The Role of E3 Ubiquitin Ligase Cbl Proteins in β-Elemene Reversing Multi-Drug Resistance of Human Gastric Adenocarcinoma Cells

Abstract
:1. Introduction
2. Results and Discussion
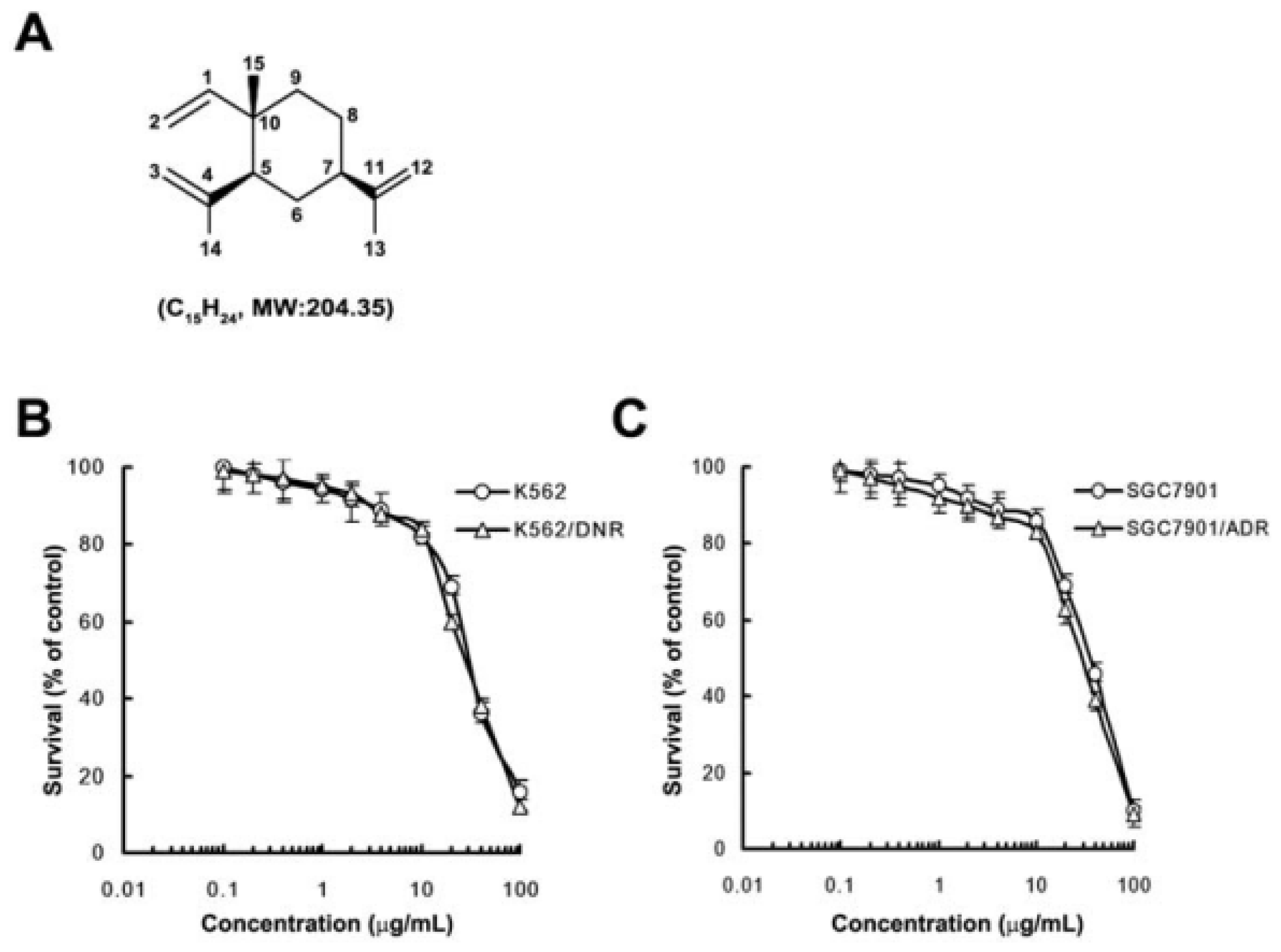
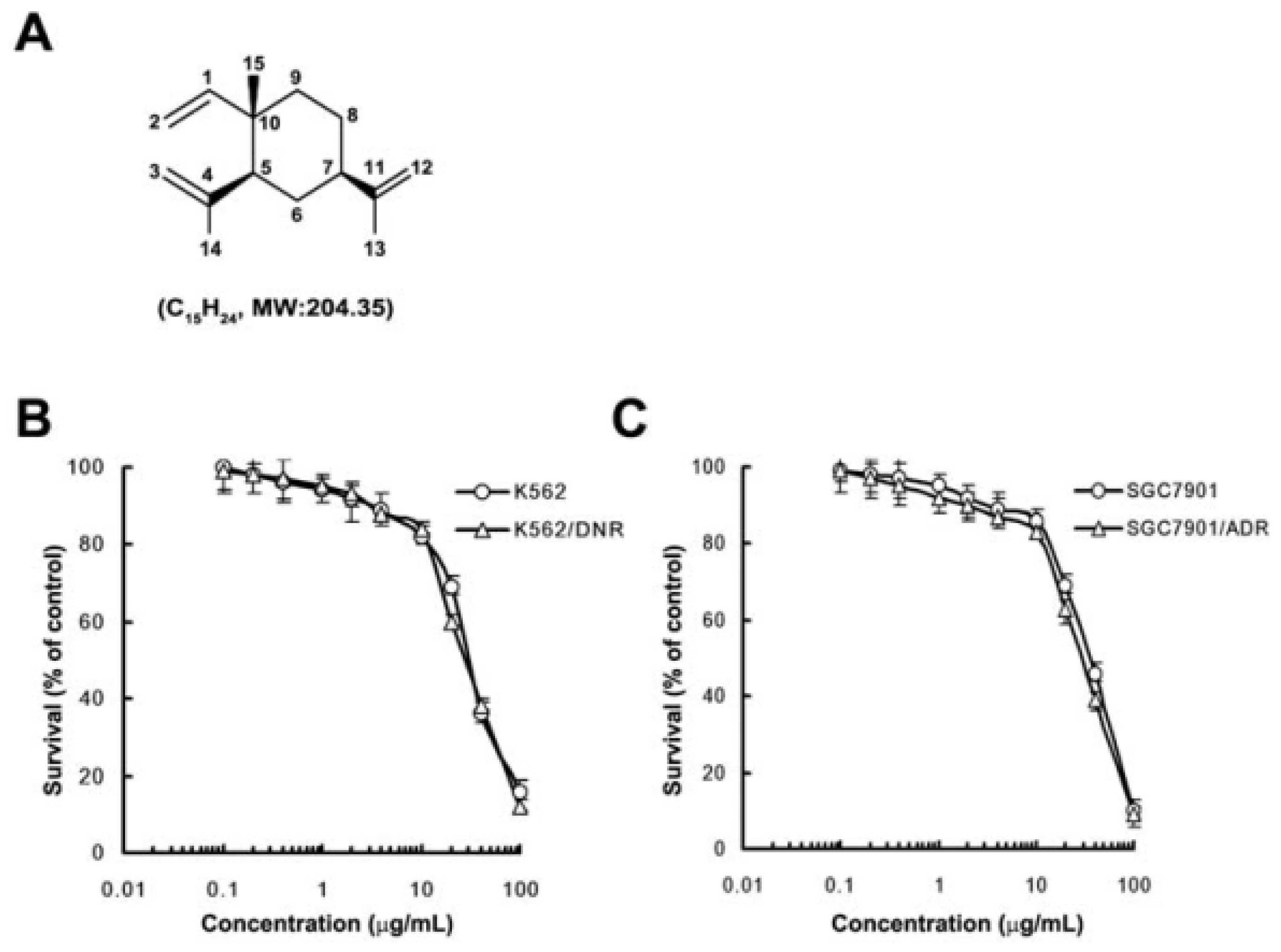
2.1. In Vitro Cytotoxicity of β-Elemene
2.2. β-Elemene Reverses P-gp-Mediated MDR in Vitro
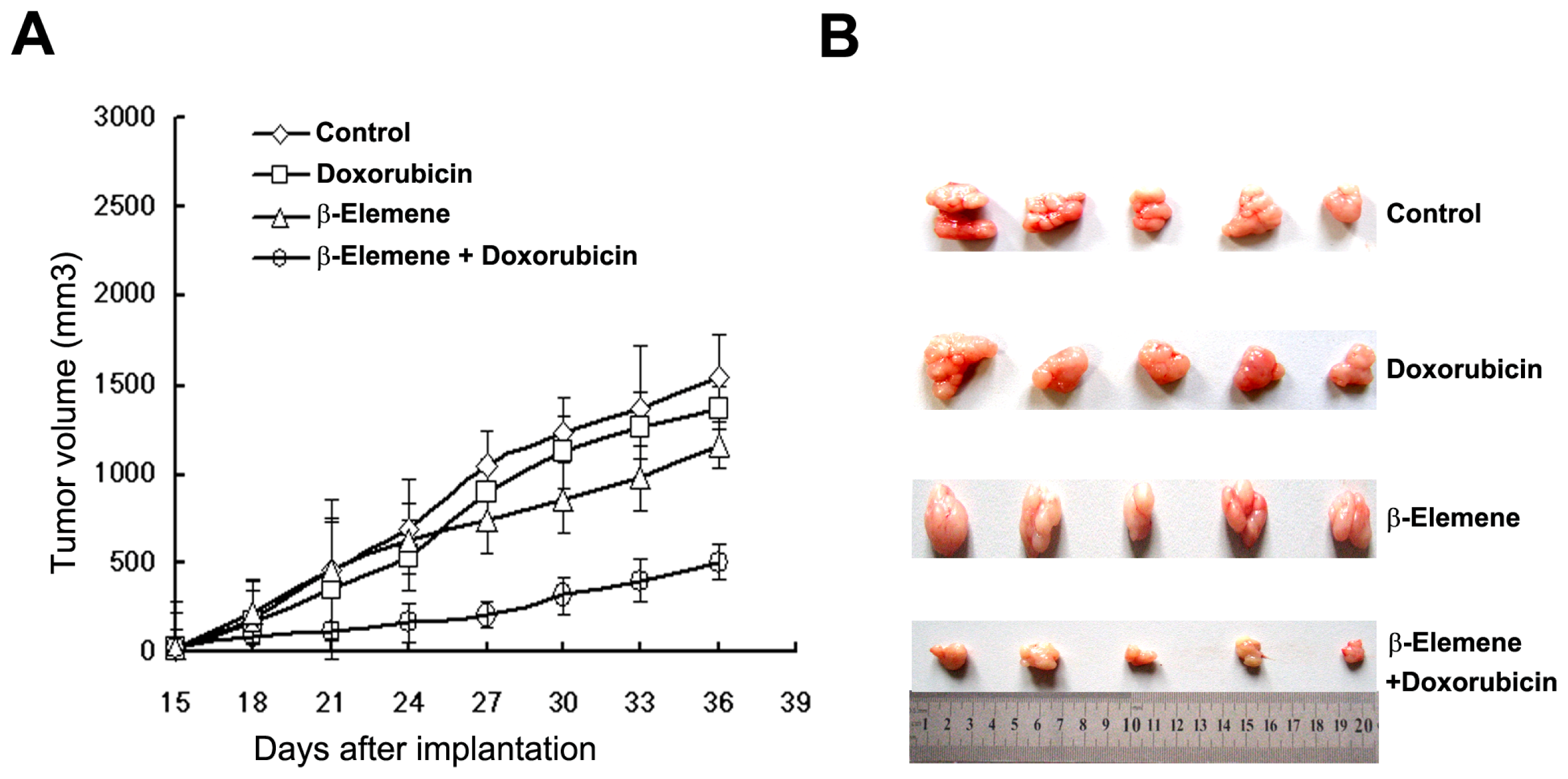
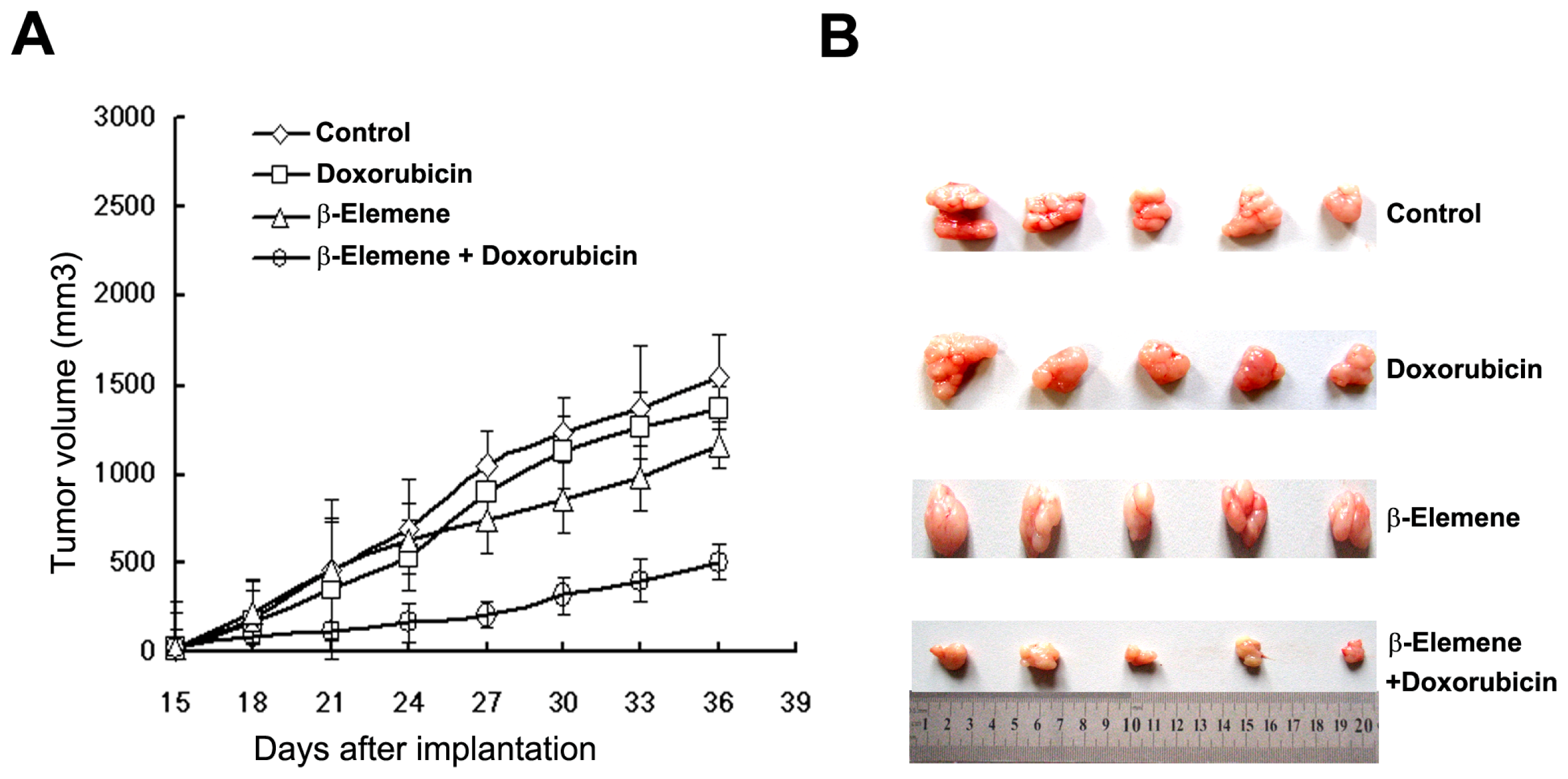
2.3. β-Elemene Reverses P-gp-Mediated MDR in Vivo
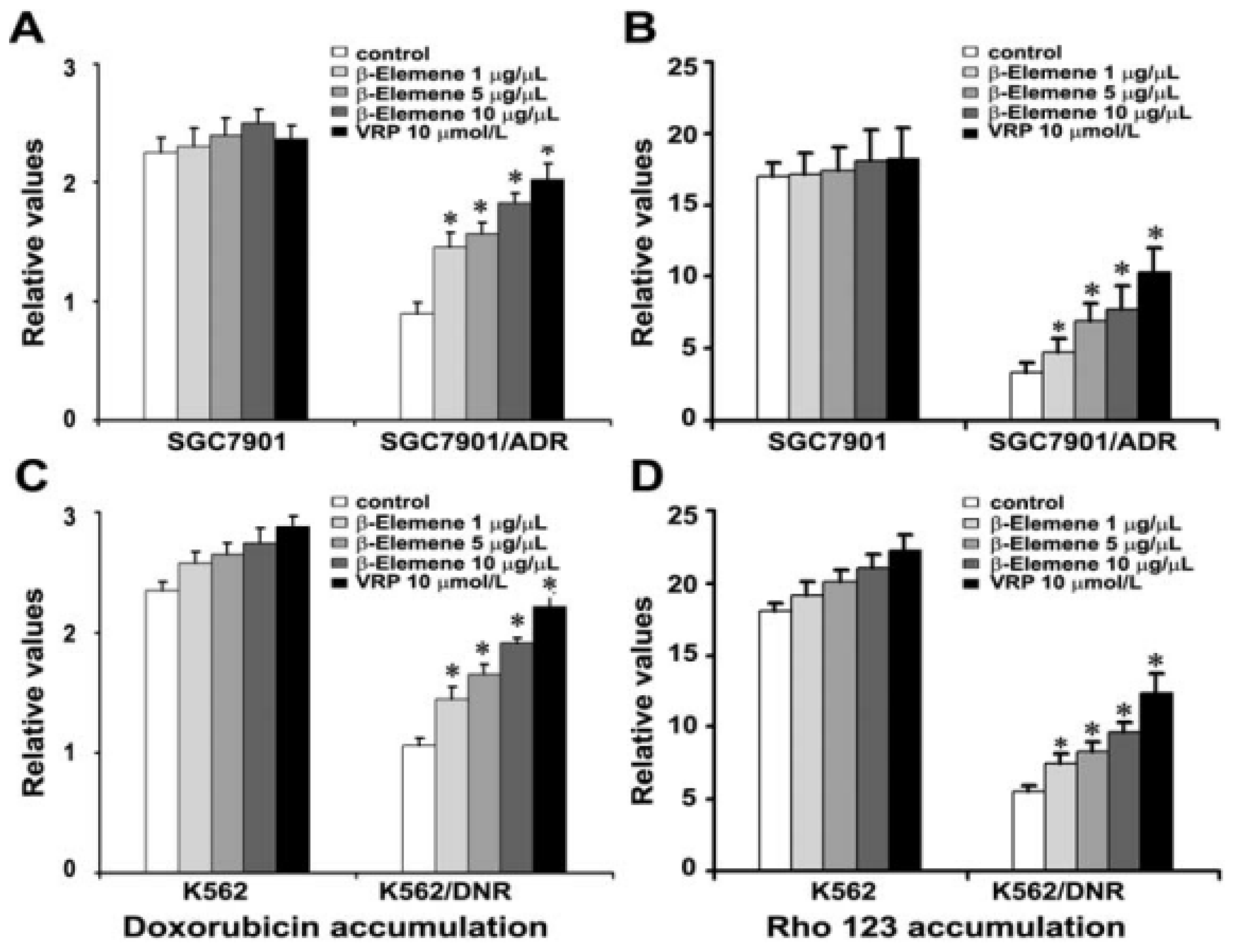
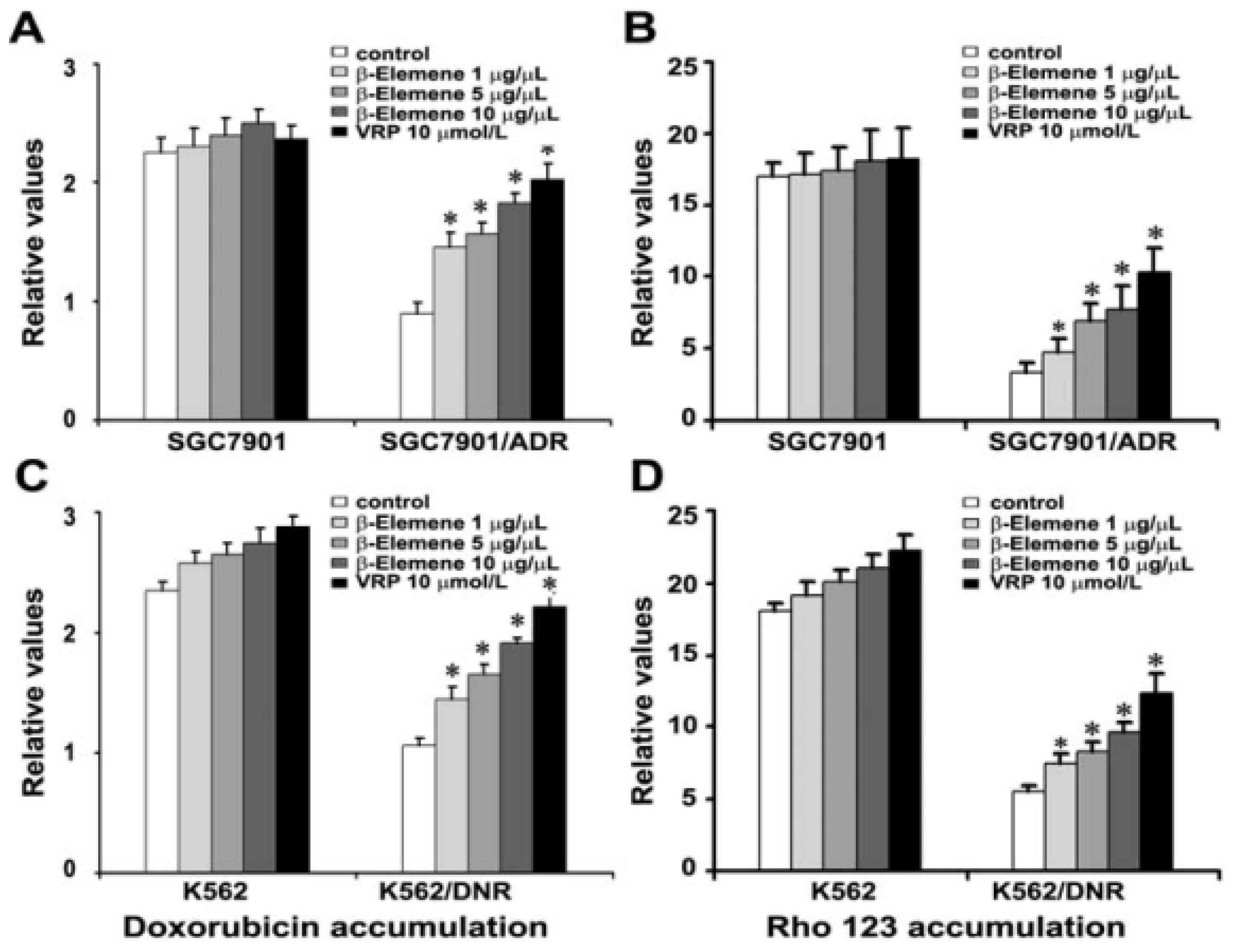
2.4. β-Elemene Increases the Accumulation of Rho 123 and DOX in Both K562/DNR and SGC7901/ADR Cells
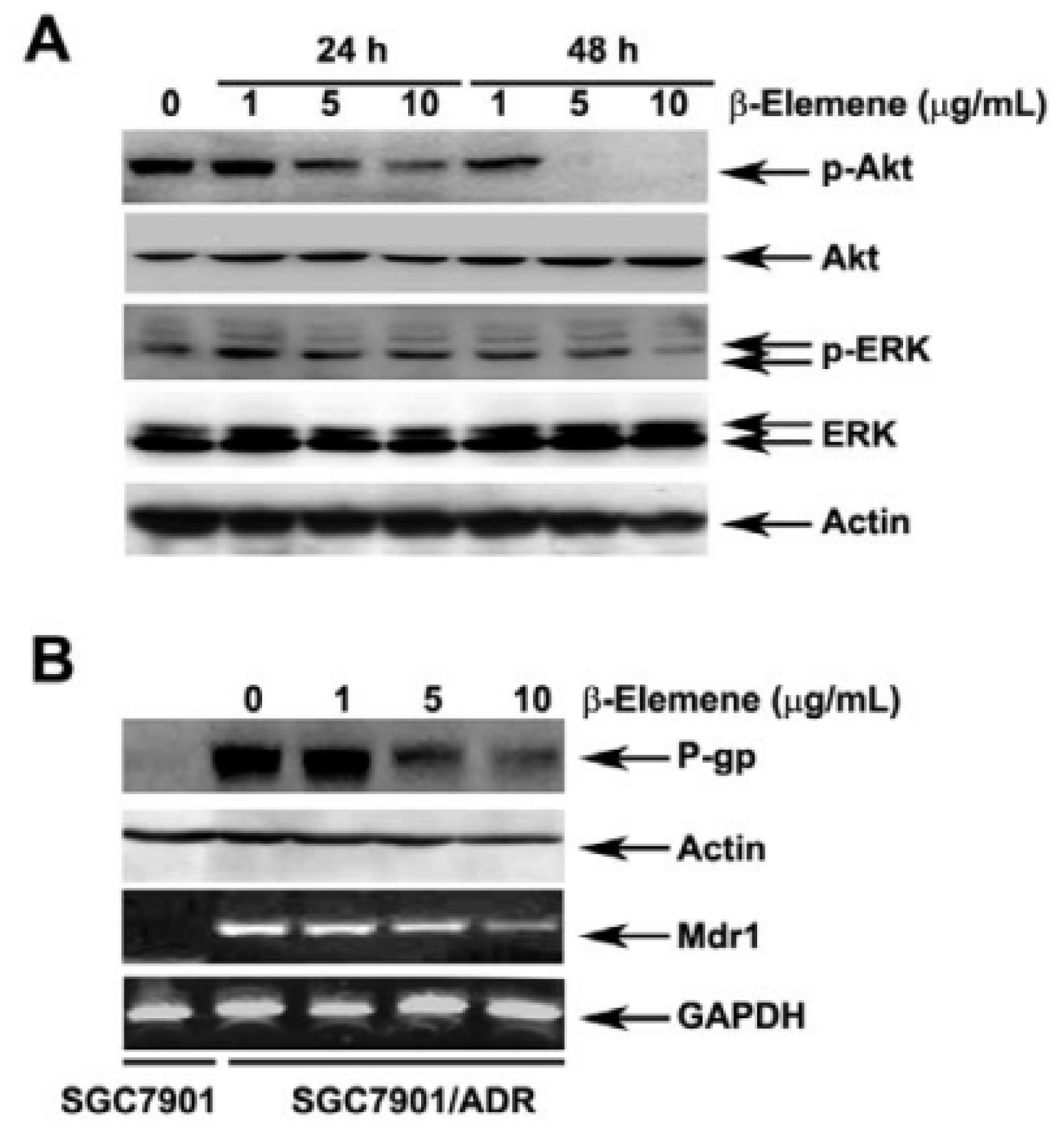
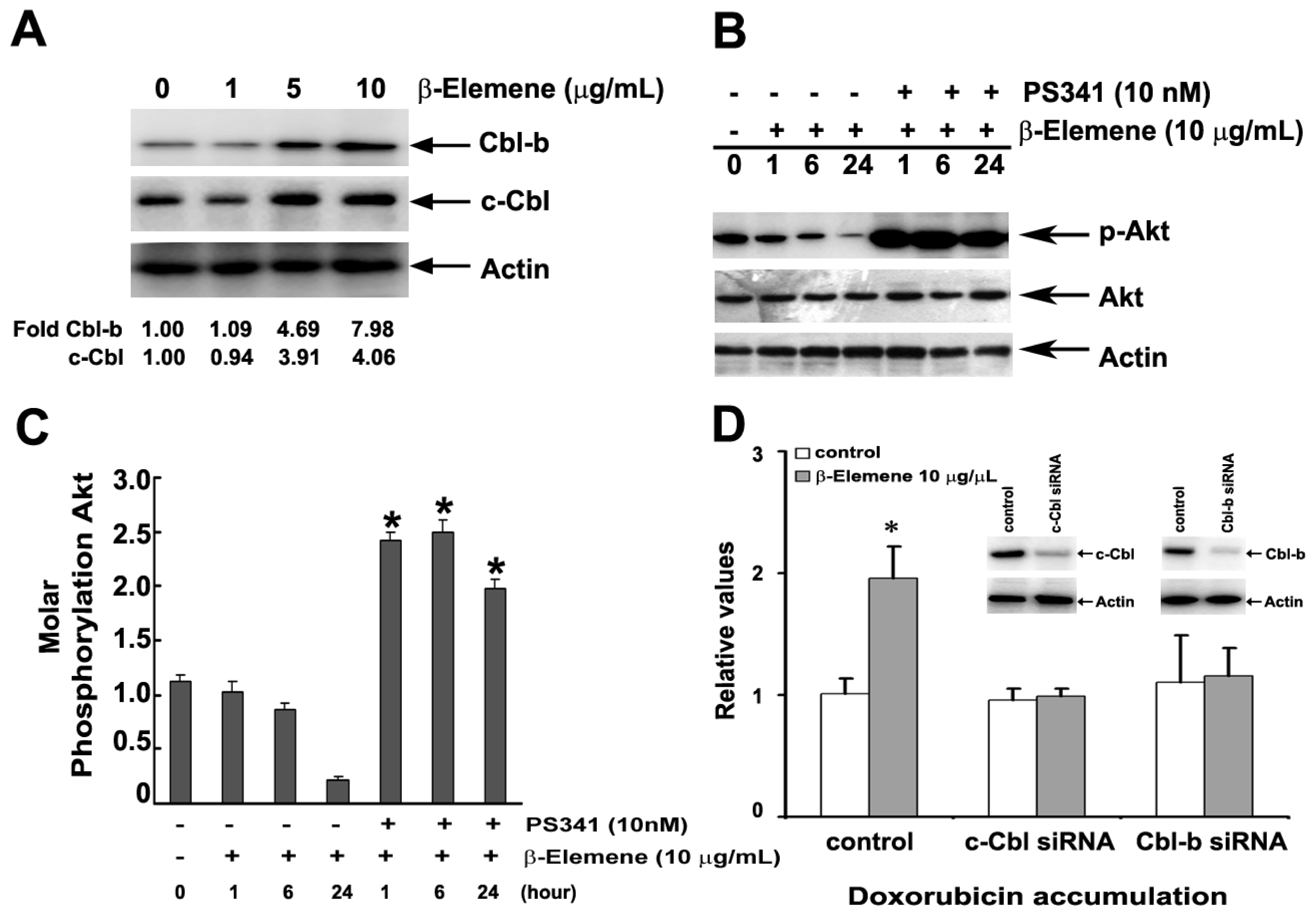
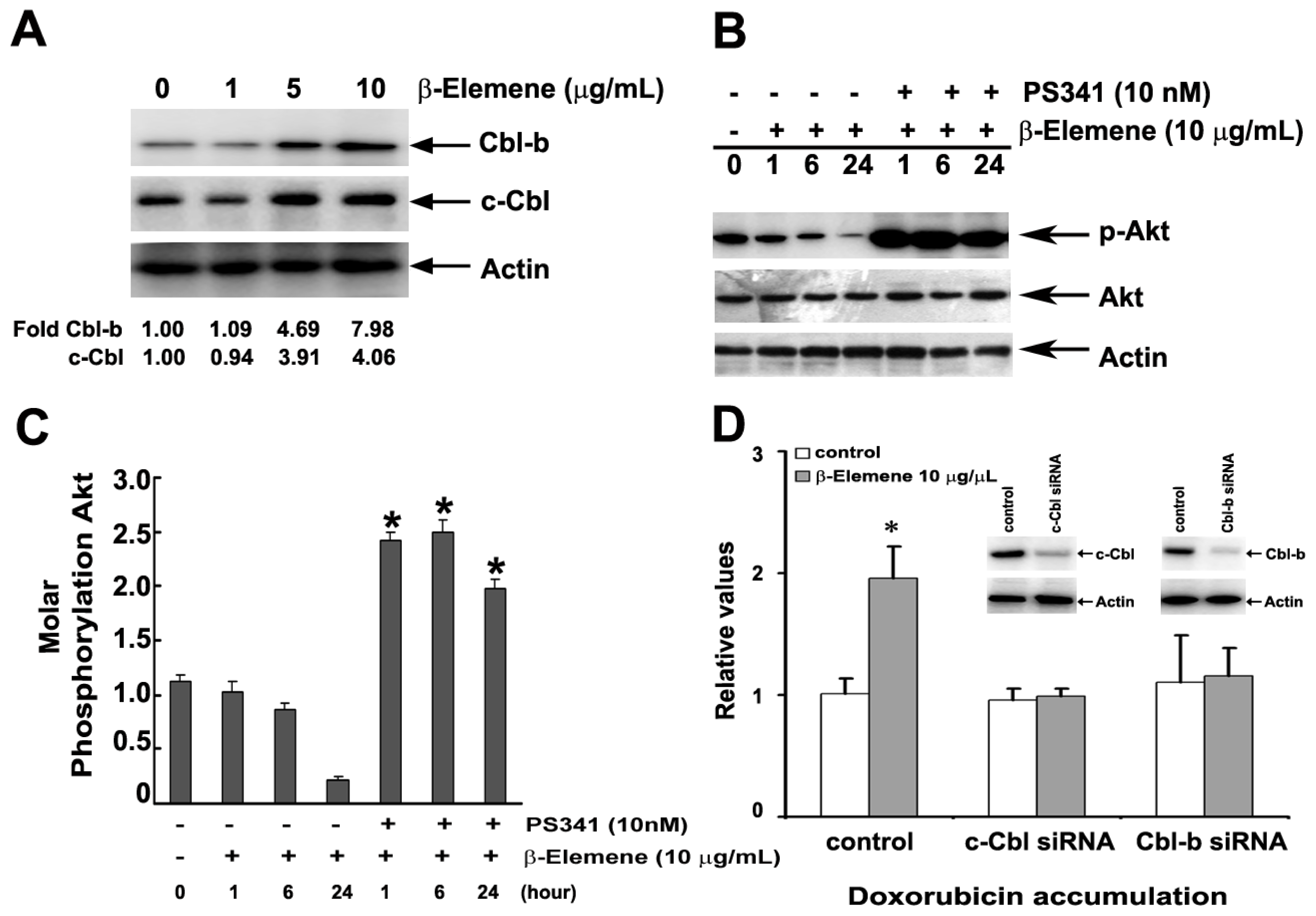
2.5. Effect of β-Elemene on the Inhibition of Akt Phosphorylation and Expression of P-gp
2.6. Effect of β-Elemene on the Up-Regulation of c-Cbl and Cbl-b Expression
2.7. Discussion
3. Experimental Section
3.1. Reagents
3.2. Cell Lines and Culture
3.3. Cell Viability Assay
3.4. Animal and Tumor Xenograft Experiments
3.5. DOX and Rho 123 Accumulation
3.6. Small Interfering RNA Transfections
3.7. Western Blot Analysis
3.8. Reverse-Transcription Polymerase Chain Reaction (RT-PCR)
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Sui, H.; Fan, Z.Z.; Li, Q. Signal transduction pathways and transcriptional mechanisms of ABCB1/Pgp-mediated multiple drug resistance in human cancer cells. J. Int. Med. Res 2012, 40, 426–435. [Google Scholar]
- Wu, H.; Kang, H.; Liu, Y.; Tong, W.; Liu, D.; Yang, X.; Lian, M.; Yao, W.; Zhao, H.; Huang, D.; et al. Roles of ABCB1 gene polymorphisms and haplotype in susceptibility to breast carcinoma risk and clinical outcomes. J. Cancer Res. Clin. Oncol 2012, 138, 1449–1462. [Google Scholar]
- Amiri-Kordestani, L.; Basseville, A.; Kurdziel, K.; Fojo, A.T.; Bates, S.E. Targeting MDR in breast and lung cancer: Discriminating its potential importance from the failure of drug resistance reversal studies. Drug Resist. Updat 2012, 15, 50–61. [Google Scholar]
- De Moraes, A.C.; Licínio, M.A.; Zampirolo, J.A.; Liedke, S.C.; Del Moral, J.A.; Machado, M.J.; Bazzo, M.L.; da Silva, M.C. Evaluation of multidrug resistance in 46 newly diagnosed patients with acute leukemia. Hematology 2012, 17, 59–65. [Google Scholar]
- Robey, R.W.; Massey, P.R.; Amiri-Kordestani, L.; Bates, S.E. ABC transporters: Unvalidated therapeutic targets in cancer and the CNS. Anticancer Agents Med. Chem 2010, 10, 625–633. [Google Scholar]
- Yu, Z.; Wang, R.; Xu, L.; Xie, S.; Dong, J.; Jing, Y. β-Elemene piperazine derivatives induce apoptosis in human leukemia cells through downregulation of c-FLIP and generation of ROS. PLoS One 2011, 6, e15843. [Google Scholar]
- Li, Q.Q.; Wang, G.; Huang, F.; Banda, M.; Reed, E. Antineoplastic effect of beta-elemene on prostate cancer cells and other types of solid tumour cells. J. Pharm. Pharmacol 2010, 62, 1018–1027. [Google Scholar]
- Li, L.J.; Zhong, L.F.; Jiang, L.P.; Geng, C.Y.; Zou, L.J. β-Elemene radiosensitizes lung cancer A549 cells by enhancing DNA damage and inhibiting DNA repair. Phytother. Res 2011, 25, 1095–1097. [Google Scholar]
- Li, L.J.; Zhong, L.F.; Jiang, L.P.; Geng, C.Y.; Zhu, T.Z.; Xu, Y.H.; Wang, Q.; Qu, Y.; Shao, J.; Zou, L.J. Lysosomal membrane permeabilization contributes to elemene emulsion-induced apoptosis in A549 cells. Free Radic. Res 2011, 45, 1232–1240. [Google Scholar]
- Lee, R.X.; Li, Q.Q.; Reed, E. β-elemene effectively suppresses the growth and survival of both platinum-sensitive and -resistant ovarian tumor cells. Anticancer Res 2012, 32, 3103–3113. [Google Scholar]
- Zhan, Y.H.; Liu, J.; Qu, X.J.; Hou, K.Z.; Wang, K.F.; Liu, Y.P.; Wu, B. β-Elemene induces apoptosis in human renal-cell carcinoma 786-0 cells through inhibition of MAPK/ERK and PI3K/Akt/mTOR signalling pathways. Asian Pac. J. Cancer Prev 2012, 13, 2739–2744. [Google Scholar]
- Chen, X.; Wang, Y.; Luo, H.; Luo, Z.; Zhang, T.; Yang, N.; Long, X.; Xie, H.; Qiu, W.; Zhang, B.; et al. β-elemene acts as an antitumor factor and downregulates the expression of survivin, Bcl-xL and Mta-1. Mol. Med. Report 2012, 6, 989–995. [Google Scholar]
- Zhu, T.; Xu, Y.; Dong, B.; Zhang, J.; Wei, Z.; Xu, Y.; Yao, Y. β-elemene inhibits proliferation of human glioblastoma cells through the activation of glia maturation factor β and induces sensitization to cisplatin. Oncol. Rep 2011, 26, 405–413. [Google Scholar]
- Zhu, T.; Zhao, Y.; Zhang, J.; Li, L.; Zou, L.; Yao, Y.; Xu, Y. ß-Elemene inhibits proliferation of human glioblastoma cells and causes cell-cycle G0/G1 arrest via mutually compensatory activation of MKK3 and MKK6. Int. J. Oncol 2011, 38, 419–426. [Google Scholar]
- Wang, B.; Peng, X.X.; Sun, R.; Li, J.; Zhan, X.R.; Wu, L.J.; Wang, S.L.; Xie, T. Systematic review of β-elemene injection as adjunctive treatment for lung cancer. Chin. J. Integr. Med 2012, 18, 813–823. [Google Scholar]
- Xu, H.B.; Li, L.; Fu, J.; Mao, X.P.; Xu, L.Z. Reversion of multidrug resistance in a chemoresistant human breast cancer cell line by β-elemene. Pharmacology 2012, 89, 303–312. [Google Scholar]
- McCubrey, J.A.; Steelman, L.S.; Abrams, S.L.; Lee, J.T.; Chang, F.; Bertrand, F.E.; Navolanic, P.M.; Terrian, D.M.; Franklin, R.A.; D’Assoro, A.B.; et al. Roles of the RAF/MEK/ERK and PI3K/PTEN/AKT pathways in malignant transformation and drug resistance. Adv. Enzyme Regul 2006, 46, 249–279. [Google Scholar]
- Zhang, Y.; Qu, X.J.; Liu, Y.P.; Yang, X.H.; Hou, K.Z.; Teng, Y.E.; Zhang, J.D. Reversal effect of PI3-K inhibitor LY294002 on P-glycoprotein-mediated multidrug resistance of human leukemia cell line K562/DNR and gastric cancer cell line SGC7901/ADR. Ai Zheng 2009, 28, 97–99. [Google Scholar]
- Barancík, M.; Bohácová, V.; Sedlák, J.; Sulová, Z.; Breier, A. LY294,002, a specific inhibitor of PI3K/Akt kinase pathway, antagonizes P-glycoprotein-mediated multidrug resistance. Eur. J. Pharm. Sci 2006, 29, 426–434. [Google Scholar]
- Steelman, L.S.; Stadelman, K.M.; Chappell, W.H.; Horn, S.; Bäsecke, J.; Cervello, M.; Nicoletti, F.; Libra, M.; Stivala, F.; Martelli, A.M.; et al. Akt as a therapeutic target in cancer. Expert Opin. Ther. Targets 2008, 12, 1139–1165. [Google Scholar]
- Dulinska-Litewka, J.; McCubrey, J.A.; Laidler, P. Increased akt signaling resulting from the loss of androgen responsiveness in prostate cancer. Curr. Med. Chem 2013, 20, 144–157. [Google Scholar]
- Choi, B.H.; Kim, C.G.; Lim, Y.; Shin, S.Y.; Lee, Y.H. Curcumin down-regulates the multidrug-resistance mdr1b gene by inhibiting the PI3K/Akt/NF kappa B pathway. Cancer Lett 2008, 18, 111–118. [Google Scholar]
- Mohapatra, B.; Ahmad, G.; Nadeau, S.; Zutshi, N.; An, W.; Scheffe, S.; Dong, L.; Feng, D.; Goetz, B.; Arya, P.; et al. Protein tyrosine kinase regulation by ubiquitination: Critical roles of Cbl-family ubiquitin ligases. Biochim. Biophys. Acta 2013, 1833, 122–139. [Google Scholar]
- Brennan, T.; Adapala, N.S.; Barbe, M.F.; Yingling, V.; Sanjay, A. Abrogation of Cbl-PI3K interaction increases bone formation and osteoblast proliferation. Calcif. Tissue Int 2011, 89, 396–410. [Google Scholar]
- Adapala, N.S.; Barbe, M.F.; Langdon, W.Y.; Nakamura, M.C.; Tsygankov, A.Y.; Sanjay, A. The loss of Cbl-phosphatidylinositol 3-kinase interaction perturbs RANKL-mediated signaling, inhibiting bone resorption and promoting osteoclast survival. J. Biol. Chem 2010, 285, 36745–36758. [Google Scholar]
- Zhang, Y.; Qu, X.; Hu, X.; Yang, X.; Hou, K.; Teng, Y.; Zhang, J.; Sada, K.; Liu, Y. Reversal of P-glycoprotein-mediated multi-drug resistance by the E3 ubiquitin ligase Cbl-b in human gastric adenocarcinoma cells. J. Pathol 2009, 218, 248–255. [Google Scholar]
- Li, Q.Q.; Lee, R.X.; Liang, H.; Zhong, Y. Anticancer Activity of β-Elemene and its synthetic analogs in human malignant brain tumor cells. Anticancer Res 2013, 33, 65–76. [Google Scholar]
- Li, L.; Xu, L.; Qu, X.; Zhao, M.; Yu, P.; Kang, J.; Liu, Y.; Hu, X. Cbl-regulated Akt and ERK signals are involved in β-elemene-induced cell apoptosis in lung cancer cells. Mol. Med. Report 2011, 4, 1243–1246. [Google Scholar]
- Wang, J.W.; Zhang, H.P.; Sun, Y. Phase IlI clinical trial of elemene emulsion in the management of malignant pleural and peritoneal effusion. Chin. J. Oncol 1996, 18, 464. [Google Scholar]
- Tan, P.; Zhong, W.; Cai, W. Clinical study on treatment of 40 cases of malignant brain tumor by elemene emulsion injection. Zhongguo Zhong Xi Yi Jie He Za Zhi 2000, 20, 645–648. [Google Scholar]
- Li, Q.Q.; Lee, R.X.; Liang, H.; Zhong, Y.; Reed, E. Enhancement of cisplatin-induced apoptosis by β-elemene in resistant human ovarian cancer cells. Med. Oncol 2013, 30, 424. [Google Scholar]
- Zhang, F.; Xu, L.; Qu, X.; Zhao, M.; Jin, B.; Kang, J.; Liu, Y.; Hu, X. Synergistic antitumor effect of β-elemene and etoposide is mediated via induction of cell apoptosis and cell cycle arrest in non-small cell lung carcinoma cells. Mol. Med. Report 2011, 4, 1189–1193. [Google Scholar]
- Lee, J.T., Jr; Steelman, L.S.; McCubrey, J.A. Phosphatidylinositol 3′-kinase activation leads to multidrug resistance protein-1 expression and subsequent chemoresistance in advanced prostate cancer cells. Cancer Res 2004, 64, 8397–8404. [Google Scholar]
- Xu, L.; Zhang, Y.; Liu, J.; Qu, J.; Hu, X.; Zhang, F.; Zheng, H.; Qu, X.; Liu, Y. TRAIL-activated EGFR by Cbl-b-regulated EGFR redistribution in lipid rafts antagonises TRAIL-induced apoptosis in gastric cancer cells. Eur. J. Cancer 2012, 48, 3288–3299. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | IC50 ± SDs μmol/L (fold reversal) | IC50 ± SDs μmol/L (fold reversal) | ||
|---|---|---|---|---|
| K562 | K562/DNR | SGC7901 | SGC7901/ADR | |
| Doxorubicin | 0.096 ± 0.012 (1.00) | 5.014 ± 0.106 (1.00) | 2.032 ± 0.041 (1.00) | 20.011 ± 0.006 (1.00) |
| +1 μg/mL β-elemene | 0.086 ± 0.009 (1.12) | 2.219 ± 0.092 ** (2.26) | 2.026 ± 0.011 (1.01) | 14.276 ± 0.092 ** (1.40) |
| +5 μg/mL β-elemene | 0.088 ± 0.017 (1.09) | 0.519 ± 0.036 ** (9.65) | 2.014 ± 0.016 (1.01) | 4.003 ± 0.107 ** (5.00) |
| +10 μg/mL β-elemene | 0.082 ± 0.022 (1.17) | 0.303 ± 0.024 ** (16.55) | 2.002 ± 0.022 (1.02) | 2.503 ± 0.012 ** (8.01) |
| +10 μM verapamil | 0.081 ± 0.034 (1.18) | 0.231 ± 0.017 ** (21.67) | 2.002 ± 0.024 (1.02) | 2.202 ± 0.106 ** (9.09) |
| Daunorubicin | 0.0091 ± 0.0004 (1.00) | 1.229 ± 0.021 (1.00) | 0.288 ± 0.011 (1.00) | 2.269 ± 0.013 (1.00) |
| +1 μg/mL β-elemene | 0.0088 ± 0.0005 (1.03) | 1.538 ± 0.009 ** (3.26) | 0.279 ± 0.007 (1.03) | 1.881 ± 0.007 ** (1.21) |
| +5 μg/mL β-elemene | 0.0090 ± 0.0003 (1.01) | 0.707 ± 0.012 ** (7.09) | 0.280 ± 0.013 (1.03) | 0.764 ± 0.090 ** (2.97) |
| +10 μg/mL β-elemene | 0.0090 ± 0.0002 (1.01) | 0.344 ± 0.032 ** (14.57) | 0.281 ± 0.022 (1.02) | 0.206 ± 0.022 ** (11.02) |
| +10 μM verapamil | 0.0087 ± 0.0001 (1.04) | 0.238 ± 0.025 ** (20.09) | 0.277 ± 0.014 (1.04) | 0.129 ± 0.025 ** (17.59) |
| Epirubicin | 0.019 ± 0.005 (1.00) | 0.963 ± 0.009 (1.00) | 2.114 ± 0.013 (1.00) | 18.075 ± 0.061 (1.00) |
| +1 μg/mL β-elemene | 0.018 ± 0.003 (1.04) | 0.231 ± 0.004 ** (4.16) | 2.069 ± 0.009 (1.02) | 11.296 ± 0.004 ** (1.60) |
| +5 μg/mL β-elemene | 0.019 ± 0.003 (1.02) | 0.133 ± 0.007 ** (7.24) | 2.046 ± 0.010 (1.03) | 5.124 ± 0.013 ** (3.53) |
| +10 μg/mL β-elemene | 0.017 ± 0.012 (1.07) | 0.064 ± 0.002 ** (15.13) | 2.043 ± 0.032 (1.03) | 1.663 ± 0.067 ** (10.87) |
| +10 μM verapamil | 0.018 ± 0.003 (1.05) | 0.043 ± 0.005 ** (22.53) | 2.046 ± 0.019 (1.03) | 0.993 ± 0.025 ** (18.20) |
| Cisplatin | 3.174 ± 0.012 (1.00) | 2.861 ± 0.036 (1.00) | 3.991 ± 0.040 (1.00) | 4.203 ± 0.109 (1.00) |
| +10 μg/mL β-elemene | 2.784 ± 0.097 (1.14) | 2.983 ± 0.103 (0.96) | 3.905 ± 0.011 (1.02) | 4.119 ± 0.078 (1.02) |
| +10 μM verapamil | 2.994 ± 0.021 (1.06) | 2.778 ± 0.015 (1.03) | 3.823 ± 0.013 (1.04) | 4.108 ± 0.062 (1.02) |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, Y.; Mu, X.-D.; Li, E.-Z.; Luo, Y.; Song, N.; Qu, X.-J.; Hu, X.-J.; Liu, Y.-P. The Role of E3 Ubiquitin Ligase Cbl Proteins in β-Elemene Reversing Multi-Drug Resistance of Human Gastric Adenocarcinoma Cells. Int. J. Mol. Sci. 2013, 14, 10075-10089. https://doi.org/10.3390/ijms140510075
Zhang Y, Mu X-D, Li E-Z, Luo Y, Song N, Qu X-J, Hu X-J, Liu Y-P. The Role of E3 Ubiquitin Ligase Cbl Proteins in β-Elemene Reversing Multi-Drug Resistance of Human Gastric Adenocarcinoma Cells. International Journal of Molecular Sciences. 2013; 14(5):10075-10089. https://doi.org/10.3390/ijms140510075
Chicago/Turabian StyleZhang, Ye, Xiao-Dong Mu, En-Zhe Li, Ying Luo, Na Song, Xiu-Juan Qu, Xue-Jun Hu, and Yun-Peng Liu. 2013. "The Role of E3 Ubiquitin Ligase Cbl Proteins in β-Elemene Reversing Multi-Drug Resistance of Human Gastric Adenocarcinoma Cells" International Journal of Molecular Sciences 14, no. 5: 10075-10089. https://doi.org/10.3390/ijms140510075