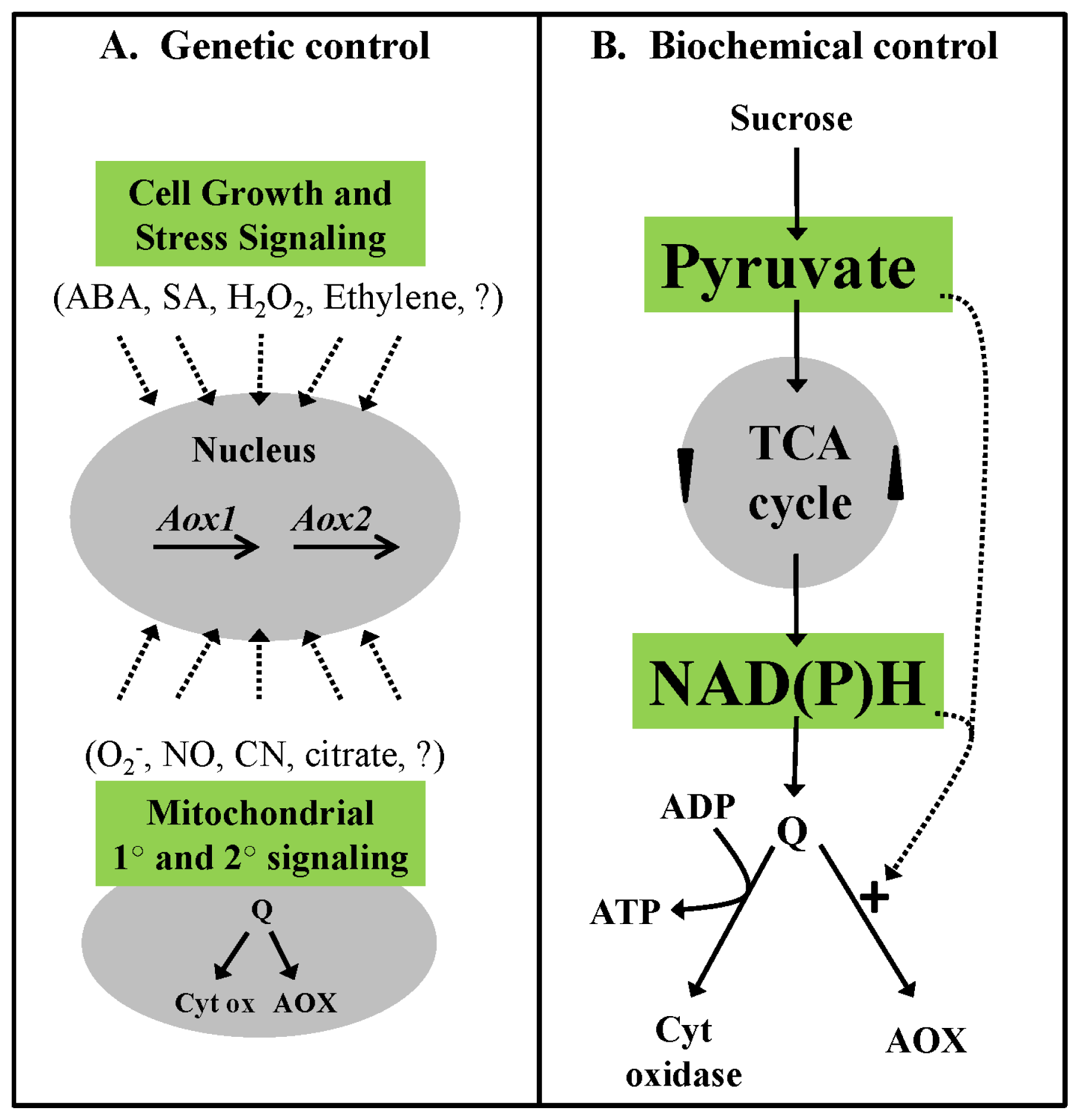
2.3. Genetic Control of AOX Respiration
2.4. Biochemical Control of AOX Respiration
2.5. Control of Mitochondrial ROS and RNS Generation by AOX
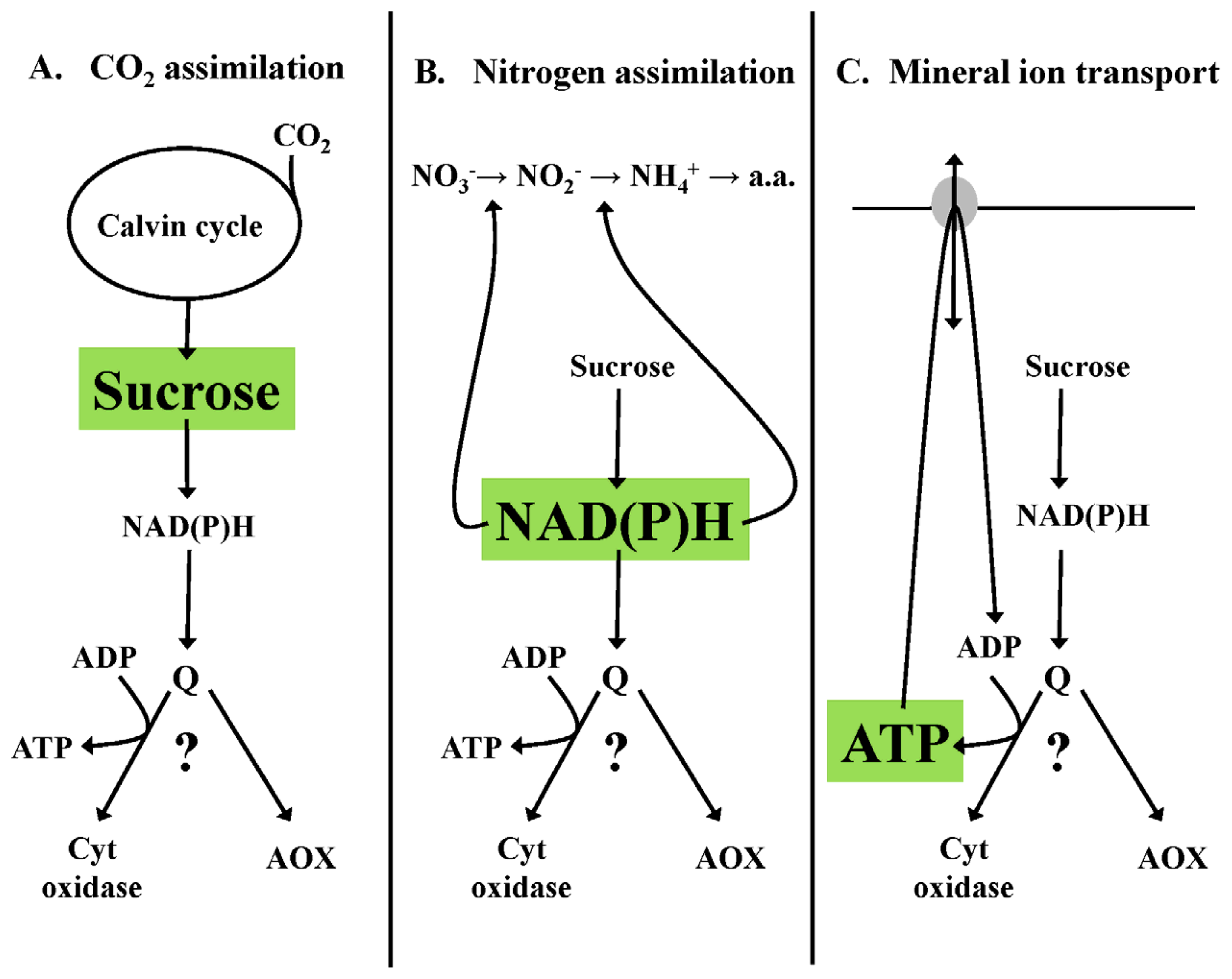
3. A Role for Alternative Oxidase to Maintain Metabolic Homeostasis
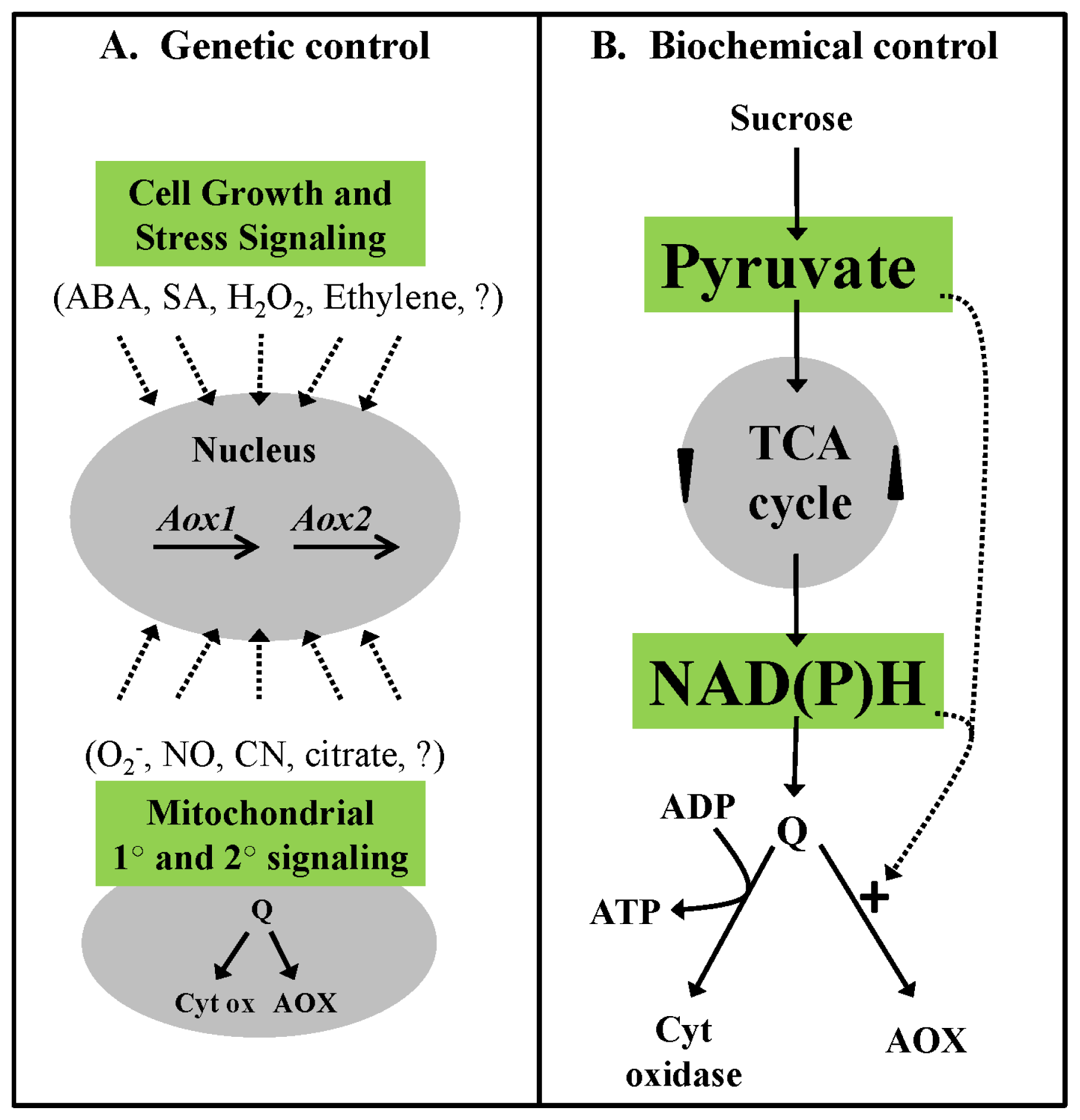
3.1. Homeostasis of Carbon Pools—An Example
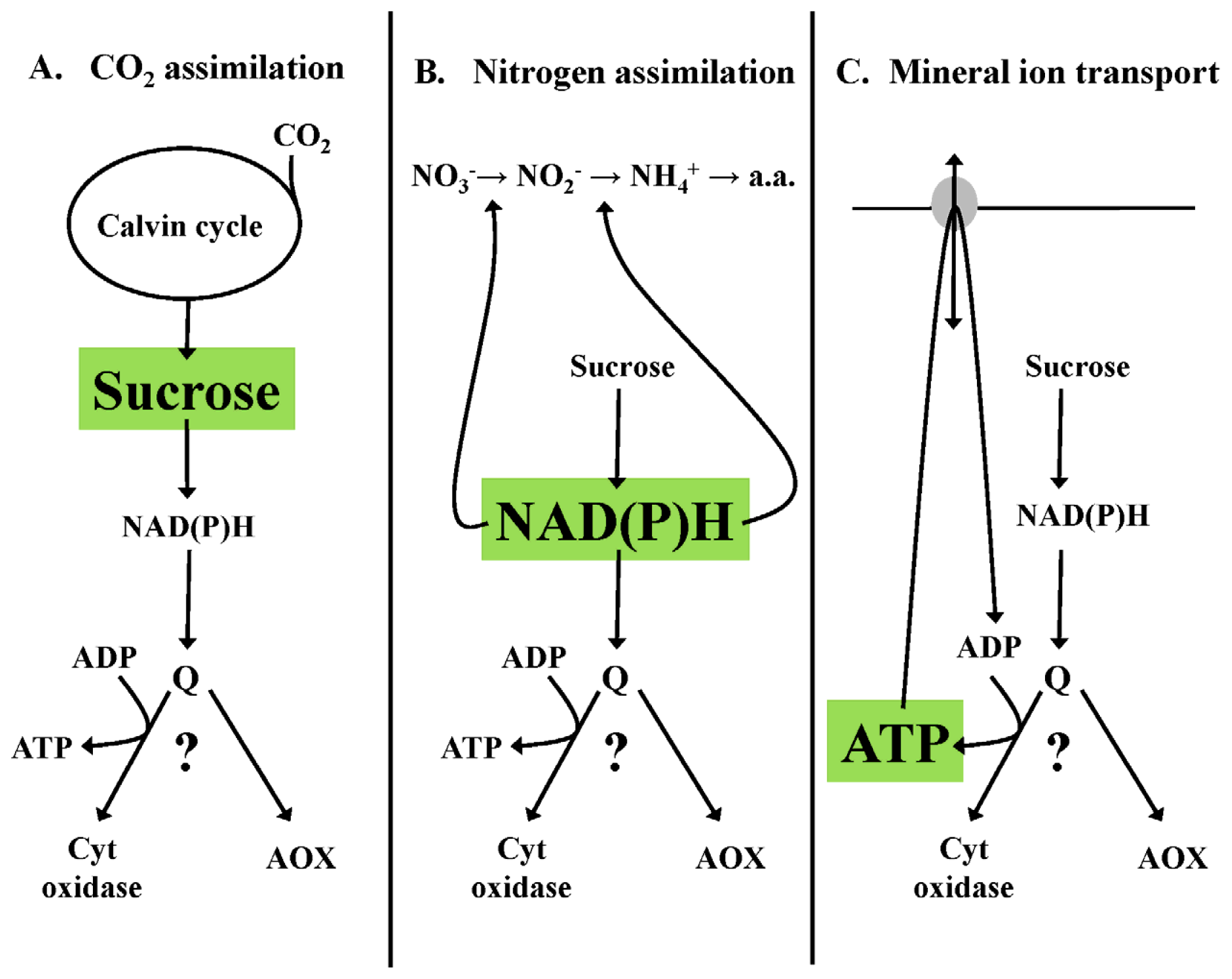
3.2. Homeostasis of Redox State—An Example
3.3. Homeostasis of Energy Status—An Example
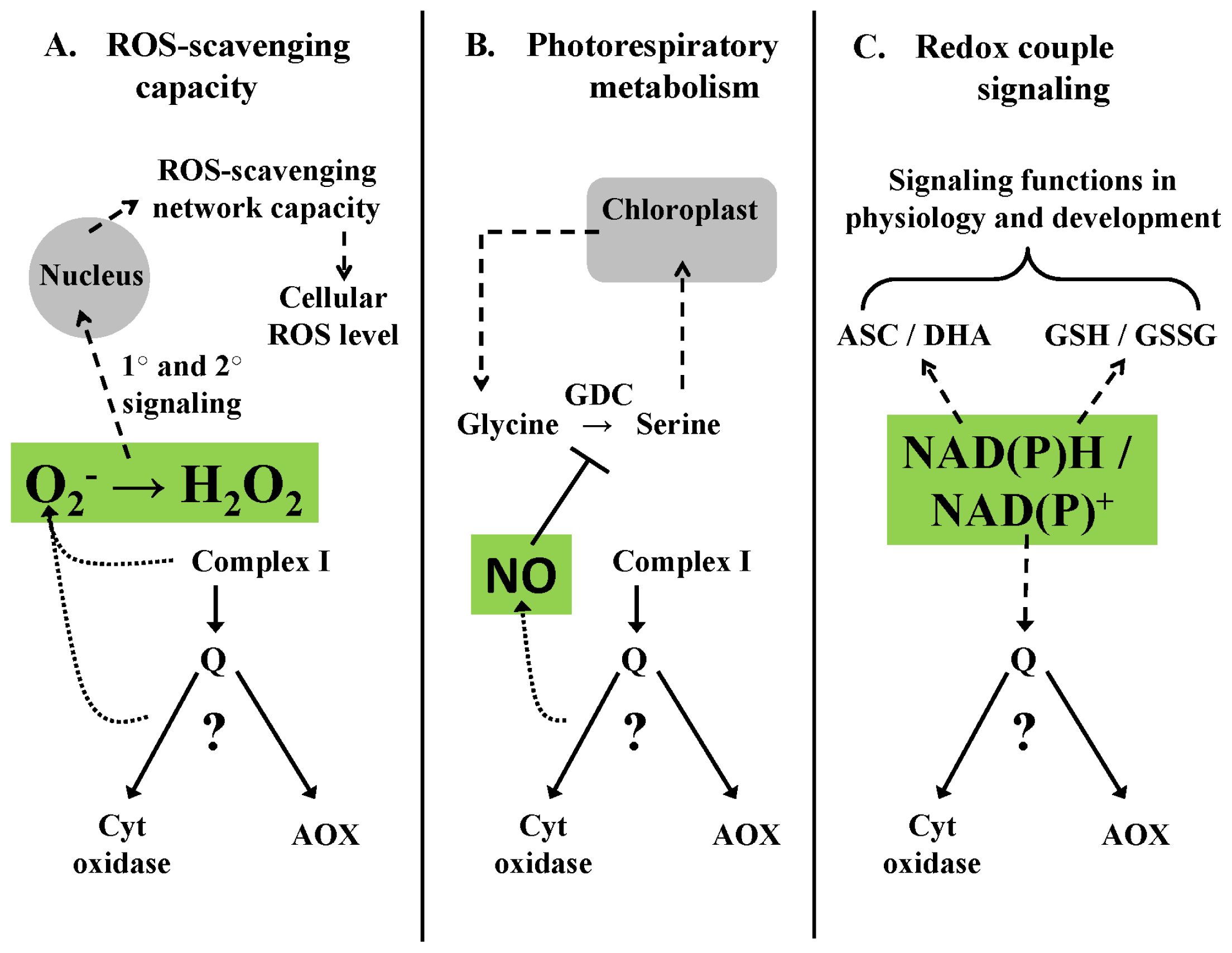
4. A Role for Alternative Oxidase to Maintain Signaling Homeostasis
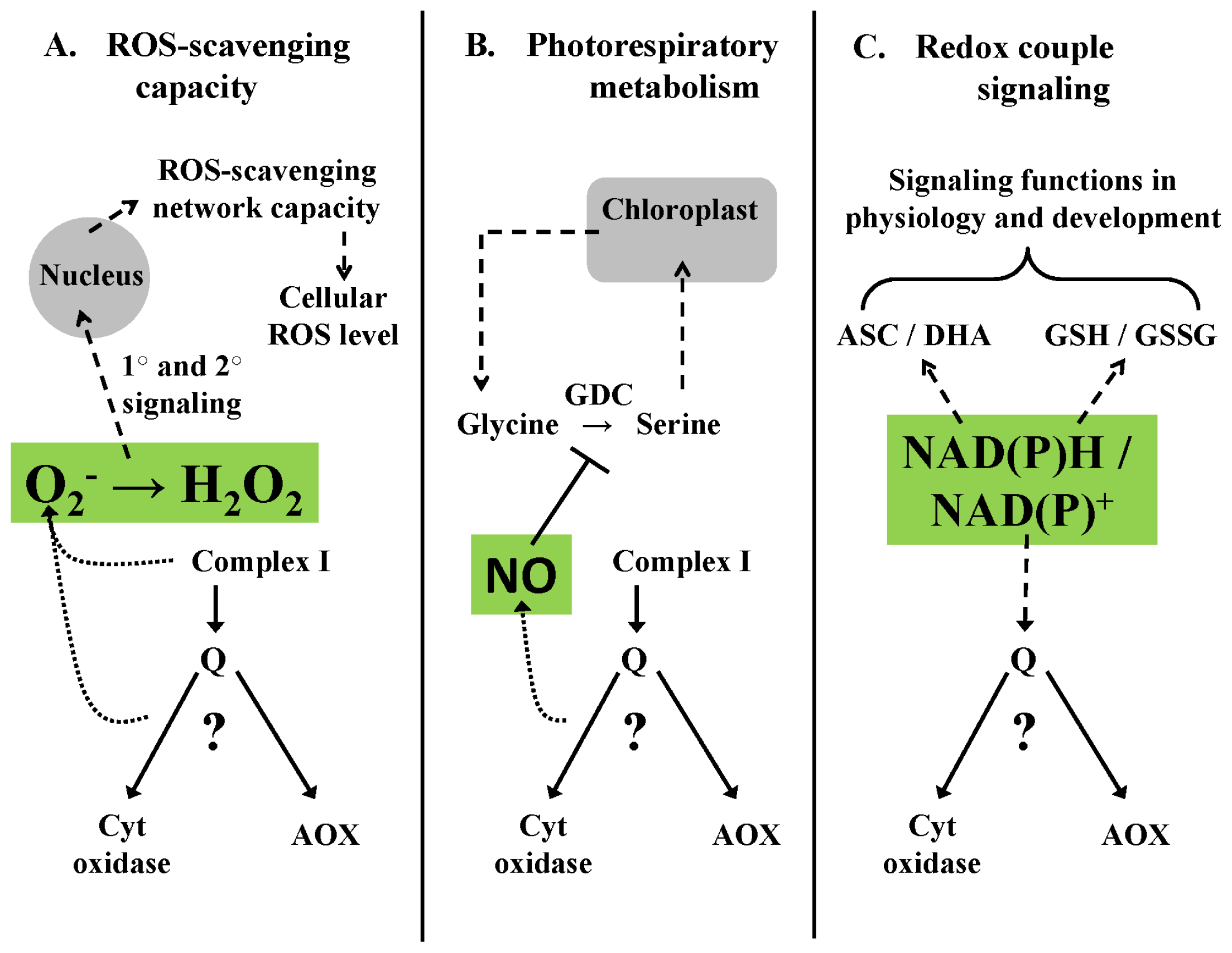
4.1. Homeostasis of ROS Signaling—An Example
4.2. Homeostasis of RNS Signaling—An Example
4.3. Homeostasis of Metabolite Signaling—An Example
5. Alternative Oxidase during Abiotic and Biotic Stress
5.1. Temperature Stress
5.2. Drought Stress
5.3. Nutrient Limitation
5.4. Salt Stress
5.5. Ozone
5.6. Metal Toxicity
5.7. Low Oxygen
5.8. Bacterial Pathogens
5.9. Viral Pathogens
5.10. Fungal Pathogens
6. Alternative Oxidase, Plant Growth and Stress Tolerance
6.1. A Role for AOX in the Optimization of Photosynthetic Metabolism
6.2. A Role for AOX during Specific Perturbation of the ETC
6.3. A Role for AOX during Oxidative Stress Conditions
6.4. A Role for AOX in Defining Susceptibility to Programmed Cell Death
6.5. A Role for AOX in Protection against Biotic Stress
6.6. The Relation of AOX with Overall Growth and Productivity
7. Conclusions
Acknowledgments
Abbreviations
| AA | antimycin A |
| ACC | 1-aminocyclopropane-1-carboxylic acid |
| AOX | alternative oxidase |
| CAS | β-cyanoalanine synthase |
| cyt | cytochrome |
| ETC | electron transport chain |
| GDC | glycine decarboxylase |
| G6PDH | glucose-6-phosphate dehydrogenase |
| HR | hypersensitive response |
| IMM | inner mitochondrial membrane |
| IMS | inner membrane space |
| MnSOD | manganese superoxide dismutase |
| NO | nitric oxide |
| O2− | superoxide |
| OAS-C | O-acetylserine(thiol)lyase C |
| OPP pathway | oxidative pentose phosphate pathway |
| RNS | reactive nitrogen species |
| ROS | reactive oxygen species |
| SA | salicylic acid |
| SAR | systemic acquired resistance |
| SHAM | salicylhydroxamic acid |
| TCA cycle | tricarboxylic acid cycle |
| TMV | tobacco mosaic virus |
| WT | wild-type |
Conflict of Interest
- Notes Added in ProofRecently, Shiba et al. [264] reported the first crystal structure of an AOX protein, in this case from the tropical parasite Trypanosoma brucei.
References
- Millar, A.H.; Whelan, J.; Soole, K.L.; Day, D.A. Organization and regulation of mitochondrial respiration in plants. Annu. Rev. Plant Biol 2011, 62, 79–104. [Google Scholar]
- Finnegan, P.M.; Soole, K.L.; Umbach, A.L. Alternative Mitochondrial Electron Transport Proteins in Plants. In Plant Mitochondria: From Genome to Function; Day, D.A., Millar, A.H., Whelan, J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 163–230. [Google Scholar]
- Gifford, R.M. Plant respiration in productivity models: Conceptualization, representation and issues of global terrestrial carbon-cycle research. Funct. Plant Biol 2003, 30, 171–186. [Google Scholar]
- Ciais, Ph.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, Chr.; Carrara, A.; et al. Europe-Wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar]
- Amthor, J.S. From sunlight to biomass: On the potential efficiency of converting solar radiation to phyto-energy. New Phytol 2010, 188, 939–959. [Google Scholar]
- Zhu, X.-G.; Long, S.P.; Ort, D.R. Improving photosynthetic efficiency for greater yield. Annu. Rev. Plant Biol 2010, 61, 235–261. [Google Scholar]
- Wilson, K.E.; Ivanov, A.G.; Öquist, G.; Grodzinski, B.; Sarhan, F.; Hüner, N.P.A. Energy balance, organellar redox status, and acclimation to environmental stress. Can. J. Bot 2006, 84, 1355–1370. [Google Scholar]
- Scharte, J.; Schön, H.; Weis, E.; von Schaewen, A. Isoenzyme replacement of glucose-6-phosphate dehydrogenase in the cytosol improves stress tolerance in plants. Proc. Natl. Acad. Sci. USA 2009, 106, 8061–8066. [Google Scholar]
- Dal Santo, S.; Stampfl, H.; Krasensky, J.; Kempa, S.; Gibon, Y.; Petutschnig, E.; Rozhon, W.; Heuck, A.; Clausen, T.; Jonak, C. Stress-Induced GSK3 regulates the redox stress response by phosphorylating glucose-6-phosphate dehydrogenase in Arabidopsis. Plant Cell 2012, 24, 3380–3392. [Google Scholar]
- Møller, I.M. Plant mitochondria and oxidative stress: Electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annu. Rev. Plant Physiol. Plant Mol. Biol 2001, 52, 561–591. [Google Scholar]
- Poyton, R.O.; Ball, K.A.; Castello, P.R. Mitochondrial generation of free radicals and hypoxic signaling. Trends Endocrinol. Metab 2009, 20, 332–340. [Google Scholar]
- Morgan, M.J.; Lehmann, M.; Schwarzländer, M.; Baxter, C.J.; Sienkiewicz-Porzucek, A.; Williams, T.C.R.; Schauer, N.; Fernie, M.D.; Ratcliffe, R.G.; Sweetlove, L.J.; et al. Decrease in manganese superoxide dismutase leads to reduced root growth and affects tricarboxylic acid cycle flux and mitochondrial redox homeostasis. Plant Physiol 2008, 147, 101–114. [Google Scholar]
- Planchet, E.; Gupta, K.J.; Sonoda, M.; Kaiser, W.M. Nitric oxide emission from tobacco leaves and cell suspensions: Rate limiting factors and evidence for the involvement of mitochondrial electron transport. Plant J 2005, 41, 732–743. [Google Scholar]
- Gupta, K.J.; Fernie, A.R.; Kaiser, W.M.; van Dongen, J.T. On the origins of nitric oxide. Trends Plant Sci 2010, 16, 160–168. [Google Scholar]
- Gleason, C.; Huang, S.; Thatcher, L.F.; Foley, R.C.; Anderson, C.R.; Carroll, A.J.; Millar, A.H.; Singh, K.B. Mitochondrial complex II has a key role in mitochondria-derived reactive oxygen species influence on plant stress gene regulation and defense. Proc. Natl. Acad. Sci. USA 2011, 108, 10768–10773. [Google Scholar]
- He, J.; Duan, Y.; Hua, D.; Fan, G.; Wang, L.; Liu, Y.; Chen, Z.; Han, L.; Qu, L.-J.; Gong, Z. DEXH Box RNA helicase-mediated mitochondrial reactive oxygen species production in Arabidopsis mediates crosstalk between abscisic acid and auxin signaling. Plant Cell 2012, 24, 1815–1833. [Google Scholar]
- Rhoads, D.M.; Plant, Mitochondrial. Retrograde Regulation. In Plant Mitochondria; Kempken, F., Ed.; Springer Science + Business Media, LLC 2011: New York, USA, 2011; Volume 1, pp. 411–437. [Google Scholar]
- Garmier, M.; Carroll, A.J.; Delannoy, E.; Vallet, C.; Day, D.A.; Small, I.D.; Millar, A.H. Complex I dysfunction redirects cellular and mitochondrial metabolism in Arabidopsis. Plant Physiol 2008, 148, 1324–1341. [Google Scholar]
- Yu, J.; Nickels, R.; McIntosh, L. A genome approach to mitochondrial-nuclear communication in Arabidopsis. Plant Physiol. Biochem 2001, 39, 345–353. [Google Scholar]
- Umbach, A.L.; Zarkovic, J.; Yu, J.; Ruckle, M.E.; McIntosh, L.; Hock, J.J.; Bingham, S.; White, S.J.; George, R.M.; Subbaiah, C.C.; et al. Comparision of intact Arabidopsis thaliana leaf transcript profiles during treatment with inhibitors of mitochondrial electron transport and TCA cycle. PLoS One 2012, 7, e44339. [Google Scholar]
- Busi, M.V.; Gomez-Lobato, M.E.; Rius, S.P.; Turowski, V.R.; Casati, P.; Zabaleta, E.J.; Gomez-Casati, D.F.; Araya, A. Effect of mitochondrial dysfunction on carbon metabolism and gene expression in flower tissues of Arabidopsis thaliana. Mol. Plant 2011, 4, 127–143. [Google Scholar]
- Geisler, D.A.; Päpke, C.; Obata, T.; Nunes-Nesi, A.; Matthes, A.; Schneitz, K.; Maximova, E.; Araújo, W.L.; Fernie, A.R.; Persson, S. Downregulation of the δ-subunit reduces mitochondrial ATP synthase levels, alters respiration, and restricts growth and gametophyte development in Arabidopsis. Plant Cell 2012, 24, 2792–2811. [Google Scholar]
- Van Aken, O.; Zhang, B.; Carrie, C.; Uggalla, V.; Paynter, E.; Giraud, E.; Whelan, J. Defining the mitochondrial stress response in Arabidopsis thaliana. Mol. Plant 2010, 2, 1310–1324. [Google Scholar]
- Schwarzländer, M.; König, A-C.; Sweetlove, L.J.; Finkemeier, I. The impact of impaired mitochondrial function on retrograde signalling: A meta-analysis of transcriptomic responses. J. Exp. Bot. 2012, 63, 1735–1750. [Google Scholar]
- Van Aken, O.; Whelan, J. Comparison of transcriptional changes to chloroplast and mitochondrial perturbations reveals common and specific responses inArabidopsis. Front. Plant Sci. 2012. [Google Scholar] [CrossRef]
- Kuzmin, E.V.; Karpova, O.V.; Elthon, T.E.; Newton, K.J. Mitochondrial respiratory deficiencies signal up-regulation of genes for heat shock proteins. J. Biol. Chem 2004, 279, 20672–20677. [Google Scholar]
- León, G.; Holuigue, L.; Jordana, X. Mitochondrial complex II is essential for gametophyte development in Arabidopsis. Plant Physiol 2007, 143, 1534–1546. [Google Scholar]
- Falcon de Longevialle, A.; Meyer, E.H.; Andrés, C.; Taylor, N.L.; Lurin, C.; Millar, A.H.; Small, I.D. The pentatricopeptide repeat gene OTP43 is required or trans-splicing of the mitochondrial nad1 intron 1 in Arabidopsis thaliana. Plant Cell 2007, 19, 3256–3265. [Google Scholar]
- Van Aken, O.; Pečenková, T.; van de Cotte, B.; de Rycke, R.; Eeckhout, D.; Fromm, H.; de Jaeger, G.; Witters, E.; Beemster, G.T.S.; Inzé, D.; et al. Mitochondrial type-I prohibitins of Arabidopsis thaliana are required for supporting proficient meristem development. Plant J 2007, 52, 850–864. [Google Scholar]
- Vidal, G.; Ribas-Carbo, M.; Garmier, M.; Dubertret, G.; Rasmusson, A.G.; Mathieu, C.; Foyer, C.H.; de Paepe, R. Lack of respiratory chain Complex I impairs alternative oxidase engagement and modulates redox signaling during elicitor-induced cell death in tobacco. Plant Cell 2007, 19, 640–655. [Google Scholar]
- Zsigmond, L.; Rigó, G.; Szarka, A.; Székely, G.; Ötvös, K.; Darula, Z.; Medzihradszky, K.F.; Koncz, C.; Koncz, Z.; Szabados, L. Arabidopsis PPR40 connects abiotic stress responses to mitochondrial electron transport. Plant Physiol 2008, 146, 1721–1737. [Google Scholar]
- Meyer, E.H.; Tomaz, T.; Carroll, A.J.; Estavillo, G.; Delannoy, E.; Tanz, S.K.; Small, I.D.; Pogson, B.J.; Millar, A.H. Remodeled respiration in ndufs4 with low phosphorylation efficiency suppresses Arabidopsis germination and growth and alters control of metabolism at night. Plant Physiol 2009, 151, 603–619. [Google Scholar]
- Liu, Y.; He, J.; Chen, Z.; Ren, X.; Hong, X.; Gong, Z. ABA overly-sensitive 5 (ABO5), encoding a pentatricopeptide repeat protein required for cis-splicing of mitochondrial nad2 intron 3, is involved in the abscisic acid response in Arabidopsis. Plant J 2010, 63, 749–765. [Google Scholar]
- Araújo, W.L.; Nunes-Nesi, A.; Nikoloski, Z.; Sweetlove, L.J. Metabolic control and regulation of the tricarboxylic acid cycle in photosynthetic and heterotrophic plant tissues. Plant Cell Environ 2012, 35, 1–21. [Google Scholar]
- Gupta, K.J.; Shah, J.K.; Brotman, Y.; Jahnke, K.; Willmitzer, L.; Kaiser, W.M.; Bauwe, H.; Igamberdiev, A.U. Inhibition of aconitase by nitric oxide leads to induction of the alternative oxidase and to a shift of metabolism towards biosynthesis of amino acids. J. Exp. Bot 2012, 63, 1773–1784. [Google Scholar]
- Solheim, C.; Li, L.; Hatzopoulus, P.; Millar, A.H. Loss of Lon1 in Arabidopsis changes the mitochondrial proteome leading to altered metabolic profiles and growth retardation without an accumulation of oxidative damage. Plant Physiol 2012, 160, 1187–1203. [Google Scholar]
- Sosso, D.; Mbelo, S.; Vernoud, V.; Gendrot, G.; Dedieu, A.; Chambrier, P.; Dauzat, M.; Heurtevin, L.; Guyon, V.; Takenaka, M.; et al. PPR2263, a DYW-subgroup pentatricopeptide repeat protein, is required for mitochondrial nad5 and cob transcript editing, mitochondrion biogenesis, and maize growth. Plant Cell 2012, 24, 676–691. [Google Scholar]
- Welchen, E.; Hildebrandt, T.M.; Lewejohann, D.; Gonzalez, D.H.; Braun, H.-P. Lack of cytochrome c in Arabidopsis decreases stability of Complex IV and modifies redox metabolism without affecting Complexes I and III. Biochim. Biophys. Acta 2012, 1817, 990–1001. [Google Scholar]
- McDonald, A.E.; Vanlerberghe, G.C. Origins, evolutionary history, and taxonomic distribution of alternative oxidase and plastoquinol terminal oxidase. Comp. Biochem. Physiol. D 2006, 1, 357–364. [Google Scholar]
- Watling, J.R.; Robinson, S.A.; Seymour, R.S. Contribution of the alternative pathway to respiration during thermogenesis in flowers of the sacred lotus. Plant Physiol 2006, 140, 1367–1373. [Google Scholar]
- Wagner, A.M.; Krab, K.; Wagner, M.J.; Moore, A.L. Regulation of thermogenesis in flowering Araceae: The role of the alternative oxidase. Biochim. Biophys. Acta 2008, 1777, 993–1000. [Google Scholar]
- Miller, R.E.; Grant, N.M.; Giles, L.; Ribas-Carbo, M.; Berry, J.A.; Watling, J.R.; Robinson, S.A. In the heat of the night—Alternative pathway respiration drives thermogenesis in Philodendron bipinnatifidum. New Phytol 2011, 189, 1013–1026. [Google Scholar]
- Stenmark, P.; Nordlund, P. A prokaryotic alternative oxidase present in the bacterium Novosphingobium. aromaticivorans. FEBS Lett 2003, 552, 189–192. [Google Scholar]
- Missall, T.A.; Lodge, J.K.; McEwen, J.E. Mechanisms of resistance to oxidative and nitrosative stress: implications for fungal survival in mammalian hosts. Eukaryot. Cell 2004, 3, 835–846. [Google Scholar]
- Chaudhuri, M.; Ott, R.D.; Hill, G.C. Trypanosome alternative oxidase: From molecule to function. Trends Parasitol 2006, 22, 484–491. [Google Scholar]
- Nargang, F.E.; Adames, K.; Rüb, C.; Cheung, S.; Easton, N.; Nargang, C.E. Identification of genes required for alternative oxidase production in the Neurospora crassa gene knockout library. Genes Genomes Genet 2012, 2, 1345–1356. [Google Scholar]
- Scheckhuber, C.Q.; Houthoofd, K.; Weil, A.C.; Werner, A.; de Vresse, A.; Vanfleteren, J.R.; Osiewacz, H.D. Alternative oxidase dependent respiration leads to an increased mitochondrial content in two long-lived mutants of the ageing model Podospora anserina. PLoS One 2011, 6, e16620. [Google Scholar]
- McDonald, A.E.; Vanlerberghe, G.C. Branched mitochondrial electron transport in the Animalia: Presence of alternative oxidase in several animal phyla. IUBMB Life 2004, 56, 333–341. [Google Scholar]
- Cannino, G.; El-Khoury, R.; Pirinen, M.; Hutz, B.; Rustin, P.; Jacobs, H.T.; Dufour, E. Glucose modulates respiratory Complex I activity in response to acute mitochondrial dysfunction. J. Biol. Chem 2012, 287, 38729–38740. [Google Scholar]
- Humphrey, D.M.; Parsons, R.B.; Ludlow, Z.N.; Riemensperger, T.; Esposito, G.; Verstreken, P.; Jacobs, H.T.; Birman, S.; Hirth, F. Alternative oxidase rescues mitochondria-mediated dopaminergic cell loss in Drosophila. Hum. Mol. Genet 2012, 21, 2698–2712. [Google Scholar]
- El-Khoury, R.; Dufour, E.; Rak, M.; Ramanantsoa, N.; Grandchamp, N.; Csaba, Z.; Duvillié, B.; Bénit, P.; Gallego, J.; Gressens, P.; et al. Alternative oxidase expression in the mouse enables bypassing cytochrome c oxidase blockade and limits mitochondrial ROS overproduction. PLoS Genet 2013, 9, e1003182. [Google Scholar]
- Moller, I.M.; Berczi, A.; van der Plas, L.H.W.; Lambers, H. Measurement of the activity and capacity of the alternative pathway in intact plant tissues: Identification of problems and possible solutions. Physiol. Plant 1988, 72, 642–649. [Google Scholar]
- McDonald, A.E.; Sieger, S.M.; Vanlerberghe, G.C. Methods and approaches to study plant mitochondrial alternative oxidase. Physiol. Plant 2002, 116, 135–143. [Google Scholar]
- Guy, R.D.; Berry, J.A.; Fogel, M.L.; Hoering, T.C. Differential fractionation of oxygen isotopes by cyanide-resistant and cyanide-sensitive respiration in plants. Planta 1989, 177, 483–491. [Google Scholar]
- Nagel, O.W.; Waldron, S.; Jones, H.G. An off-line implementation of the stable isotope technique for measurements of alternative respiratory pathway activities. Plant Physiol 2001, 127, 1279–1286. [Google Scholar]
- Kornfeld, A.; Horton, T.W.; Yakir, D.; Searle, S.Y.; Griffin, K.L.; Atkin, O.K.; Subke, J.-A.; Turnbull, M.H. A field-compatible method for measuring alternative pathway activities in vivo using stable O2 isotopes. Plant Cell Environ 2012, 35, 1518–1532. [Google Scholar]
- Kornfeld, A.; Atkin, O.K.; Griffin, K.L.; Horton, T.W.; Yakir, D.; Turnbull, M.H. Modulation of respiratory metabolism in response to nutrient changes along a soil chronosequence. Plant Cell Environ. 2013. [Google Scholar] [CrossRef]
- Millar, A.H.; Atkin, O.K.; Menz, I.; Henry, B.; Farquhar, G.; Day, D.A. Analysis of respiratory chain regulation in roots of soybean seedlings. Plant Physiol 1998, 117, 1083–1093. [Google Scholar]
- Florez-Sarasa, I.; Flexas, J.; Rasmusson, A.G.; Umback, A.L.; Siedow, J.N.; Ribas-Carbo, M. In vivo cytochrome and alternative pathway respiration in leaves of Arabidopsis thaliana plants with altered alternative oxidase under different light conditions. Plant Cell Environ 2011, 34, 1373–1383. [Google Scholar]
- Yoshida, K.; Watanabe, C.K.; Terashima, I.; Noguchi, K. Physiological impact of mitochondrial alternative oxidase on photosynthesis and growth in Arabidopsis thaliana. Plant Cell Environ 2011, 34, 1890–1899. [Google Scholar]
- Gandin, A.; Duffes, C.; Day, D.A.; Cousins, A.B. The absence of alternative oxidase AOX1A results in altered response of photosynthetic carbon assimilation to increasing CO2 in Arabidopsis thaliana. Plant Cell Physiol 2012, 53, 1627–1637. [Google Scholar]
- Robinson, S.A.; Ribas-Carbo, M.; Yakir, D.; Giles, L.; Reuveni, Y.; Berry, J.A. Beyond SHAM and cyanide: Opportunities for studying the alternative oxidase in plant respiration using oxygen isotope discrimination. Aust. J. Plant Physiol 1995, 22, 487–496. [Google Scholar]
- Henry, B.K.; Atkin, O.K.; Day, D.A.; Millar, A.H.; Menz, R.I.; Farquhar, G.D. Calculation of the oxygen isotope discrimination factor for studying plant respiration. Aust. J. Plant Physiol 1999, 26, 773–780. [Google Scholar]
- Considine, M.J.; Holtzapffel, R.C.; Day, D.A.; Whelan, J.; Millar, A.H. Molecular distinction between alternative oxidase from monocots and dicots. Plant Physiol 2002, 129, 949–953. [Google Scholar]
- Clifton, R.; Millar, A.H.; Whelan, J. Alternative oxidases in Arabidopsis: A comparative analysis of differential expression in the gene family provides new insights into function of non-phosphorylating bypasses. Biochim. Biophys. Acta 2006, 1757, 730–741. [Google Scholar]
- Saisho, D.; Nakazono, M.; Lee, K.H.; Tsutsumi, N.; Akita, S.; Hirai, A. The gene for alternative oxidase-2 (AOX2) from Arabidopsis thaliana consists of five exons unlike other AOX genes and is transcribed at an early stage during germination. Genes Genet. Syst 2001, 76, 89–97. [Google Scholar]
- Chai, T.-T.; Simmonds, D.; Day, D.A.; Colmer, T.D.; Finnegan, P.M. Photosynthetic performance and fertility are repressed in GmAOX2b antisense soybean. Plant Physiol 2010, 152, 1638–1649. [Google Scholar]
- Macherel, D.; Benamar, A.; Avelange-Macherel, M.-H.; Tolleter, D. Function and stress tolerance of seed mitochondria. Physiol. Plant 2007, 129, 233–241. [Google Scholar]
- Vanlerberghe, G.C.; McIntosh, L. Mitochondrial electron transport regulation of nuclear gene expression: Studies with the alternative oxidase gene of tobacco. Plant Physiol 1994, 105, 867–874. [Google Scholar]
- Karpova, O.V.; Kuzmin, E.V.; Elthon, T.E.; Newton, K.J. Differential expression of alternative oxidase genes in maize mitochondrial mutants. Plant Cell 2002, 14, 3271–3284. [Google Scholar]
- Vanlerberghe, G.C.; McIntosh, L. Signals regulating the expression of the nuclear gene encoding alternative oxidase of plant mitochondria. Plant Physiol 1996, 111, 589–595. [Google Scholar]
- Juszczuk, I.M.; Szal, B.; Rychter, A.M. Oxidation-Reduction and reactive oxygen species homeostasis in mutant plants with respiratory chain complex I dysfunction. Plant Cell Environ 2012, 35, 296–307. [Google Scholar]
- Zarkovic, J.; Anderson, S.L.; Rhoads, D.M. A reporter gene system used to study developmental expression of alternative oxidase and isolate mitochondrial retrograde regulation mutants in Arabidopsis. Plant Mol. Biol 2005, 57, 871–888. [Google Scholar]
- Ng, S.; Giraud, E.; Duncan, O.; Law, S.R.; Wang, Y.; Xu, L.; Narsai, R.; Carrie, C.; Walker, H.; Day, D.A.; et al. Cyclin-Dependent kinase E;1 (CDKE;1) provides a cellular switch in plants between growth and stress responses. J. Biol. Chem 2013, 288, 3449–3459. [Google Scholar]
- Giraud, E.; van Aken, O.; Ho, L.H.M.; Whelan, J. The transcription factor ABI4 is a regulator of mitochondrial retrograde expression of ALTERNATIVE OXIDASE1a. Plant Physiol 2009, 150, 1286–1296. [Google Scholar]
- León, P.; Gregorio, J.; Cordoba, E. ABI4 and its role in chloroplast retrograde communication. Frontiers in Plant. Science 2013, 3. [Google Scholar] [CrossRef]
- Vanderauwera, S.; Vandenbroucke, K.; Inzé, A.; van de Cotte, B.; Mühlenbock, P.; de Rycke, R.; Naouar, N.; van Gaever, T.; van Montagu, M.C.E.; van Breusegem, F. AtWRKY15 perturbation abolishes the mitochondrial stress response that steers osmotic stress tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 20113–20118. [Google Scholar]
- Dojcinovic, D.; Krosting, J.; Harris, A.J.; Wagner, D.J.; Rhoads, D.M. Identification of a region of the Arabidopsis AtAOX1a promoter necessary for mitochondrial retrograde regulation of expression. Plant Mol. Biol 2005, 58, 159–175. [Google Scholar]
- Millenaar, F.F.; Gonzalez-Meler, M.A.; Siedow, J.N.; Wagner, A.M.; Lambers, H. Role of sugars and organic acids in regulating the concentration and activity of the alternative oxidase in Poa annua roots. J. Exp. Bot. 2002, 53, 1081–1088. [Google Scholar]
- Ho, L.H.M.; Giraud, E.; Uggalla, V.; Lister, R.; Clifton, R.; Glen, A.; Thirkettle-Watts, D.; Van Aken, O.; Whelan, J. Identification of regulatory pathways controlling gene expression of stress-responsive mitochondrial proteins in Arabidopsis. Plant Physiol 2008, 147, 1858–1873. [Google Scholar]
- Maxwell, D.P.; Nickels, R.; McIntosh, L. Evidence of mitochondrial involvement in the transduction of signals required for the induction of genes associated with pathogen attack and senescence. Plant J 2002, 29, 269–279. [Google Scholar]
- Li, C.-R.; Liang, D.-D.; Li, J.; Duan, Y.-B.; Li, H.; Yang, Y.-C.; Qin, R.Y.; Li, L.; Wei, P.-C.; Yang, J.-B. Unravelling mitochondrial retrograde regulation in the abiotic stress induction of rice ALTERNATIVE OXIDASE 1 genes. Plant Cell Environ 2013, 36, 775–788. [Google Scholar]
- Takahashi, Y.; Berberich, T.; Miyazaki, A.; Seo, S.; Ohashi, Y.; Kusano, T. Spermine signaling in tobacco: Activation of mitogen-activated protein kinases by spermine is mediated through mitochondrial dysfunction. Plant J 2003, 36, 820–829. [Google Scholar]
- Arpagaus, S.; Rawyler, A.; Braendle, R. Occurrence and characteristics of the mitochondrial permeability transition in plants. J. Biol. Chem 2002, 277, 1780–1787. [Google Scholar]
- Tiwari, B.S.; Belenghi, B.; Levine, A. Oxidative stress increased respiration and generation of reactive oxygen species, resulting in ATP depletion, opening of mitochondrial permeability transition, and programmed cell death. Plant Physiol 2002, 128, 1271–1281. [Google Scholar]
- Rhoads, D.M.; McIntosh, L. Salicylic acid regulation of respiration in higher plants: Alternative oxidase expression. Plant Cell 1992, 4, 1131–1139. [Google Scholar]
- Cvetkovska, M.; Vanlerberghe, G.C. Coordination of a mitochondrial superoxide burst during the hypersensitive response to bacterial pathogen in Nicotiana tabacum. Plant Cell Environ 2012, 35, 1121–1136. [Google Scholar]
- Clifton, R.; Lister, R.; Parker, K.L.; Sappl, P.G.; Elhafez, D.; Millar, A.H.; Day, D.A.; Whelan, J. Stress-Induced co-expression of alternative respiratory chain components in Arabidopsis thaliana. Plant Mol. Biol 2005, 58, 193–212. [Google Scholar]
- Ho, L.H.M.; Giraud, E.; Lister, R.; Thirkettle-Watts, D.; Low, J.; Clifton, R.; Howell, K.A.; Carrie, C.; Donald, T.; Whelan, J. Characterization of the regulatory and expression context of an alternative oxidase gene provides insights into cyanide-insensitive respiration during growth and development. Plant Physiol 2007, 143, 1519–1533. [Google Scholar]
- Smith, C.A.; Melino, V.J.; Sweetman, C.; Soole, K.L. Manipulation of alternative oxidase can influence salt tolerance in Arabidopsis thaliana. Physiol. Plant 2009, 137, 459–472. [Google Scholar]
- Smith, C.; Barthet, M.; Melino, V.; Smith, P.; Day, D.; Soole, K. Alterations in the mitochondrial alternative NAD(P)H dehydrogenase NDB4 lead to changes in mitochondrial electron transport chain composition, plant growth and response to oxidative stress. Plant Cell Physiol 2011, 52, 1222–1237. [Google Scholar]
- Berthold, D.A.; Stenmark, P. Membrane-bound diiron carboxylate proteins. Annu. Rev. Plant Biol 2003, 54, 497–517. [Google Scholar]
- Albury, M.S.; Affourtit, C.; Crichton, P.G.; Moore, A.L. Structure of the plant alternative oxidase: Site-Directed mutagenesis provides new information on the active site and membrane topology. J. Biol. Chem 2002, 277, 1190–1194. [Google Scholar]
- Moore, A.L.; Albury, M.S. Further insights into the structure of the alternative oxidase: From plants to parasites. Biochem. Soc. Trans 2008, 36, 1022–1026. [Google Scholar]
- Berthold, D.A.; Voevodskaya, N.; Stenmark, P.; Gräsland, A.; Nordlund, P. EPR studies of the mitochondrial alternative oxidase: Evidence for a diiron carboxylate center. J. Biol. Chem 2002, 277, 43608–43614. [Google Scholar]
- Moore, A.L.; Carré, J.E.; Affourtit, C.; Albury, M.S.; Crichton, P.G.; Kita, K.; Heathcote, P. Compelling EPR evidence that the alternative oxidase is a diiron carboxylate protein. Biochim. Biophys. Acta 2008, 1777, 327–33. [Google Scholar]
- Albury, M.S.; Elliott, C.; Moore, A.L. Ubiquinol-binding site in the alternative oxidase: Mutagenesis reveals features important for substrate binding and inhibition. Biochim. Biophys. Acta 2010, 1797, 1933–1939. [Google Scholar]
- Kido, Y.; Shiba, T.; Inaoka, D.K.; Sakamoto, K.; Nara, T.; Aoki, T.; Honma, T.; Tanaka, A.; Inoue, M.; Matsuoka, S.; et al. Crystallization and preliminary analysis of cyanide-insensitive alternative oxidase from Trypanosoma brucei brucei. Acta Crystallogr 2010, F66, 275–278. [Google Scholar]
- Umbach, A.L.; Siedow, J.N. Covalent and noncovalent dimers of the cyanide-resistant alternative oxidase protein in higher plant mitochondria and their relationship to enzyme activity. Plant Physiol 1993, 103, 845–854. [Google Scholar]
- Rhoads, D.M.; Umbach, A.L.; Sweet, C.R.; Lennon, A.M.; Rauch, G.S.; Siedow, J.N. Regulation of the cyanide-resistant alternative oxidase of plant mitochondria: Identification of the cysteine residue involved in α-keto acid stimulation and intersubunit disulfide bond formation. J. Biol. Chem 1998, 273, 30750–30756. [Google Scholar]
- Vanlerberghe, G.C.; McIntosh, L.; Yip, J.Y.H. Molecular localization of a redox-modulated process regulating plant mitochondrial electron transport. Plant Cell 1998, 10, 1551–1560. [Google Scholar]
- Vanlerberghe, G.C.; Day, D.A.; Wiskich, J.T.; Vanlerberghe, A.E.; McIntosh, L. Alternative oxidase activity in tobacco leaf mitochondria: Dependence on tricarboxylic acid cycle-mediated redox regulation and pyruvate activation. Plant Physiol 1995, 109, 353–361. [Google Scholar]
- Gelhaye, E.; Rouhier, N.; Gérard, J.; Jolivet, Y.; Gualberto, J.; Navrot, N.; Ohlsson, P.-I.; Wingsle, G.; Hirasawa, M.; Knaff, D.B.; et al. A specific form of thioredoxin h occurs in plant mitochondria and regulates the alternative oxidase. Proc. Natl. Acad. Sci. USA 2004, 101, 14545–14550. [Google Scholar]
- Millar, A.H.; Wiskich, J.T.; Whelan, J.; Day, D.A. Organic acid activation of the alternative oxidase of plant mitochondria. FEBS Lett 1993, 329, 259–262. [Google Scholar]
- Carré, J.E.; Affourtit, C.; Moore, A.L. Interaction of purified alternative oxidase from thermogenic Arum maculatum with pyruvate. FEBS Lett 2011, 585, 397–401. [Google Scholar]
- Umbach, A.L.; Gonzàlez-Meler, M.A.; Sweet, C.R.; Siedow, J.N. Activation of the plant mitochondrial alternative oxidase: Insights from site-directed mutagenesis. Biochim. Biophys. Acta 2002, 1554, 118–128. [Google Scholar]
- Holtzapffel, R.C.; Castelli, J.; Finnegan, P.M.; Millar, A.H.; Whelan, J.; Day, D.A. A tomato alternative oxidase protein with altered regulatory properties. Biochim. Biophys. Acta 2003, 1606, 153–162. [Google Scholar]
- Grant, N.; Onda, Y.; Kakizaki, Y.; Ito, K.; Watling, J.; Robinson, S. Two Cys or not two Cys? That is the question; alternative oxidase in the thermogenic plant sacred lotus. Plant Physiol 2009, 150, 987–995. [Google Scholar]
- Crichton, P.G.; Affourtit, C.; Albury, M.S.; Carré, J.E.; Moore, A.L. Constitutive activity of Sauromatum guttatum alternative oxidase in Schizosaccharomyces pombe implicates residues in addition to conserved cysteines in α-keto acid activation. FEBS Lett 2005, 579, 331–336. [Google Scholar]
- Umbach, A.L.; Ng, V.S.; Siedow, J.N. Regulation of plant alternative oxidase activity: A tale of two cysteines. Biochim. Biophys. Acta 2006, 1757, 135–142. [Google Scholar]
- Guy, R.D.; Vanlerberghe, G.C. Partitioning of respiratory electrons in the dark in leaves of transgenic tobacco with modified levels of alternative oxidase. Physiol. Plant 2005, 125, 171–180. [Google Scholar]
- Purvis, A.C. Role of the alternative oxidase in limiting superoxide production by plant mitochondria. Physiol. Plant 1997, 100, 165–170. [Google Scholar]
- Cvetkovska, M.; Vanlerberghe, G.C. Alternative oxidase modulates leaf mitochondrial concentrations of superoxide and nitric oxide. New Phytol 2012, 195, 32–39. [Google Scholar]
- Cvetkovska, M.; Vanlerberghe, G.C. Alternative oxidase impacts the plant response to biotic stress by influencing the mitochondrial generation of reactive oxygen species. Plant Cell Environ 2013, 36, 721–732. [Google Scholar]
- Leakey, A.D.B.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. J. Exp. Bot 2009, 60, 2859–2876. [Google Scholar]
- Gomez-Casanovas, N.; Blanc-Betes, E.; Gonzalez-Meler, M.A.; Azcon-Bieto, J. Changes in respiratory mitochondrial machinery and cytochrome and alternative pathway activities in response to energy demand underlie the acclimation of respiration to elevated CO2 in thee invasive Opuntia ficus-indica. Plant Physiol 2007, 145, 49–61. [Google Scholar]
- Gandin, A.; Lapointe, L.; Dizengremel, P. The alternative respiratory pathway allows sink to cope with changes in carbon availability in the sink-limited plant Erythronium americanum. J. Exp. Bot 2009, 60, 4235–4248. [Google Scholar]
- Escobar, M.A.; Geisler, D.A.; Rasmusson, A.G. Reorganization of the alternative pathways of the Arabidopsis respiratory chain by nitrogen supply: Opposing effects of ammonium and nitrate. Plant J 2006, 45, 775–788. [Google Scholar]
- Hachiya, T.; Watanabe, C.K.; Boom, C.; Tholen, D.; Takahara, K.; Kawai-Yamada, M.; Uchimiya, H.; Uesono, Y.; Terashima, I.; Noguchi, K. Ammonium-dependent respiratory increase is dependent on the cytochrome pathway in Arabidopsis thaliana shoots. Plant Cell Environ 2010, 33, 1888–1897. [Google Scholar]
- Lambers, H.; Szaniawski, R.K.; de Visser, R. Respiration for growth, maintenance and ion uptake: An evaluation of concept, methods, values and their significance. Physiol. Plant 1983, 58, 556–563. [Google Scholar]
- Poorter, H.; van der Werf, A.; Atkin, O.K.; Lambers, H. Respiratory energy requirements of roots vary with the potential growth rate of a plant species. Physiol. Plant 1991, 83, 469–475. [Google Scholar]
- Britto, D.T.; Siddiqi, M.Y.; Glass, A.D.M.; Kronzucker, H.J. Futile transmembrane NH4 + cycling: A cellular hypothesis to explain ammonium toxicity in plants. Proc. Natl. Acad. Sci. USA 2001, 98, 4255–4258. [Google Scholar]
- Kurimoto, K.; Millar, A.H.; Lambers, H.; Day, D.A.; Noguchi, K. Maintenance of growth rate at low temperature in rice nad wheat cultivars with a high degree of respiratory homeostasis is associated with a high efficiency of respiratory ATP production. Plant Cell Physiol 2004, 45, 1015–1022. [Google Scholar]
- Malagoli, P.; Britto, D.T.; Schulze, L.M.; Kronzucker, H.J. Futile Na+ cycling at the root plasma membrane in rice (Oryza. sativa, L.): Kinetics, energetics, and relationship to salinity tolerance. J. Exp. Bot 2008, 59, 4109–4117. [Google Scholar]
- Noctor, G.; De Paepe, R.; Foyer, C.H. Mitochondrial redox biology and homeostasis in plants. Trends Plant Sci 2007, 12, 125–134. [Google Scholar]
- Diamond, M.; McCabe, P.F. Mitochondrial Regulation of Plant Programmed Cell Death. In Plant Mitochondria; Kempken, F., Ed.; Springer Science + Business Media, LLC 2011: New York, NY, USA, 2011; Volume 1, pp. 439–465. [Google Scholar]
- Echtay, K.S. Mitochondrial uncoupling proteins—What is their physiological function? Free Radic. Biol. Med 2007, 43, 1351–1371. [Google Scholar]
- Amirsadeghi, S.; Robson, C.A.; McDonald, A.E.; Vanlerberghe, G.C. Changes in plant mitochondrial electron transport alter cellular levels of reactive oxygen species and susceptibility to cell death signaling molecules. Plant Cell Physiol. 2006, 47, 1509–1519. [Google Scholar]
- Giraud, E.; Ho, L.H.M.; Clifton, R.; Carroll, A.; Estavillo, G.; Tan, Y.-F.; Howell, K.A.; Ivanov, A.; Pogson, B.J.; Millar, A.H.; et al. The absence of ALTERNATIVE OXIDASE1a in Arabidopsis results in acute sensitivity to combined light and drought stress. Plant Physiol 2008, 147, 595–610. [Google Scholar]
- Sweetlove, L.J.; Lytovchenko, A.; Morgan, M.; Nunes-Nesi, A.; Taylor, N.L.; Baxter, C.J.; Eickmeier, I.; Fernie, A.R. Mitochondrial uncoupling protein is required for efficient photosynthesis. Proc. Natl. Acad. Sci. USA 2006, 103, 19587–19592. [Google Scholar]
- Timm, S.; Florian, A.; Arrivault, S.; Stitt, M.; Fernie, A.R. Glycine decarboxylase controls photosynthesis and plant growth. FEBS Lett 2012, 586, 3692–3697. [Google Scholar]
- Palmieri, M.C.; Lindermayr, C.; Bauwe, H.; Steinhauser, C.; Durner, J. Regulation of plant glycine decarboxylase by S-nitrosylation and glutathionylation. Plant Physiol 2010, 152, 1514–1528. [Google Scholar]
- Taylor, N.L.; Day, D.A.; Millar, A.H. Environmental stress causes oxidative damage to plant mitochondria leading to inhibition of glycine decarboxylase. J. Biol. Chem 2002, 277, 42663–42668. [Google Scholar]
- Noctor, G. Metabolic signaling in defense and stress: The central roles of soluble redox couples. Plant Cell Environ 2006, 29, 409–425. [Google Scholar]
- Liu, Y.-J.; Nunes-Nesi, A.; Wallström, S.V.; Lager, I.; Michalecka, A.M.; Norberg, F.E.B.; Widell, S.; Fredlund, K.M.; Fernie, A.R.; Rasmusson, A.G. A redox-mediated modulation of stem bolting in transgenic Nicotiana sylvestris differentially expressing the external mitochondrial NADPH dehydrogenase. Plant Physiol 2009, 150, 1248–1259. [Google Scholar]
- Verdin, E.; Hirschey, M.D.; Finley, L.W.S.; Haigis, M.C. Sirtuin regulation of mitochondria: Energy production, apoptosis, and signaling. Trends Biochem. Sci 2010, 35, 669–675. [Google Scholar]
- Finkemeier, I.; Laxa, M.; Miguet, L.; Howden, A.J.M.; Sweetlove, L.J. Proteins of diverse function and subcellular location are lysine acetylated in Arabidopsis. Plant Physiol 2011, 155, 1779–1790. [Google Scholar]
- Wu, X.; Oh, M.-H.; Schwarz, E.M.; Larue, C.T.; Sivaguru, M.; Imai, B.S.; Yau, P.M.; Ort, D.R.; Huber, S.C. Lysine acetylation is a widespread protein modification for diverse proteins in Arabidopsis. Plant Physiol 2011, 155, 1769–1778. [Google Scholar]
- Millar, A.H.; Mittova, V.; Kiddle, G.; Heazlewood, J.L.; Bartoli, C.T.G.; Theodoulou, F.L.; Foyer, C.H. Control of ascorbate synthesis by respiration and its implications for stress responses. Plant Physiol 2003, 133, 443–447. [Google Scholar]
- Bartoli, C.G.; Gómez, F.; Fernández, L.; McIntosh, L.; Foyer, C.H. Inter-Relationships between light and respiration in the control of ascorbic acid synthesis and accumulation in Arabidopsis thaliana leaves. J. Exp. Bot 2006, 57, 1621–1631. [Google Scholar]
- Wang, J.; Rajakulendran, N.; Amirsadeghi, S.; Vanlerberghe, G.C. Impact of mitochondrial alternative oxidase expression on the response of Nicotiana tabacum to cold temperature. Physiol. Plant 2011, 142, 339–351. [Google Scholar]
- Vanlerberghe, G.C.; McIntosh, L. Lower growth temperature increases alternative pathway capacity and alternative oxidase protein in tobacco. Plant Physiol 1992, 100, 115–119. [Google Scholar]
- Ito, Y.; Saisho, D.; Nakazono, M.; Tsutsumi, N.; Hirai, A. Transcript levels of tandem-arranged alternative oxidase genes in rice are increased by low temperature. Gene 1997, 203, 121–129. [Google Scholar]
- Gonzàlez-Meler, M.A.; Ribas-Carbo, M.; Giles, L.; Siedow, J.N. The effect of growth and measurement temperature on the activity of the alternative respiratory pathway. Plant Physiol 1999, 120, 765–772. [Google Scholar]
- Fiorani, F.; Umbach, A.L.; Siedow, J.N. The alternative oxidase of plant mitochondria is involved in the acclimation of shoot growth at low temperature. A study of Arabidopsis AOX1a transgenic plants. Plant Physiol 2005, 139, 1795–1805. [Google Scholar]
- Sugie, A.; Naydenov, N.; Mizuno, N.; Nakamura, C.; Takumi, S. Overexpression of wheat alternative oxidase gene Waox1a alters respiration capacity and response to reactive oxygen species under low temperature in transgenic Arabidopsis. Genes Genet. Syst 2006, 81, 349–354. [Google Scholar]
- Campbell, C.; Atkinson, L.; Zaragoza-Castells, J.; Lundmark, M.; Atkin, O.; Hurry, V. Acclimation of photosynthesis and respiration is asynchronous in response to changes in temperature regardless of plant functional group. New Phytol 2007, 176, 375–389. [Google Scholar]
- Armstrong, A.F.; Badger, M.R.; Day, D.A.; Barthet, M.M.; Smith, P.M.C.; Millar, A.H.; Whelan, J.; Atkin, O.K. Dynamic changes in the mitochondrial electron transport chain underpinning cold acclimation of leaf respiration. Plant Cell Environ 2008, 31, 1156–1169. [Google Scholar]
- Mizuno, N.; Sugie, A.; Kobayashi, F.; Takumi, S. Mitochondrial alternative pathway is associated with development of freezing tolerance in common wheat. J. Plant Physiol 2008, 165, 462–467. [Google Scholar]
- Usadel, B.; Bläsing, O.E.; Gibon, Y.; Poree, F.; Höhne, M.; Günter, M.; Trethewey, R.; Kamlage, B.; Poorter, H.; Stitt, M. Multilevel genomic analysis of the response of transcripts, enzyme activities and metabolites in Arabidopsis rosettes to a progressive decrease of temperature in the non-freezing range. Plant Cell Environ 2008, 31, 518–547. [Google Scholar]
- Watanabe, C.K.; Hachiya, T.; Terashima, I.; Noguchi, K. The lack of alternative oxidase at low temperature leads to a disruption of the balance in carbon and nitrogen metabolism, and to an up-regulation of antioxidant defense systems in Arabidopsis thaliana leaves. Plant Cell Environ 2008, 31, 1190–1202. [Google Scholar]
- Umbach, A.L.; Lacey, E.P.; Richter, S.J. Temperature-Sensitive alternative oxidase protein content and its relationship to floral reflectance in natural Plantago lanceolata populations. New Phytol 2009, 181, 662–671. [Google Scholar]
- Wang, H.; Huang, J.; Liang, X.; Bi, Y. Involvement of hydrogen peroxide, calcium, and ethylene in the induction of the alternative pathway in chilling-stressed Arabidopsis callus. Planta 2012, 235, 53–67. [Google Scholar]
- Shi, K.; Fu, L.-J.; Zhang, S.; Li, X.; Liao, Y.-W.-K.; Xia, X.-J.; Zhou, Y.-H.; Wang, R.-Q.; Chen, Z.-X.; Yu, J.-Q. Flexible change and cooperation between mitochondrial electron transport and cytosolic glycolysis as the basis for chilling tolerance in tomato plants. Planta 2013, 237, 589–601. [Google Scholar]
- MacFarlane, C.; Hansen, L.D.; Florez-Sarasa, I.; Ribas-Carbo, M. Plant mitochondria electron partitioning is independent of short-term temperature changes. Plant Cell Environ 2009, 32, 585–591. [Google Scholar]
- Ribas-Carbo, M.; Aroca, R.; Gonzàlez-Meler, M.A.; Irigoyen, J.J.; Sánchez-Díaz, M. The electron partitioning between the cytochrome and alternative respiratory pathways during chilling recovery in two cultivars of maize differing in chilling sensitivity. Plant Physiol 2000, 122, 199–204. [Google Scholar]
- Searle, S.Y.; Thomas, S.; Griffin, K.L.; Horton, T.; Kornfeld, A.; Yakir, D.; Hurry, V.; Turnbull, M.H. Leaf respiration and alternative oxidase in field-grown alpine grasses respond to natural changes in temperature and light. New Phytol 2011, 189, 1027–1039. [Google Scholar]
- Rachmilevitch, S.; Xu, Y.; Gonzalez-Meler, M.A.; Huang, B.; Lambers, H. Cytochrome and alternative pathway activity in roots of thermal and non-thermal Agrostis species in response to high soil temperature. Physiol. Plant 2007, 129, 163–174. [Google Scholar]
- Searle, S.Y.; Turnbull, M.H. Seasonal variation of leaf respiration and the alternative pathway in field-grown Populus × canadensis. Physiol. Plant 2011, 141, 332–342. [Google Scholar]
- Searle, S.Y.; Bitterman, D.S.; Thomas, S.; Griffin, K.L.; Atkin, O.K.; Turnbull, M.H. Respiratory alternative oxidase responds to both low- and high-temperature stress in Quercus rubra leaves along an urban-rural gradient in New York. Funct. Ecol 2011, 25, 1007–1017. [Google Scholar]
- Murakami, Y.; Toriyama, K. Enhanced high temperature tolerance in transgenic rice seedlings with elevated levels of alternative oxidase, OsAOX1a. Plant Biotechnol 2008, 25, 361–364. [Google Scholar]
- Flexas, J.; Bota, J.; Galmés, J.; Medrano, H.; Ribas-Carbo, M. Keeping a positive carbon balance under adverse conditions: Responses of photosynthesis and respiration to water stress. Physiol. Plant 2006, 127, 343–352. [Google Scholar]
- Atkin, O.K.; Macherel, D. The crucial role of plant mitochondria in orchestrating drought tolerance. Ann. Bot 2009, 103, 581–597. [Google Scholar]
- Lawlor, D.W.; Tezara, W. Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: A critical evaluation of mechanisms and integration of processes. Ann. Bot 2009, 103, 561–579. [Google Scholar]
- Bartoli, C.G.; Gomez, F.; Gergoff, G.; Guiamét, J.J.; Puntarulo, S. Up-Regulation of the mitochondrial alternative oxidase pathway enhances photosynthetic electron transport under drought conditions. J. Exp. Bot 2005, 56, 1269–1276. [Google Scholar]
- Vassileva, V.; Simova-Stoilova, L.; Demirevska, K.; Feller, U. Variety-specific response of wheat (Triticum aestivum, L.) leaf mitochondria to drought stress. J. Plant Res 2009, 122, 445–454. [Google Scholar]
- Ribas-Carbo, M.; Taylor, N.L.; Giles, L.; Busquets, S.; Finnegan, P.M.; Day, D.A.; Lambers, H.; Medrano, H.; Berry, J.A.; Flexas, J. Effects of water stress on respiration in soybean leaves. Plant Physiol 2005, 139, 466–473. [Google Scholar]
- Filippou, P.; Antoniou, C.; Fotopoulos, V. Effect of drought and rewatering on the cellular status and antioxidant response of Medicago trunculata plants. Plant Signal. Behav 2011, 6, 270–277. [Google Scholar]
- Galle, A.; Florez-Sarasa, I.; Thameur, A.; de Paepe, R.; Flexas, J.; Ribas-Carbo, M. Effects of drought stress and subsequent rewatering on photosynthesis and respiratory pathways in Nicotiana sylvestris wild type and the mitochondrial complex I-deficient CMSII. J. Exp. Bot 2010, 61, 765–775. [Google Scholar]
- Juszczuk, I.M.; Wagner, A.M.; Rychter, A.M. Regulation of alternative oxidase activity during phosphate deficiency in bean roots (Phaseolus vulgaris). Physiol. Plant 2001, 113, 185–192. [Google Scholar]
- Gonzàlez-Meler, M.A.; Giles, L.; Thomas, R.B.; Siedow, J.N. Metabolic regulation of leaf respiration and alternative oxidase activity in response to phosphate supply. Plant Cell Environ 2001, 24, 205–215. [Google Scholar]
- Parsons, H.L.; Yip, J.Y.H.; Vanlerberghe, G.C. Increased respiratory restriction during phosphate-limited growth in transgenic tobacco cells lacking alternative oxidase. Plant Physiol 1999, 121, 1309–1320. [Google Scholar]
- Vijayraghavan, V.; Soole, K. Effect of short- and long-term phosphate stress on the non-phosphorylating pathway of mitochondrial electron transport in Arabidopsis thaliana. Funct. Plant Biol 2010, 37, 455–466. [Google Scholar]
- Rychter, A.M.; Mikulska, M. The relationship between phosphate status and cyanide-resistant respiration in bean roots. Physiol. Plant 1990, 79, 663–667. [Google Scholar]
- Theodorou, M.E.; Plaxton, W.C. Metabolic adaptations of plant respiration to nutritional phosphate deprivation. Plant Physiol 1993, 101, 339–344. [Google Scholar]
- Sieger, S.M.; Kristensen, B.K.; Robson, C.A.; Amirsadeghi, S.; Eng, E.W.Y.; Abdel-Mesih, A.; Møller, I.M.; Vanlerberghe, G.C. The role of alternative oxidase in modulating carbon use efficiency and growth during macronutrient stress in tobacco cells. J. Exp. Bot 2005, 56, 1499–1515. [Google Scholar]
- Yip, J.Y.H.; Vanlerberghe, G.C. Mitochondrial alternative oxidase acts to dampen the generation of active oxygen species during a period of rapid respiration induced to support a high rate of nutrient uptake. Physiol. Plant 2001, 112, 327–333. [Google Scholar]
- Noguchi, K.; Terashima, I. Responses of spinach leaf mitochondria to low N availability. Plant Cell Environ 2006, 29, 710–719. [Google Scholar]
- Kornfeld, A.; Heskel, M.; Atkin, O.K.; Gough, L.; Griffin, K.L.; Horton, T.W.; Turnbull, M.H. Respiratory flexibility and efficiency are affected by simulated global change in Arctic plants. New Phytol 2013, 197, 1161–1172. [Google Scholar]
- Mittova, V.; Tal, M.; Volokita, M.; Guy, M. Up-regulation of the leaf mitochondrial and peroxisomal antioxidative systems in response to salt-induced oxidative stress in the wild salt-tolerant tomato species Lycopersicon pennellii. Plant Cell Environ 2003, 26, 845–856. [Google Scholar]
- Mittova, V.; Guy, M.; Tal, M.; Volokita, M. Salinity up-regulates the antioxidative system in root mitochondria and peroxisomes of the wild salt-tolerant tomato species Lycopersicon pennellii. J. Exp. Bot 2004, 55, 1105–1113. [Google Scholar]
- Chen, X.; Wang, Y.; Li, J.; Jiang, A.; Cheng, Y.; Zhang, W. Mitochondrial proteome during salt stress-induced programmed cell death in rice. Plant Physiol. Biochem 2009, 47, 407–415. [Google Scholar]
- Zsigmond, L.; Szepesi, A.; Tari, I.; Rigó, G.; Király, A.; Szabados, L. Overexpression of the mitochondrial PPR40 gene improves salt tolerance in Arabidopsis. Plant Sci 2012, 182, 87–93. [Google Scholar]
- Andronis, E.A.; Roubelakis-Angelakis, K.A. Short-Term salinity stress in tobacco leads to the onset of animal-like PCD hallmarks in planta in contrast to long-term stress. Planta 2010, 231, 437–448. [Google Scholar]
- Kreps, J.A.; Wu, Y.; Chang, H.-S.; Zhu, T.; Wang, X.; Harper, J.F. Transcriptome changes for Arabidopsis in response to salt, osmotic and cold stress. Plant Physiol 2002, 130, 2129–2141. [Google Scholar]
- Seki, M.; Narusaka, M.; Ishida, J.; Nanjo, T.; Fijita, M.; Oono, Y.; Kamiya, A.; Nakajima, M.; Enju, A.; Sakurai, T.; et al. Monitoring the expression profiles of 7000 Arabidopsis genes under drought, cold and high-salinity stresses using a full-length cDNA microarray. Plant J 2002, 31, 279–292. [Google Scholar]
- Marti, M.C.; Florez-Sarasa, I.; Camejo, D.; Ribas-Carbó, M.; Lázaro, J.J.; Sevilla, F.; Jiménez, A. Response of mitochondrial thioredoxin PsTrxo1, antioxidant enzymes, and respiration to salinity in pea (Pisum sativum, L.) leaves. J. Exp. Bot 2011, 62, 3863–3874. [Google Scholar]
- Ottow, E.A.; Brinker, M.; Teichmann, T.; Fritz, E.; Kaiser, W.; Brosché, M.; Kangasjärvi, J.; Jiang, X.; Polle, A. Populus euphratica displays apoplastic sodium accumulation, osmotic adjustment by decreases in calcium and soluble carbohydrates, and develops leaf succulence under salt stress. Plant Physiol 2005, 139, 1762–1772. [Google Scholar]
- Wang, H.; Liang, X.; Huang, J.; Zhang, D.; Lu, H.; Liu, Z.; Bi, Y. Involvement of ethylene and hydrogen peroxide in induction of alternative respiratory pathway in salt-treated Arabidopsis calluses. Plant Cell Physiol 2010, 51, 1754–1765. [Google Scholar]
- Ederli, L.; Morettini, R.; Borgogni, A.; Wasternack, C.; Miersch, O.; Reale, L.; Ferranti, F.; Tosti, N.; Pasqualini, S. Interaction between nitric oxide and ethylene in the induction of alternative oxidase in ozone-treated tobacco plants. Plant Physiol 2006, 142, 595–608. [Google Scholar]
- Pasqualini, S.; Reale, L.; Calderini, O.; Pagiotti, R.; Ederli, L. Involvement of protein kinases and calcium in the NO-signalling cascade for defence-gene induction in ozonated tobacco plants. J. Exp. Bot 2012, 63, 4485–4496. [Google Scholar]
- Tosti, N.; Pasqualini, S.; Borgogni, A.; Ederli, L.; Falistocco, E.; Crispi, S.; Paolocci, F. Gene expression profiles of O3-treated Arabidopsis plants. Plant Cell Environ. 2006, 29, 1686–1702. [Google Scholar]
- Kristensen, B.K.; Askerlund, P.; Bykova, N.V.; Egsgaard, H.; Møller, I.M. Identification of oxidized proteins in the matrix of rice leaf mitochondria by immunoprecipitation and two-dimensional liquid chromatography-tandem mass spectrometry. Phytochemistry 2004, 65, 1839–1851. [Google Scholar]
- Tan, Y.-F.; O’Toole, N.; Taylor, N.L.; Millar, A.H. Divalent metal ions in plant mitochondria and their role in interactions with proteins and oxidative stress-induced damage to respiratory function. Plant Physiol 2010, 152, 747–761. [Google Scholar]
- Yamamoto, Y.; Kobayashi, Y.; Devi, S.R.; Matsumoto, H. Aluminum toxicity is associated with mitochondrial dysfunction and the production of reactive oxygen species in plant cells. Plant Physiol 2002, 128, 63–72. [Google Scholar]
- Li, Z.; Xing, D. Mechanistic study of mitochondria-dependent programmed cell death induced by aluminum phytotoxicity using fluorescence techniques. J. Exp. Bot 2011, 62, 331–343. [Google Scholar]
- Panda, S.K.; Sahoo, L.; Katsuhara, M.; Matsumoto, H. Overexpression of alternative oxidase gene confers aluminum tolerance by altering the respiratory capacity and the response to oxidative stress in tobacco cells. Mol. Biotechnol. 2013. [Google Scholar] [CrossRef]
- Castro-Guerrero, N.A.; Rodríquez-Zavala, J.S.; Marín-Hernández, A.; Rodríguez-Enríquez, S.; Moreno-Sánchez, R. Enhanced alternative oxidase and antioxidant enzymes under Cd2+ stress in Euglena. J. Bioenerg. Biomembr 2008, 40, 227–235. [Google Scholar]
- Pádua, M.; Aubert, S.; Casimiro, A.; Bligny, R.; Millar, A.H.; Day, D.A. Induction of alternative oxidase by excess copper in sycamore cell suspensions. Plant Physiol. Biochem 1999, 37, 131–137. [Google Scholar]
- Amor, Y.; Chevion, M.; Levine, A. Anoxia pretreatment protects soybean cells against H2O2-induced cell death: possible involvement of peroxidases and of alternative oxidase. FEBS Lett 2000, 477, 175–180. [Google Scholar]
- Tsuji, H.; Nakazono, M.; Saisho, D.; Tsutsumi, N.; Hirai, A. Transcript levels of the nuclear-encoded respiratory genes in rice decrease by oxygen deprivation: Evidence for involvement of calcium in expression of the alternative oxidase 1a gene. FEBS Lett 2000, 471, 201–204. [Google Scholar]
- Klok, E.J.; Wilson, I.W.; Wilson, D.; Chapman, S.C.; Ewing, R.M.; Somerville, S.C.; Peacock, W.J.; Dolferus, R.; Dennis, E.S. Expression profile analysis of the low-oxygen response in Arabidopsis root cultures. Plant Cell 2002, 14, 2481–2494. [Google Scholar]
- Szal, B.; Jolivet, Y.; Hasenfratz-Sauder, M.-P.; Dizengremel, P.; Rychter, A.M. Oxygen concentration regulates alternative oxidase expression in barley roots during hypoxia and post-hypoxia. Physiol. Plant 2003, 119, 494–502. [Google Scholar]
- Liu, F.; VanToai, T.; Moy, L.P.; Bock, G.; Linford, L.D.; Quackenbush, J. Global transcription profiling revelas comprehensive insights into hypoxic response in Arabidopsis. Plant Physiol 2005, 137, 1115–1129. [Google Scholar]
- Kreuzwieser, J.; Hauberg, J.; Howell, K.A.; Carroll, A.; Rennenberg, H.; Millar, A.H.; Whelan, J. Differential response of gray poplar leaves and roots underpins stress adaptation during hypoxia. Plant Physiol 2009, 149, 461–473. [Google Scholar]
- Skutnik, M.; Rychter, A.M. Differential response of antioxidant systems in leaves and roots of barley subjected to anoxia and post-anoxia. J. Plant Physiol 2009, 166, 926–937. [Google Scholar]
- Vergara, R.; Parada, F.; Rubio, S.; Pérez, J. Hypoxia induces H2O2 production and activates antioxidant defence system in grapevine buds through mediation of H2O2 and ethylene. J. Exp. Bot 2012, 63, 4123–4131. [Google Scholar]
- Millar, A.H.; Bergersen, F.J.; Day, D.A. Oxygen affinity of terminal oxidases in soybean mitochondria. Plant Physiol. Biochem 1994, 32, 847–852. [Google Scholar]
- Ribas-Carbo, M.; Berry, J.A.; Azcon-Bieto, J.; Siedow, J.N. The reaction of the plant mitochondrial cyanide-resistant alternative oxidase with oxygen. Biochim. Biophys. Acta 1994, 118, 205–212. [Google Scholar]
- Leitner, M.; Vandelle, E.; Gaupels, F.; Bellin, D.; Delledonne, M. NO signals in the haze. Nitric oxide signaling in plant defense. Curr. Opin. Plant Biol 2009, 12, 451–458. [Google Scholar]
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol 2009, 47, 177–206. [Google Scholar]
- Spoal, S.H.; Loake, G.J. Redox-Based protein modifications: The missing link in plant immune signaling. Curr. Opin. Plant Biol 2011, 14, 358–364. [Google Scholar]
- Norman, C.; Howell, K.A.; Millar, A.H.; Whelan, J.M.; Day, D.A. Salicylic acid is an uncoupler and inhibitor of mitochondrial electron transport. Plant Physiol 2004, 134, 492–501. [Google Scholar]
- Millar, A.H.; Day, D.A. Nitric oxide inhibits the cytochrome oxidase but not the alternative oxidase on plant mitochondria. FEBS Lett 1996, 398, 155–158. [Google Scholar]
- Lacomme, C.; Roby, D. Identification of new early markers of the hypersensitive response in Arabidopsis thaliana. FEBS Lett 1999, 459, 149–153. [Google Scholar]
- Simons, B.H.; Millenaar, F.F.; Mulder, L.; van Loon, L.C.; Lambers, H. Enhanced expression and activation of the alternative oxidase during infection of Arabidopsis with Pseudomonas syringae pv. tomato. Plant Physiol 1999, 120, 529–538. [Google Scholar]
- Amirsadeghi, S.; Robson, C.A.; Vanlerberghe, G.C. The role of the mitochondrion in plant responses to biotic stress. Physiol. Plant 2007, 129, 253–266. [Google Scholar]
- Sun, A.; Nie, S.; Xing, D. Nitric oxide-mediated maintenance of redox homeostasis contributes to NPR1-dependent plant innate immunity triggered by lipopolysaccharides. Plant Physiol 2012, 160, 1081–1096. [Google Scholar]
- Chivasa, S.; Murphy, A.M.; Naylor, M.; Carr, J.P. Salicylic acid interferes with Tobacco mosaic virus replication via a novel salicylhydroxamic acid-sensitive mechanism. Plant Cell 1997, 9, 547–557. [Google Scholar]
- Ordog, S.H.; Higgins, V.J.; Vanlerberghe, G.C. Mitochondrial alternative oxidase is not a critical component of plant viral resistance but may play a role in the hypersensitive response. Plant Physiol 2002, 129, 1858–1865. [Google Scholar]
- Gilliland, A.; Singh, D.P.; Hayward, J.M.; Moore, C.A.; Murphy, A.M.; York, C.J.; Slator, J.; Carr, J.P. Genetic modification of alternative respiration has differential effects on antimycin A-induced versus salicylic acid-induced resistance to Tobacco mosaic virus. Plant Physiol 2003, 132, 1518–1528. [Google Scholar]
- Dutilleul, C.; Garmier, M.; Noctor, G.; Mathieu, C.; Chétrit, P.; Foyer, C.H.; de Paepe, R. Leaf mitochondria modulate whole cell redox homeostasis, set antioxidant capacity, and determine stress resistance through altered signaling and diurnal regulation. Plant Cell 2003, 15, 1212–1226. [Google Scholar]
- Király, L.; Künstler, A.; Höller, K.; Fattinger, M.; Juhász, C.; Müller, M.; Gullner, G.; Zechmann, B. Sulfate supply influences compartment specific glutathione metabolism and confers enhanced resistance to Tobacco mosaic virus during a hypersensitive response. Plant Physiol. Biochem 2012, 59, 44–54. [Google Scholar]
- Zhu, L.; Li, Y.; Li, L.; Yang, J.; Zhang, M. Ethylene is involved in leafy mustard systemic resistance to Turnip mosaic virus infection through the mitochondrial alternative oxidase pathway. Physiol. Mol. Plant Pathol 2011, 76, 166–172. [Google Scholar]
- Liao, Y.-W.-K.; Shi, K.; Fu, L.-J.; Zhang, S.; Li, X.; Dong, D.-K.; Jiang, Y.-P.; Zhou, Y.-H.; Xia, X.-J.; Liang, W.-S.; et al. The reduction of reactive oxygen species formation by mitochondrial alternative respiration in tomato basal defense against TMV infection. Planta 2012, 235, 225–238. [Google Scholar]
- Norman, E.G.; Walton, A.B.; Turpin, D.H. Immediate activation of respiration in Petroselinum crispum, L. in response to the Phytophthora megasperma f. sp. Glycinea elicitor. Plant Physiol 1994, 106, 1541–1546. [Google Scholar]
- Naton, B.; Hahlbrock, K.; Schmelzer, E. Correlation of rapid cell death with metabolic changes in fungus-infected, cultured parsley cells. Plant Physiol 1996, 112, 433–444. [Google Scholar]
- Scharte, J.; Schön, H.; Weis, E. Photosynthesis and carbohydrate metabolism in tobacco leaves during an incompatible interaction with Phytophthora. nicotianae. Plant Cell Environ 2005, 28, 1421–1435. [Google Scholar]
- Manzoor, H.; Chiltz, A.; Madani, S.; Vatsa, P.; Schoefs, B.; Pugin, A.; Garcia-Brugger, A. Calcium signatures and signaling in cytosol and organelles of tobacco cells induced by plant defense elicitors. Cell Calcium 2012, 51, 434–444. [Google Scholar]
- Ashtamker, C.; Kiss, V.; Sagi, M.; Davydov, O.; Fluhr, R. Diverse subcellular locations of cryptogein-induced reactive oxygen species production in tobacco Bright Yellow-2 cells. Plant Physiol 2007, 143, 1817–1826. [Google Scholar]
- Yao, N.; Tada, Y.; Sakamoto, M.; Nakayashiki, H.; Park, P.; Tosa, Y.; Mayama, S. Mitochondrial oxidative burst involved in apoptotic response in oats. Plant J 2002, 30, 567–579. [Google Scholar]
- Seo, S.; Mitsuhara, I.; Feng, J.; Iwai, T.; Hasegawa, M.; Ohashi, Y. Cyanide, a coproduct of plant hormone ethylene biosynthesis, contributes to the resistance of rice to blast fungus. Plant Physiol 2011, 155, 502–514. [Google Scholar]
- Astier, J.; Besson-Bard, A.; Lamotte, O.; Bertoldo, J.; Bourque, S.; Terenz, H.; Wendehenne, D. Nitric oxide inhibits the ATPase activity of the chaperone-like AAA+ ATPase CDC48, a target for S-nitrosylation in cryptogein signalling in tobacco cells. Biochem. J 2012, 447, 249–260. [Google Scholar]
- Hoefnagel, M.H.N.; Atkin, O.K.; Wiskich, J.T. Interdependence between chloroplasts and mitochondria in the light and the dark. Biochim. Biophys. Acta 1998, 1366, 235–255. [Google Scholar]
- Raghavendra, A.S.; Padmasree, K. Beneficial interactions of mitochondrial metabolism with photosynthetic carbon assimilation. Trends Plant Sci 2003, 8, 546–553. [Google Scholar]
- Noctor, G.; Dutilleul, C.; de Paepe, R.; Foyer, C.H. Use of mitochondrial electron transport mutants to evaluate the effects of redox state on photosynthesis, stress tolerance and the integration of carbon/nitrogen metabolism. J. Exp. Bot 2004, 55, 49–57. [Google Scholar]
- Noguchi, K.; Yoshida, K. Interaction between photosynthesis and respiration in illuminated leaves. Mitochondrion 2008, 8, 87–99. [Google Scholar]
- Yoshida, K.; Terashima, I.; Noguchi, K. Distinct roles of the cytochrome pathway and alternative oxidase in leaf photosynthesis. Plant Cell Physiol 2006, 47, 22–31. [Google Scholar]
- Dinakar, C.; Abhaypratap, V.; Yearla, S.R.; Raghavendra, A.S.; Padmasree, K. Importance of ROS and antioxidant system during the beneficial interactions of mitochondrial metabolism with photosynthetic carbon assimilation. Planta 2010, 231, 461–474. [Google Scholar]
- Dinakar, C.; Raghavendra, A.S.; Padmasree, K. Importance of AOX pathway in optimizing photosynthesis under high light stress: Role of pyruvate and malate in activating AOX. Physiol. Plant 2010, 139, 13–26. [Google Scholar]
- Zhang, L.-T.; Zhang, Z.-S.; Gao, H.-Y.; Xue, Z.-C.; Yang, C.; Meng, X.-L. Mitochondrial alternative oxidase pathway protects plants against photoinhibition by alleviating inhibition of the repair of photodamaged PSII through preventing formation of reactive oxygen species in Rumex. K-1 leaves. Physiol. Plant 2011, 143, 396–407. [Google Scholar]
- Yoshida, K.; Noguchi, K. Differential gene expression profiles of the mitochondrial respiratory components in illuminated Arabidopsis leaves. Plant Cell Physiol 2009, 50, 1449–1462. [Google Scholar]
- Yoshida, K.; Watanabe, C.K.; Hachiya, T.; Tholen, D.; Shibata, M.; Terashima, I.; Noguchi, K. Distinct responses of the mitochondrial respiratory chain to long- and short-term high-light environments in Arabidopsis thaliana. Plant Cell Environ 2011, 34, 618–628. [Google Scholar]
- Noguchi, K.; Taylor, N.L.; Millar, A.H.; Lambers, H.; Day, D.A. Response of mitochondria to light intensity in the leaves of sun and shade species. Plant Cell Environ 2005, 28, 760–771. [Google Scholar]
- Pastore, D.; Trono, D.; Laus, M.N.; di Fonzo, N.; Passarella, S. Alternative oxidase in durum wheat mitochondria. Activation by pyruvate, hydroxypyruvate and glyoxylate and physiological role. Plant Cell Physiol 2001, 42, 1373–1382. [Google Scholar]
- Noguchi, K.; Go, C.-S.; Terashima, I.; Ueda, S.; Yoshinari, T. Activities of the cyanide-resistant respiratory pathway in leaves of sun and shade species. Austr. J. Plant Physiol 2001, 28, 27–35. [Google Scholar]
- Yoshida, K.; Terashima, I.; Noguchi, K. Up-regulation of mitochondrial alternative oxidase concomitant with chloroplast over-reduction by excess light. Plant Cell Physiol 2007, 48, 606–614. [Google Scholar]
- Rosso, D.; Bode, R.; Li, W.; Krol, M.; Saccon, D.; Wang, S.; Schillaci, L.A.; Rodermel, S.R.; Maxwell, D.P.; Hüner, N.P.A. Photosynthetic redox imbalance governs leaf sectoring in the Arabidopsis thaliana variegation mutants immutans, spotty, var1, and var2. Plant Cell 2009, 21, 3473–3492. [Google Scholar]
- Fu, A.; Liu, H.; Yu, F.; Kambakam, S.; Luan, S.; Rodermel, S. Alternative oxidases (AOX1a and AOX2) can functionally substitute for plastid terminal oxidase in Arabidopsis chloroplasts. Plant Cell 2012, 24, 1579–1595. [Google Scholar]
- Umbach, A.L.; Fiorani, F.; Siedow, J.N. Characterization of transformed Arabidopsis with altered alternative oxidase levels and analysis of effects on reactive oxygen species in tissue. Plant Physiol 2005, 139, 1806–1820. [Google Scholar]
- Zhang, D.-W.; Xu, F.; Zhang, Z.-W.; Chen, Y.-E.; Du, J.-B.; Jia, S.-D.; Yuan, S.; Lin, H.-H. Effects of light on cyanide-resistant respiration and alternative oxidase function in Arabidopsis seedlings. Plant Cell Environ 2010, 33, 2121–2131. [Google Scholar]
- Zidenga, T.; Leyva-Guerrero, E.; Moon, H.; Siritunga, D.; Sayre, R. Extending cassava root shelf life via reduction of reactive oxygen species production. Plant Physiol 2012, 159, 1396–1407. [Google Scholar]
- García, I.; Castellano, J.M.; Vioque, B.; Solano, R.; Gotor, C.; Romero, L.C. Mitochondrial β-cyanoalanine synthase is essential for root hair formation in Arabidopsis thaliana. Plant Cell 2010, 22, 3268–3279. [Google Scholar]
- Álvarez, C.; García, I.; Romero, L.C.; Gotor, C. Mitochondrial sulfide detoxification requires a functional isoform O-acetylserine(thiol)lyase C in Arabidopsis thaliana. Mol. Plant 2012, 5, 1217–1226. [Google Scholar]
- Gapper, C.; Dolan, L. Control of plant development by reactive oxygen species. Plant Physiol 2006, 141, 341–345. [Google Scholar]
- Blom, D.; Fabbri, C.; Eberl, L.; Weisskopf, L. Volatile-Mediated killing of Arabidopsis thaliana by bacteria is mainly due to hydrogen cyanide. Appl. Environ. Microbiol 2011, 77, 1000–1008. [Google Scholar]
- Pasqualini, S.; Paolocci, F.; Borgogni, A.; Morettini, R.; Ederli, L. The overexpression of an alternative oxidase gene triggers ozone sensitivity in tobacco plants. Plant Cell Environ 2007, 30, 1545–1556. [Google Scholar]
- Zhang, L.; Oh, Y.; Li, H.; Baldwin, I.T.; Galis, I. Alternative oxidase in resistance to biotic stresses: Nicotiana. attenuata AOX contributes to resistance to a pathogen and a piercing-sucking insect but not Manduca. sexta larvae. Plant Physiol 2012, 160, 1453–1467. [Google Scholar]
- Mathy, G.; Cardol, P.; Dinant, M.; Blomme, A.; Gérin, S.; Cloes, M.; Ghysels, B.; DePauw, E.; Leprince, P.; Remacle, C.; et al. Proteomic and functional characterization of a Chlamydomonas. reinhardtii mutant lacking the mitochondrial alternative oxidase 1. J. Proteome Res 2010, 9, 2825–2838. [Google Scholar]
- Millenaar, F.F.; Gonzàlez-Meler, M.A.; Fiorani, F.; Welschen, R.; Ribas-Carbo, M.; Siedow, J.N.; Wagner, A.M.; Lambers, H. Regulation of alternative oxidase in six wild monocotyledonous species. An in vivo study at the whole root level. Plant Physiol 2001, 126, 376–387. [Google Scholar]
- Florez-Sarasa, I.D.; Bouma, T.J.; Medrano, H.; Azcon-Bieto, J.; Ribas-Carbo, M. Contribution of the cytochrome and alternative pathways to growth respiration and maintenance respiration in Arabidopsis thaliana. Physiol. Plant 2007, 129, 143–151. [Google Scholar]
- Chai, T.-T.; Simmonds, D.; Day, D.A.; Colmer, T.D.; Finnegan, P.M. A GmAOX2b antisense gene compromises vegetative growth and seed production in soybean. Planta 2012, 236, 199–207. [Google Scholar]
- Skirycz, A.; de Bodt, S.; Obata, T.; de Clercq, I.; Claeys, H.; de Rycke, R.; Andriankaja, M.; van Aken, O.; van Breusegem, F.; Fernie, A.R.; et al. Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves to prolonged osmotic stress. Plant Physiol 2010, 152, 226–244. [Google Scholar]
- Shiba, T.; Kido, Y.; Sakamoto, K.; Inaoka, D.K.; Tsuge, C.; Tatsumi, R.; Takahashi, G.; Balogun, E.O.; Nara, T.; Aoki, T.; et al. Structure of the trypanosome cyanide-insensitive alternative oxidase. Proc. Natl. Acad. Sci. USA 2013. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Vanlerberghe, G.C. Alternative Oxidase: A Mitochondrial Respiratory Pathway to Maintain Metabolic and Signaling Homeostasis during Abiotic and Biotic Stress in Plants. Int. J. Mol. Sci. 2013, 14, 6805-6847. https://doi.org/10.3390/ijms14046805
Vanlerberghe GC. Alternative Oxidase: A Mitochondrial Respiratory Pathway to Maintain Metabolic and Signaling Homeostasis during Abiotic and Biotic Stress in Plants. International Journal of Molecular Sciences. 2013; 14(4):6805-6847. https://doi.org/10.3390/ijms14046805
Chicago/Turabian StyleVanlerberghe, Greg C. 2013. "Alternative Oxidase: A Mitochondrial Respiratory Pathway to Maintain Metabolic and Signaling Homeostasis during Abiotic and Biotic Stress in Plants" International Journal of Molecular Sciences 14, no. 4: 6805-6847. https://doi.org/10.3390/ijms14046805