Identification of Plasma Metabolomic Profiling for Diagnosis of Esophageal Squamous-Cell Carcinoma Using an UPLC/TOF/MS Platform

Abstract
:1. Introduction
2. Results
2.1. Demographic Characteristics
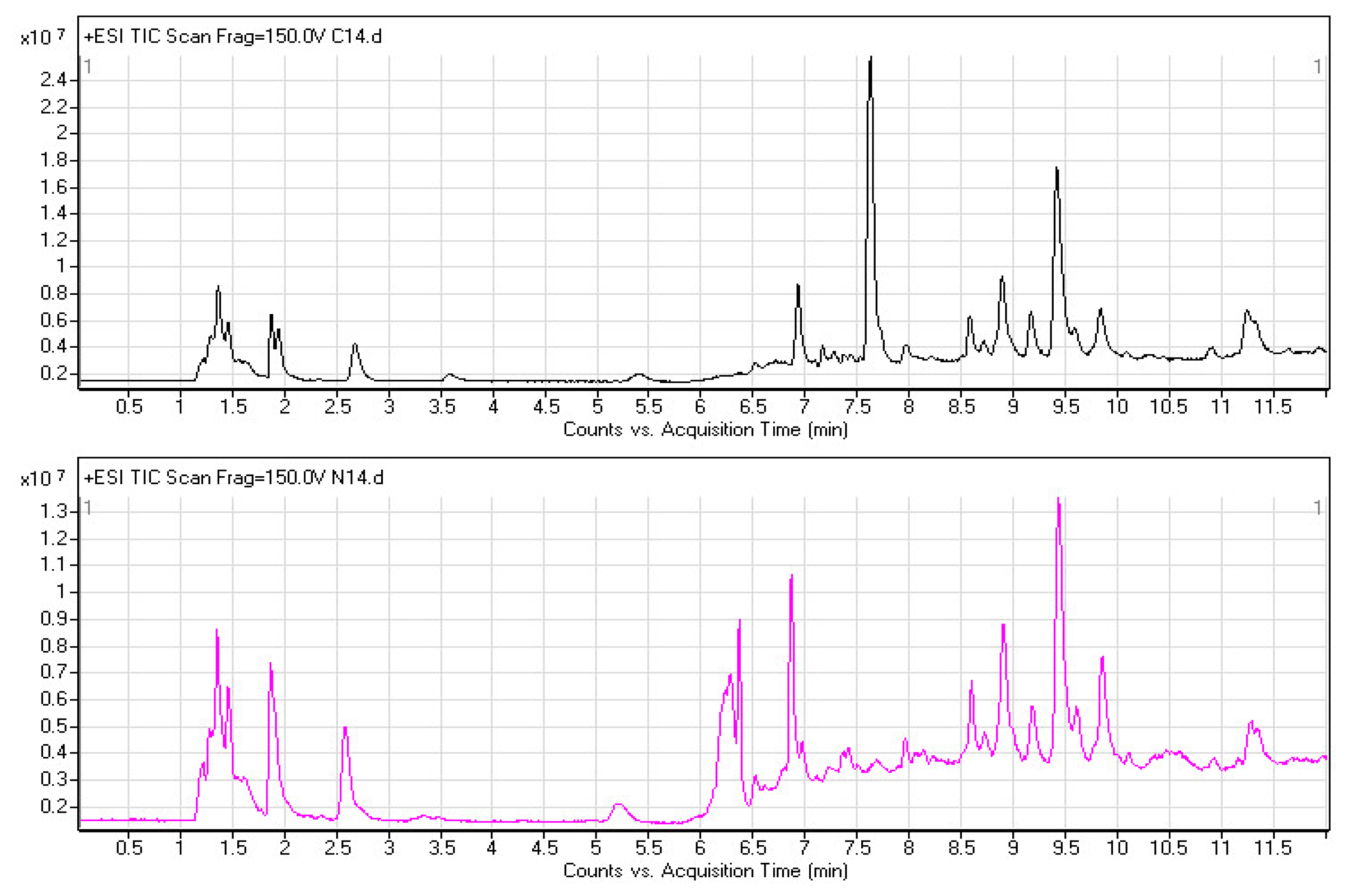
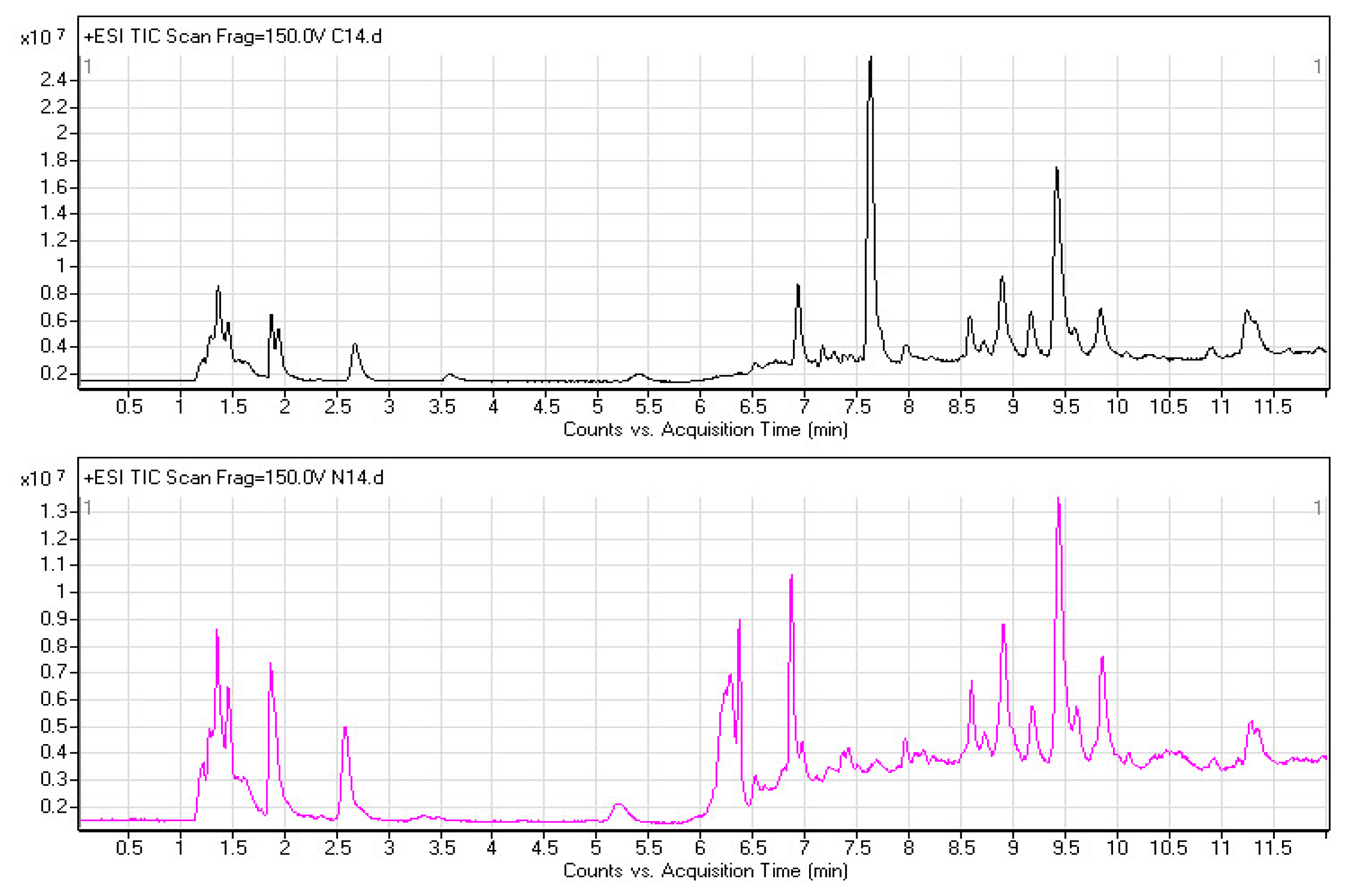
2.2. Plasma Metabolomic Profiles
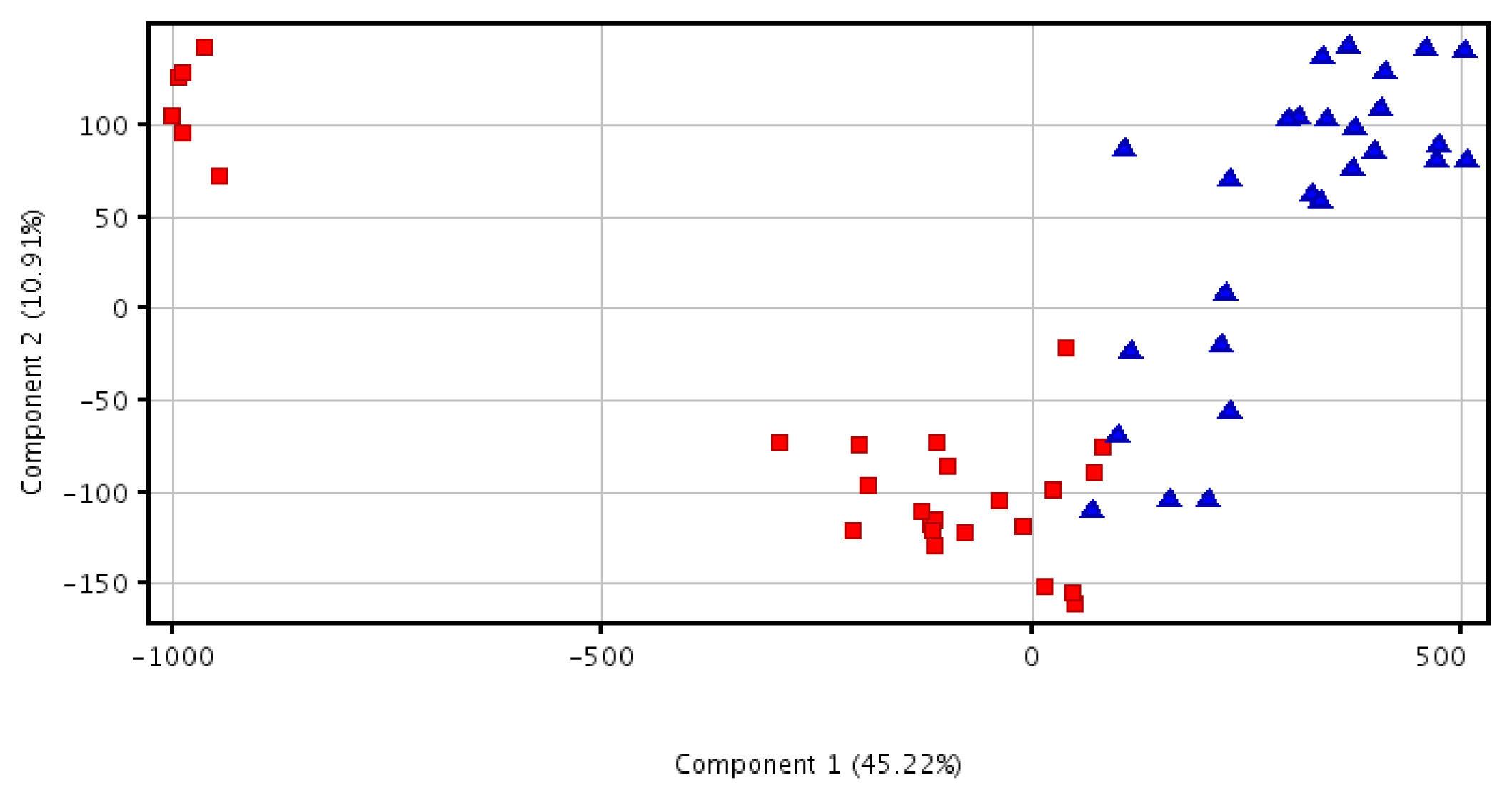
2.3. Principal Component Analysis Analysis
2.4. Discovery and Identification of Metabolic Biomarkers
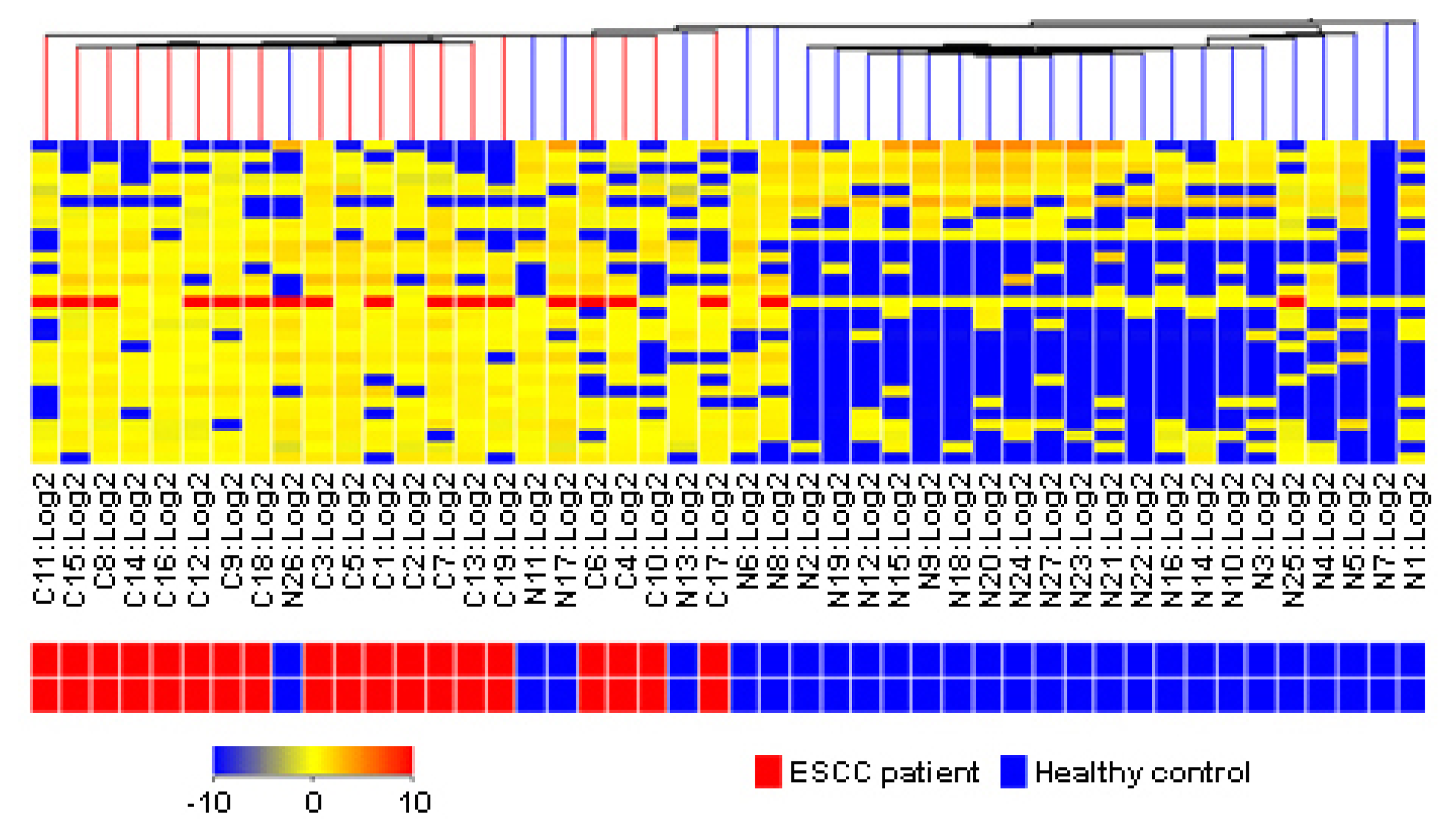
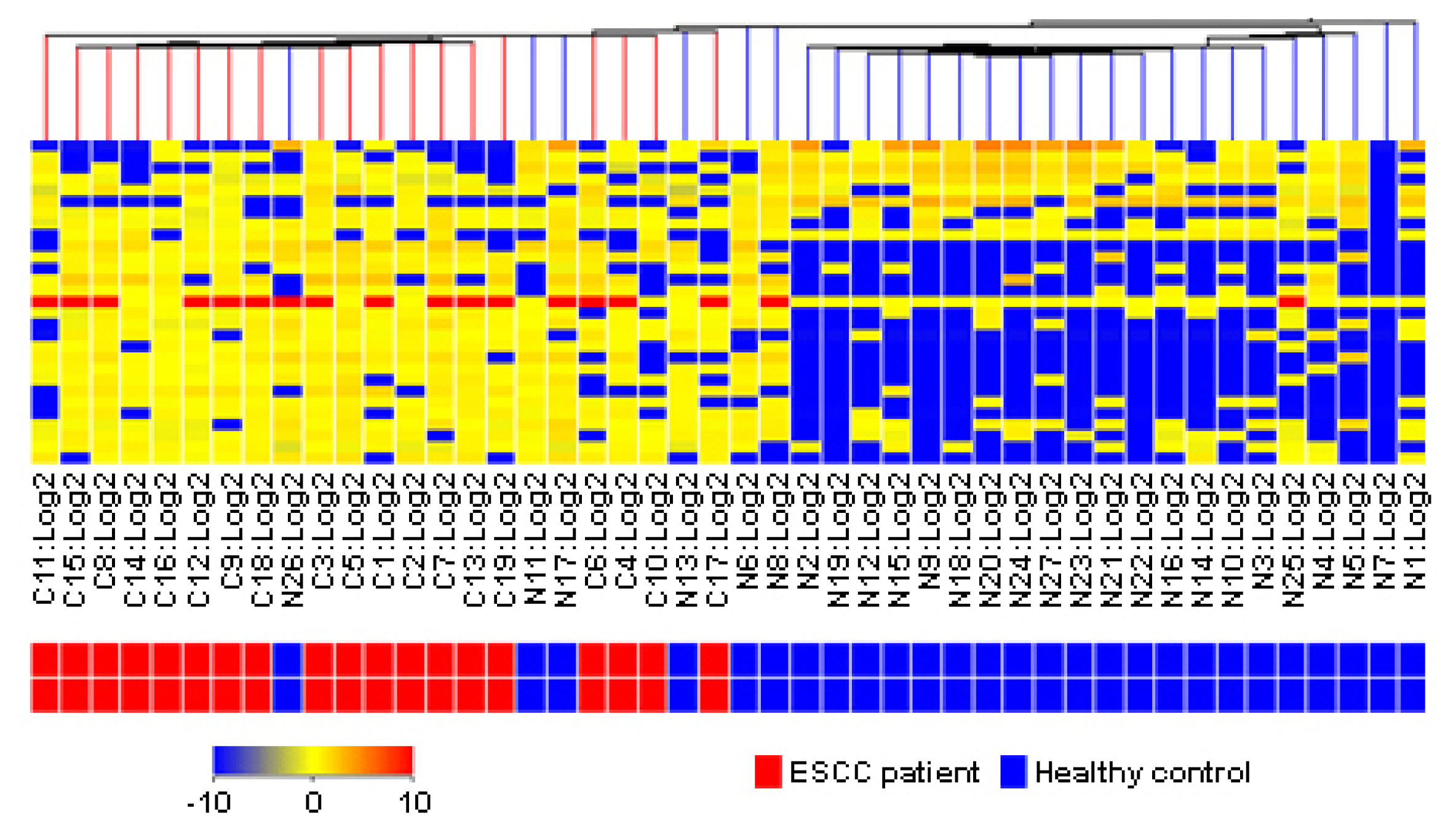
2.5. Clinical Estimation of Metabolic Biomarkers with Hierarchical Cluster Analysis
3. Material and Methods
3.1. Study Subjects
3.2. Sample Preparation and Pretreatment
3.3. Ultraperformance Liquid Chromatography
3.4. Accurate Mass Time-of-Flight Mass Spectrometry
3.5. Data Preprocessing and Annotation
3.6. Statistics
3.7. Metabolite Identification
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Enzinger, P.C.; Mayer, R.J. Esophageal cancer. N. Engl. J. Med 2003, 349, 2241–2252. [Google Scholar]
- Zhou, J.; Xiang, Q.; Tao, R.; Yang, J.; Lin, P.; Wu, M. Analysis on surveillance of cause death in Jiangsu province during 2007. Jiangsu J. Prev. Med 2008, 19, 74–75. [Google Scholar]
- Rice, T.W.; Rusch, V.W.; Apperson-Hansen, C.; Allen, M.S.; Chen, L.Q.; Hunter, J.G.; Kesler, K.A.; Law, S.; Lerut, T.E.; Reed, C.E.; et al. Worldwide esophageal cancer collaboration. Dis. Esophagus 2009, 22, 1–8. [Google Scholar]
- Nicholson, J.K.; Lindon, J.C. Systems biology: Metabonomics. Nature 2008, 455, 1054–1056. [Google Scholar]
- Idle, J.R.; Gonzalez, F.J. Metabolomics. Cell Metab 2007, 6, 348–351. [Google Scholar]
- Syggelou, A.; Iacovidou, N.; Atzori, L.; Xanthos, T.; Fanos, V. Metabolomics in the developing human being. Pediatr. Clin. N. Am 2012, 59, 1039–1058. [Google Scholar]
- Johnson, C.H.; Gonzalez, F.J. Challenges and opportunities of metabolomics. J. Cell. Physiol 2012, 227, 2975–2981. [Google Scholar]
- Ellis, D.I.; Dunn, W.B.; Griffin, J.L.; Allwood, J.W.; Goodacre, R. Metabolic fingerprinting as a diagnostic tool. Pharmacogenomics 2007, 8, 1243–1266. [Google Scholar]
- Ma, Y.; Zhang, P.; Yang, Y.; Wang, F.; Qin, H. Metabolomics in the fields of oncology: A review of recent research. Mol. Biol. Rep 2012, 39, 7505–7511. [Google Scholar]
- O’Connell, T.M. Recent advances in metabolomics in oncology. Bioanalysis 2012, 4, 431–451. [Google Scholar]
- Mamas, M.; Dunn, W.B.; Neyses, L.; Goodacre, R. The role of metabolites and metabolomics in clinically applicable biomarkers of disease. Arch. Toxicol 2011, 85, 5–17. [Google Scholar]
- Claudino, W.M.; Quattrone, A.; Biganzoli, L.; Pestrin, M.; Bertini, I.; Di Leo, A. Metabolomics: Available results, current research projects in breast cancer, and future applications. J. Clin. Oncol 2007, 25, 2840–2846. [Google Scholar]
- Qiu, Y.; Cai, G.; Su, M.; Chen, T.; Zheng, X.; Xu, Y.; Ni, Y.; Zhao, A.; Xu, L.X.; Cai, S.; et al. Serum metabolite profiling of human colorectal cancer using gc-tofms and uplc-qtofms. J. Proteome. Res 2009, 8, 4844–4850. [Google Scholar]
- Zhou, J.; Xu, B.; Huang, J.; Jia, X.; Xue, J.; Shi, X.; Xiao, L.; Li, W. 1h nmr-based metabonomic and pattern recognition analysis for detection of oral squamous cell carcinoma. Clin. Chim. Acta 2009, 401, 8–13. [Google Scholar]
- Sreekumar, A.; Poisson, L.M.; Rajendiran, T.M.; Khan, A.P.; Cao, Q.; Yu, J.; Laxman, B.; Mehra, R.; Lonigro, R.J.; Li, Y.; et al. Metabolomic profiles delineate potential role for sarcosine in prostate cancer progression. Nature 2009, 457, 910–914. [Google Scholar]
- Zhang, A.; Sun, H.; Wang, X. Serum metabolomics as a novel diagnostic approach for disease: A systematic review. Anal. Bioanal. Chem 2012, 404, 1239–1245. [Google Scholar]
- Van Meer, G.; de Kroon, A.I. Lipid map of the mammalian cell. J. Cell Sci 2011, 124, 5–8. [Google Scholar]
- Wymann, M.P.; Schneiter, R. Lipid signalling in disease. Nat. Rev. Mol. Cell Biol 2008, 9, 162–176. [Google Scholar]
- Dueck, D.A.; Chan, M.; Tran, K.; Wong, J.T.; Jay, F.T.; Littman, C.; Stimpson, R.; Choy, P.C. The modulation of choline phosphoglyceride metabolism in human colon cancer. Mol. Cell. Biochem 1996, 162, 97–103. [Google Scholar]
- Monteggia, E.; Colombo, I.; Guerra, A.; Berra, B. Phospholipid distribution in murine mammary adenocarcinomas induced by activated neu oncogene. Cancer Detect. Prev 2000, 24, 207–211. [Google Scholar]
- Dobrzynska, I.; Szachowicz-Petelska, B.; Sulkowski, S.; Figaszewski, Z. Changes in electric charge and phospholipids composition in human colorectal cancer cells. Mol. Cell. Biochem 2005, 276, 113–119. [Google Scholar]
- Ruiz-Cabello, J.; Cohen, J.S. Phospholipid metabolites as indicators of cancer cell function. NMR Biomed 1992, 5, 226–233. [Google Scholar]
- Fernandis, A.Z.; Wenk, M.R. Lipid-based biomarkers for cancer. J. Chromatogr. B 2009, 877, 2830–2835. [Google Scholar]
- Patra, S.K. Dissecting lipid raft facilitated cell signaling pathways in cancer. Biochim. Biophys. Acta 2008, 1785, 182–206. [Google Scholar]
- Di Paolo, G.; de Camilli, P. Phosphoinositides in cell regulation and membrane dynamics. Nature 2006, 443, 651–657. [Google Scholar]
- Doria, M.L.; Cotrim, Z.; Macedo, B.; Simoes, C.; Domingues, P.; Helguero, L.; Domingues, M.R. Lipidomic approach to identify patterns in phospholipid profiles and define class differences in mammary epithelial and breast cancer cells. Breast Cancer Res. Treat 2012, 133, 635–648. [Google Scholar]
- Ackerstaff, E.; Glunde, K.; Bhujwalla, Z.M. Choline phospholipid metabolism: A target in cancer cells? J. Cell. Biochem 2003, 90, 525–533. [Google Scholar]
- Jackowski, S. Cell cycle regulation of membrane phospholipid metabolism. J. Biol. Chem 1996, 271, 20219–20222. [Google Scholar]
- Jackowski, S. Coordination of membrane phospholipid synthesis with the cell cycle. J. Biol. Chem 1994, 269, 3858–3867. [Google Scholar]
- Podo, F. Tumour phospholipid metabolism. NMR Biomed 1999, 12, 413–439. [Google Scholar]
- Li, S.Y.; Yu, B.; An, P.; Liang, Z.J.; Yuan, S.J.; Cai, H.Y. Effects of cell membrane phospholipid level and protein kinase c isoenzyme expression on hepatic metastasis of colorectal carcinoma. Hepatobiliary Pancreat. Dis. Int 2004, 3, 411–416. [Google Scholar]
- Stace, C.L.; Ktistakis, N.T. Phosphatidic acid- and phosphatidylserine-binding proteins. Biochim. Biophys. Acta 2006, 1761, 913–926. [Google Scholar]
- Yeung, T.; Gilbert, G.E.; Shi, J.; Silvius, J.; Kapus, A.; Grinstein, S. Membrane phosphatidylserine regulates surface charge and protein localization. Science 2008, 319, 210–213. [Google Scholar]
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: An enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol 2003, 4, 397–407. [Google Scholar]
- Colie, S.; van Veldhoven, P.P.; Kedjouar, B.; Bedia, C.; Albinet, V.; Sorli, S.C.; Garcia, V.; Djavaheri-Mergny, M.; Bauvy, C.; Codogno, P.; et al. Disruption of sphingosine 1-phosphate lyase confers resistance to chemotherapy and promotes oncogenesis through bcl-2/bcl-xl upregulation. Cancer Res 2009, 69, 9346–9353. [Google Scholar]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol 2008, 9, 139–150. [Google Scholar]
- Segui, B.; Andrieu-Abadie, N.; Jaffrezou, J.P.; Benoist, H.; Levade, T. Sphingolipids as modulators of cancer cell death: Potential therapeutic targets. Biochim. Biophys. Acta 2006, 1758, 2104–2120. [Google Scholar]
- Ponnusamy, S.; Meyers-Needham, M.; Senkal, C.E.; Saddoughi, S.A.; Sentelle, D.; Selvam, S.P.; Salas, A.; Ogretmen, B. Sphingolipids and cancer: Ceramide and sphingosine-1-phosphate in the regulation of cell death and drug resistance. Future Oncol 2010, 6, 1603–1624. [Google Scholar]
- Wang, D.; Zhao, Z.; Caperell-Grant, A.; Yang, G.; Mok, S.C.; Liu, J.; Bigsby, R.M.; Xu, Y. S1p differentially regulates migration of human ovarian cancer and human ovarian surface epithelial cells. Mol. Cancer Ther 2008, 7, 1993–2002. [Google Scholar]
- Kind, T.; Fiehn, O. Advances in structure elucidation of small molecules using mass spectrometry. Bioanal. Rev 2010, 2, 23–60. [Google Scholar]
- Wishart, D.S.; Knox, C.; Guo, A.C.; Eisner, R.; Young, N.; Gautam, B.; Hau, D.D.; Psychogios, N.; Dong, E.; Bouatra, S.; et al. Hmdb: A knowledgebase for the human metabolome. Nucleic Acids Res 2009, 37, D603–D610. [Google Scholar]
- Smith, C.A.; O’Maille, G.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. Metlin: A metabolite mass spectral database. Ther. Drug Monit 2005, 27, 747–751. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Variable | No. of patients (n = 53) | No. of controls (n = 53) |
|---|---|---|
| Age (in years) | 62.6 ± 7.3 | 63.2 ± 8.1 |
| Gender | ||
| Male | 31 | 31 |
| Female | 22 | 22 |
| Smoking index a | ||
| Non-user | 27 * | 40 |
| Ever b | ||
| <400 | 6 | 2 |
| ≥400 | 20 | 11 |
| Alcohol use | ||
| Never | 24 * | 43 |
| Ever c | 29 | 10 |
| Family history | ||
| No | 47 * | 53 |
| Yes | 6 | 0 |
| No. | tR/min | m/z | Tendency (cancer/control) | Fold change | p value | FDR | Postulated identity |
|---|---|---|---|---|---|---|---|
| 1 | 7.0917454 | 837.5655 | up | 1182.88 | 1.43 × 10−8 | 2.86 × 10−6 | phosphatidylinositol |
| 2 | 7.077058 | 881.5899 | up | 1046.25 | 9.10 × 10−9 | 2.73 × 10−6 | |
| 3 | 7.9524574 | 775.527 | up | 887.53 | 7.62 × 10−10 | 4.57 × 10−7 | |
| 4 | 8.084705 | 687.471 | up | 716.17 | 3.98 × 10−7 | 5.97 × 10−5 | |
| 5 | 7.06508 | 925.6148 | up | 667.50 | 4.42 × 10−7 | 5.30 × 10−5 | |
| 6 | 7.6628757 | 777.5422 | up | 664.51 | 2.72 × 10−6 | 2.33 × 10−4 | |
| 7 | 6.780679 | 484.3174 | up | 318.85 | 2.45 × 10−5 | 1.63 × 10−3 | Lithocholyltaurine |
| 8 | 7.091731 | 842.5196 | up | 315.36 | 1.03 × 10−4 | 5.62 × 10−3 | |
| 9 | 7.4925284 | 953.65 | up | 304.278 | 1.14 × 10−5 | 8.55 × 10−4 | |
| 10 | 6.6252184 | 487.3345 | up | 279.64 | 1.48 × 10−6 | 1.48 × 10−4 | phosphatidic acid |
| 11 | 7.2552686 | 939.634 | up | 154.73 | 1.55 × 10−4 | 7.15 × 10−3 | |
| 12 | 6.7809725 | 638.3852 | up | 146.04 | 9.14 × 10−5 | 5.48 × 10−3 | L-Urobilinogen |
| 13 | 7.108005 | 798.4944 | up | 94.10 | 2.87 × 10−4 | 1.15 × 10−2 | |
| 14 | 7.4301453 | 520.8508 | up | 92.60 | 3.04 × 10−4 | 1.14 × 10−2 | |
| 15 | 6.925951 | 784.4782 | up | 73.87 | 3.03 × 10−3 | 5.86 × 10−2 | 9′-carboxy-gama-tocotrienol |
| 16 | 7.108472 | 793.5397 | up | 72.40 | 3.22 × 10−4 | 1.14 × 10−2 | phosphatidyl choline |
| 17 | 6.9349127 | 735.4974 | up | 65.69 | 1.88 × 10−3 | 3.88 × 10−2 | |
| 18 | 7.4347086 | 675.474 | up | 56.34 | 1.44 × 10−3 | 3.61 × 10−2 | phosphatidyl ethanolamine |
| 19 | 7.4338794 | 395.8942 | up | 38.66 | 7.39 × 10−4 | 2.33 × 10−2 | |
| 20 | 7.3625846 | 763.5281 | up | 27.46 | 1.72 × 10−3 | 3.69 × 10−2 | sphinganine 1-phosphate |
| 21 | 7.124603 | 749.513 | up | 26.86 | 1.40 × 10−3 | 3.66 × 10−2 | phosphatidylserine (16:0/14:0) |
| 22 | 7.142187 | 705.487 | up | 25.72 | 9.67 × 10−3 | 1.49 × 10−1 | |
| 23 | 7.1617465 | 661.4607 | up | 24.96 | 2.90 × 10−3 | 5.80 × 10−2 | |
| 24 | 6.9430337 | 696.4259 | up | 22.81 | 8.45 × 10−3 | 1.33 × 10−1 | |
| 25 | 7.1857386 | 617.434 | up | 22.67 | 8.05 × 10−3 | 1.31 × 10−1 | LysoPC(22:2(13Z,16Z)) |
| 26 | 7.43357 | 166.9863 | up | 21.01 | 5.30 × 10−3 | 9.09 × 10−2 | |
| 27 | 6.331661 | 792.7367 | up | 19.60 | 1.54 × 10−3 | 3.69 × 10−2 | |
| 28 | 7.1242256 | 754.4663 | up | 18.79 | 4.46 × 10−3 | 7.86 × 10−2 | Ganglioside GM2 (d18:1/24:1(15Z)) |
| 29 | 6.782452 | 594.3591 | up | 18.78 | 1.19 × 10−3 | 3.39 × 10−2 | Lithocholate 3-O-glucuronide |
| 30 | 7.144019 | 710.4422 | up | 18.49 | 1.78 × 10−3 | 4.10 × 10−2 | |
| 31 | 7.395843 | 719.5016 | up | 18.41 | 3.16 × 10−3 | 5.92 × 10−2 | |
| 32 | 6.6147885 | 443.3086 | up | 17.67 | 1.07 × 10−3 | 3.22 × 10−2 | |
| 33 | 6.6157866 | 448.2641 | up | 12.53 | 5.60 × 10−3 | 9.34 × 10−2 | 12-oxo-20-dihydroxyleukotriene B4 |
| 34 | 6.485909 | 434.249 | up | 8.25 | 3.93 × 10−3 | 7.14 × 10−2 | |
| 35 | 9.1121235 | 527.3036 | down | −13.11 | 1.19 × 10−3 | 3.25 × 10−2 | Desmosine/Isodesmosine |
| 36 | 9.865172 | 628.3065 | down | −25.38 | 2.19 × 10−3 | 4.86 × 10−2 | |
| 37 | 8.604863 | 445.2468 | down | −78.77 | 2.01 × 10−4 | 8.61 × 10−3 | |
| 38 | 7.0976486 | 533.3094 | down | −107.32 | 4.36 × 10−4 | 1.45 × 10−2 | 5-β-cyprinol sulfate |
| 39 | 8.611813 | 586.2785 | down | −233.30 | 1.11 × 10−4 | 5.55 × 10−3 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, R.; Peng, Y.; Li, X.; Wang, Y.; Pan, E.; Guo, W.; Pu, Y.; Yin, L. Identification of Plasma Metabolomic Profiling for Diagnosis of Esophageal Squamous-Cell Carcinoma Using an UPLC/TOF/MS Platform. Int. J. Mol. Sci. 2013, 14, 8899-8911. https://doi.org/10.3390/ijms14058899
Liu R, Peng Y, Li X, Wang Y, Pan E, Guo W, Pu Y, Yin L. Identification of Plasma Metabolomic Profiling for Diagnosis of Esophageal Squamous-Cell Carcinoma Using an UPLC/TOF/MS Platform. International Journal of Molecular Sciences. 2013; 14(5):8899-8911. https://doi.org/10.3390/ijms14058899
Chicago/Turabian StyleLiu, Ran, Yuan Peng, Xiaobo Li, Yi Wang, Enchun Pan, Wei Guo, Yuepu Pu, and Lihong Yin. 2013. "Identification of Plasma Metabolomic Profiling for Diagnosis of Esophageal Squamous-Cell Carcinoma Using an UPLC/TOF/MS Platform" International Journal of Molecular Sciences 14, no. 5: 8899-8911. https://doi.org/10.3390/ijms14058899