The Influence of Polyunsaturated Fatty Acids on the Phospholipase D Isoforms Trafficking and Activity in Mast Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Distribution of PLD Isoforms in C2 Mast Cells with Regard to Stimulation Status
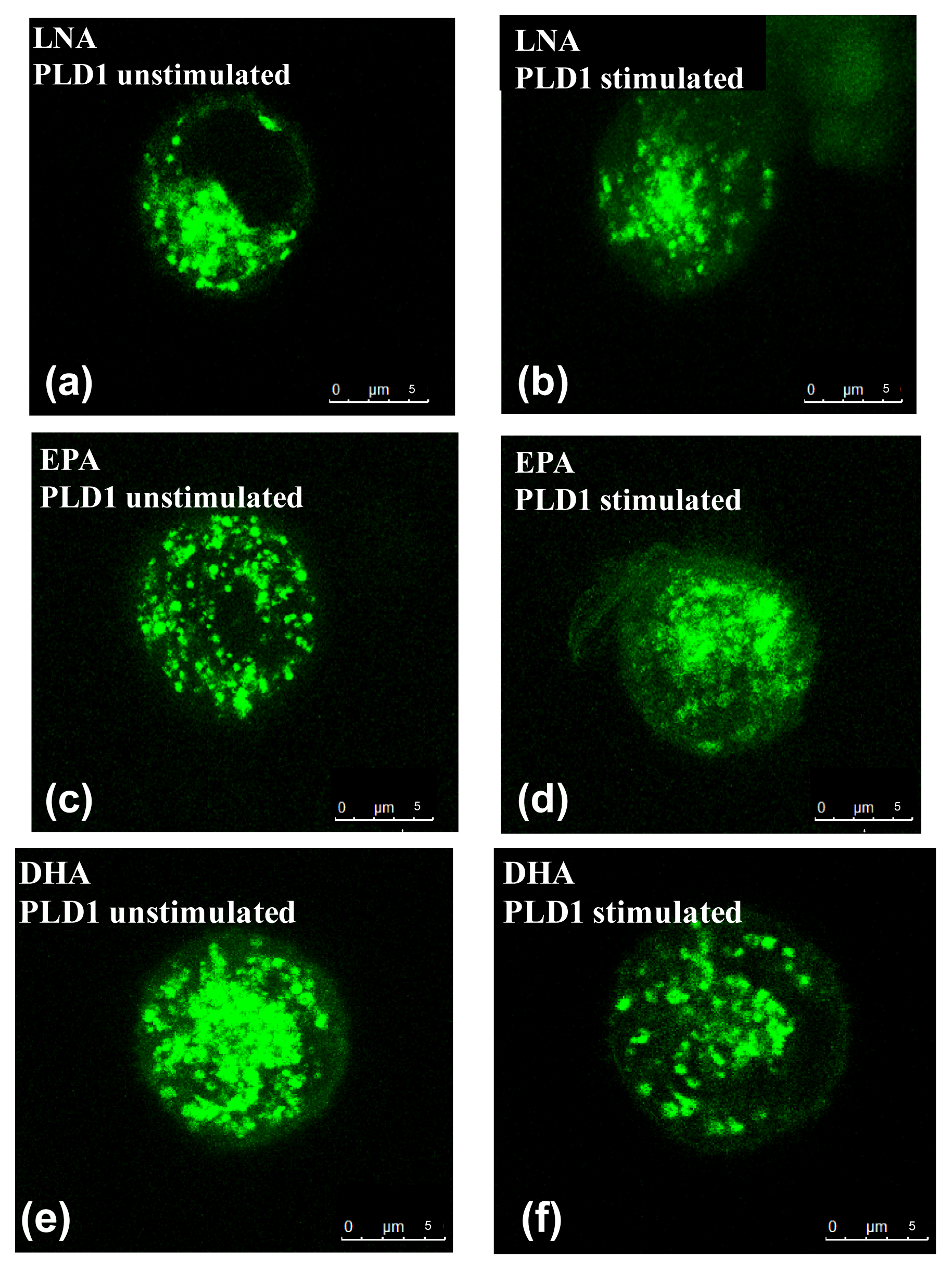
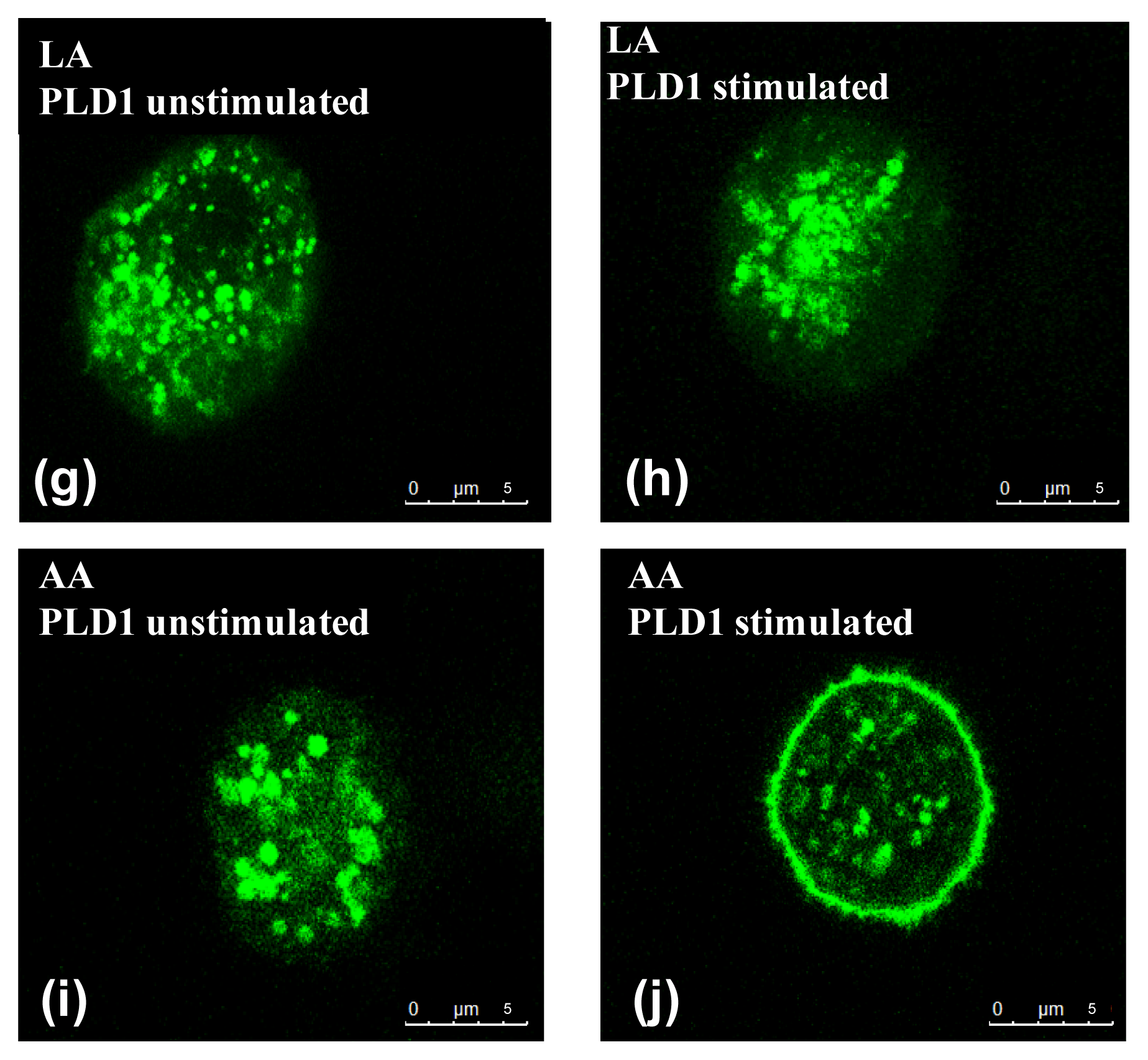
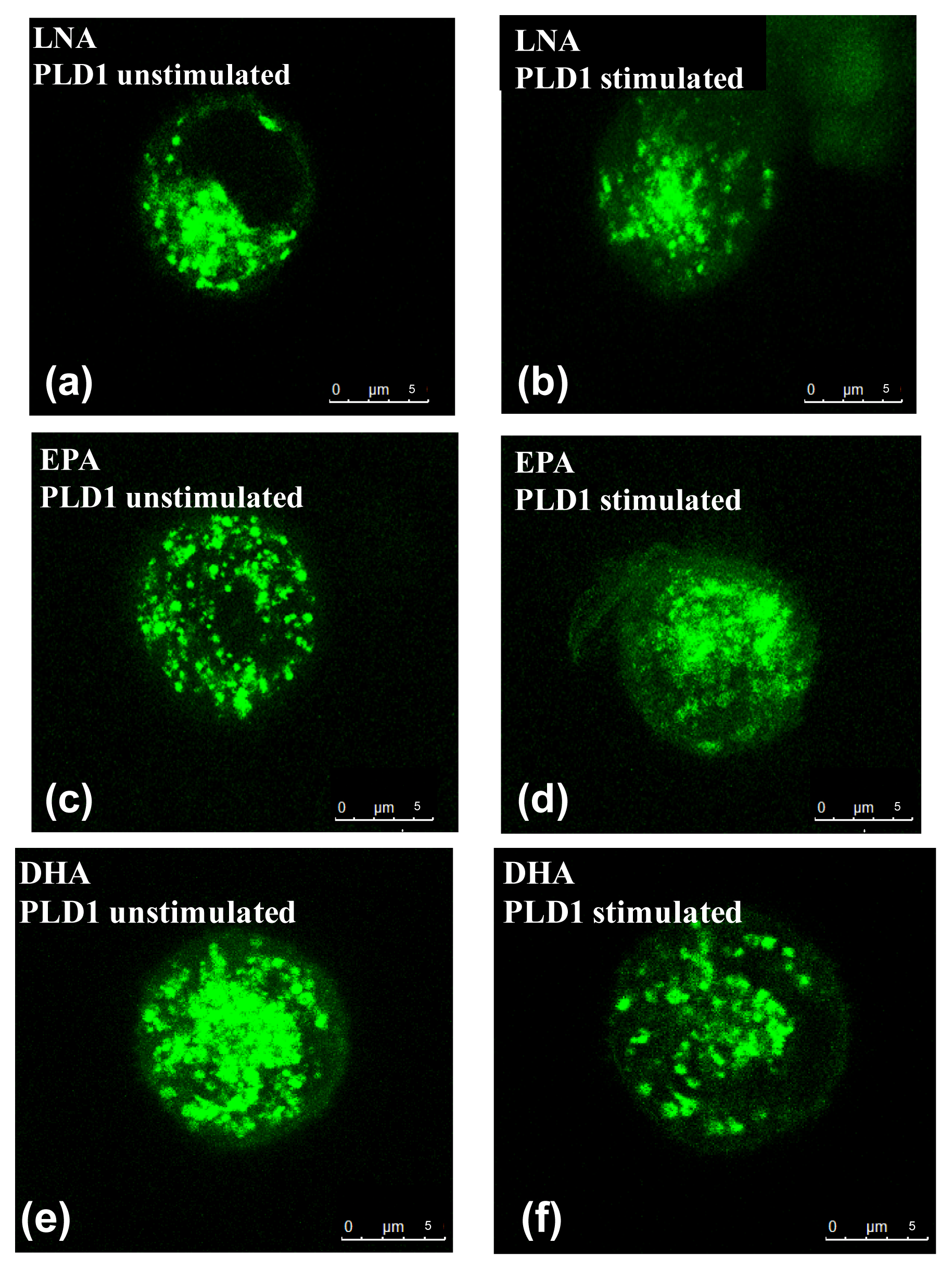
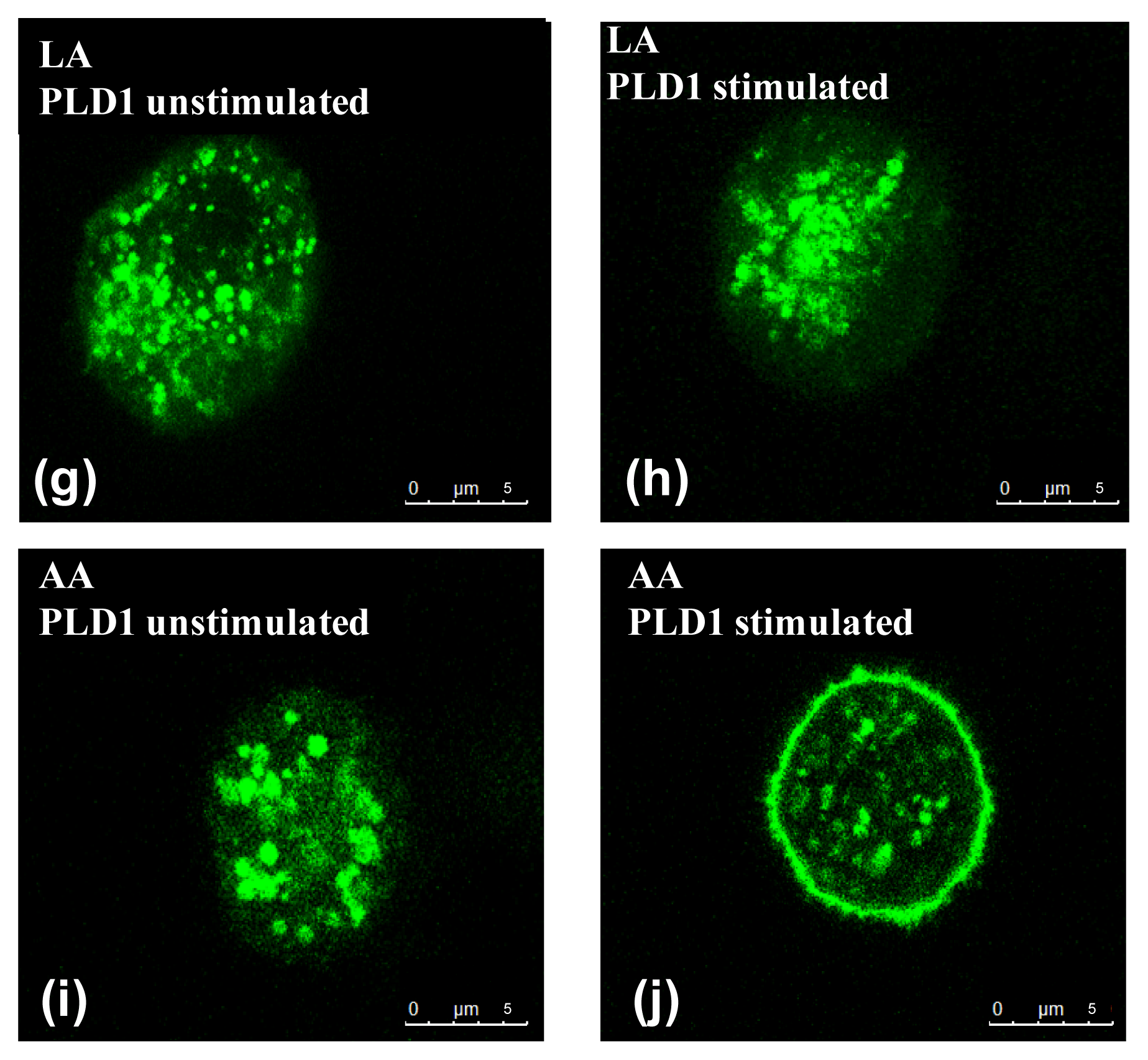
2.2. Influence of PUFA on Distribution of PLD Isoforms with Regard to Stimulation Status
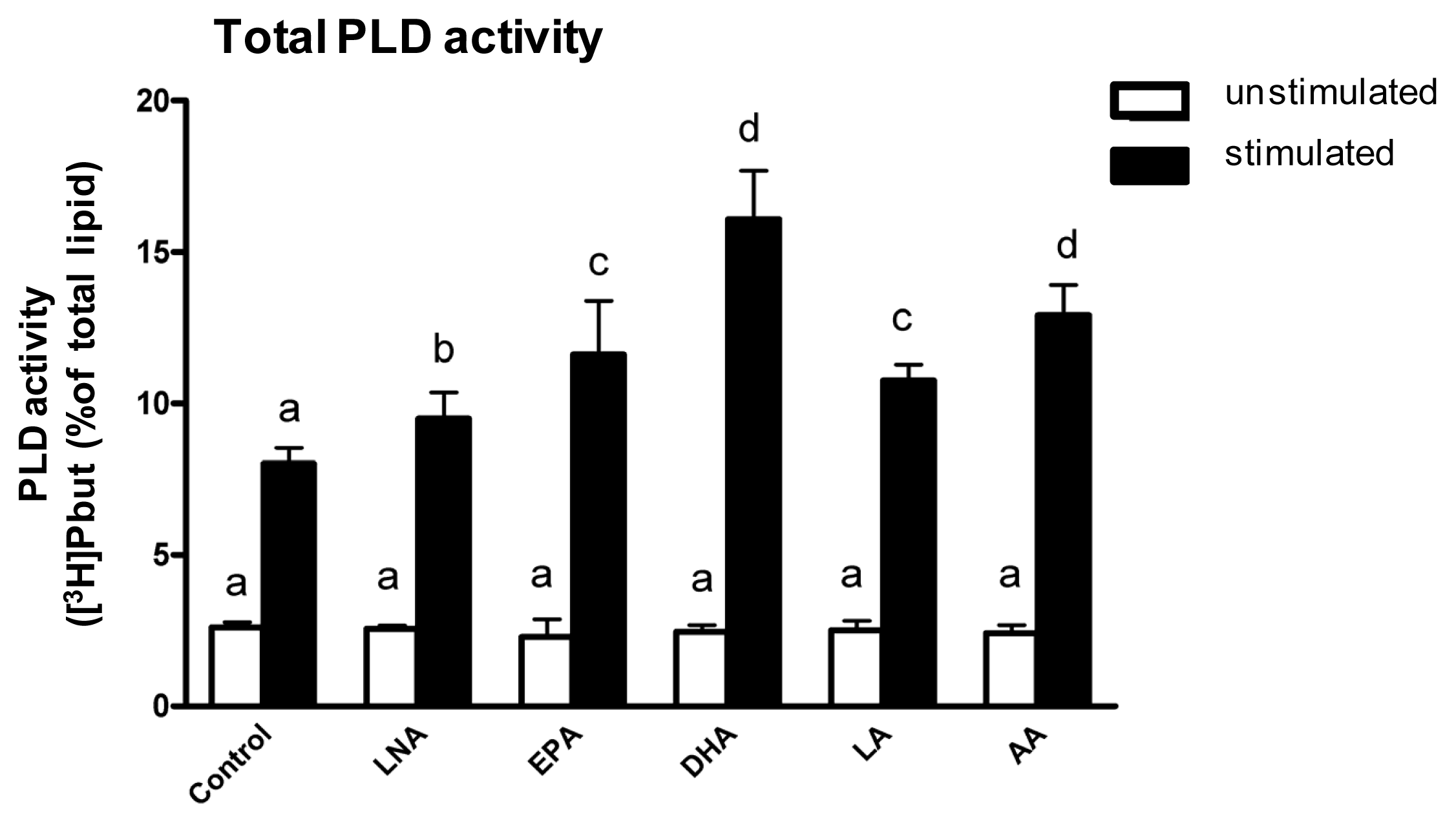
2.3. Influence of PUFA Supplementation on Total PLD Activity
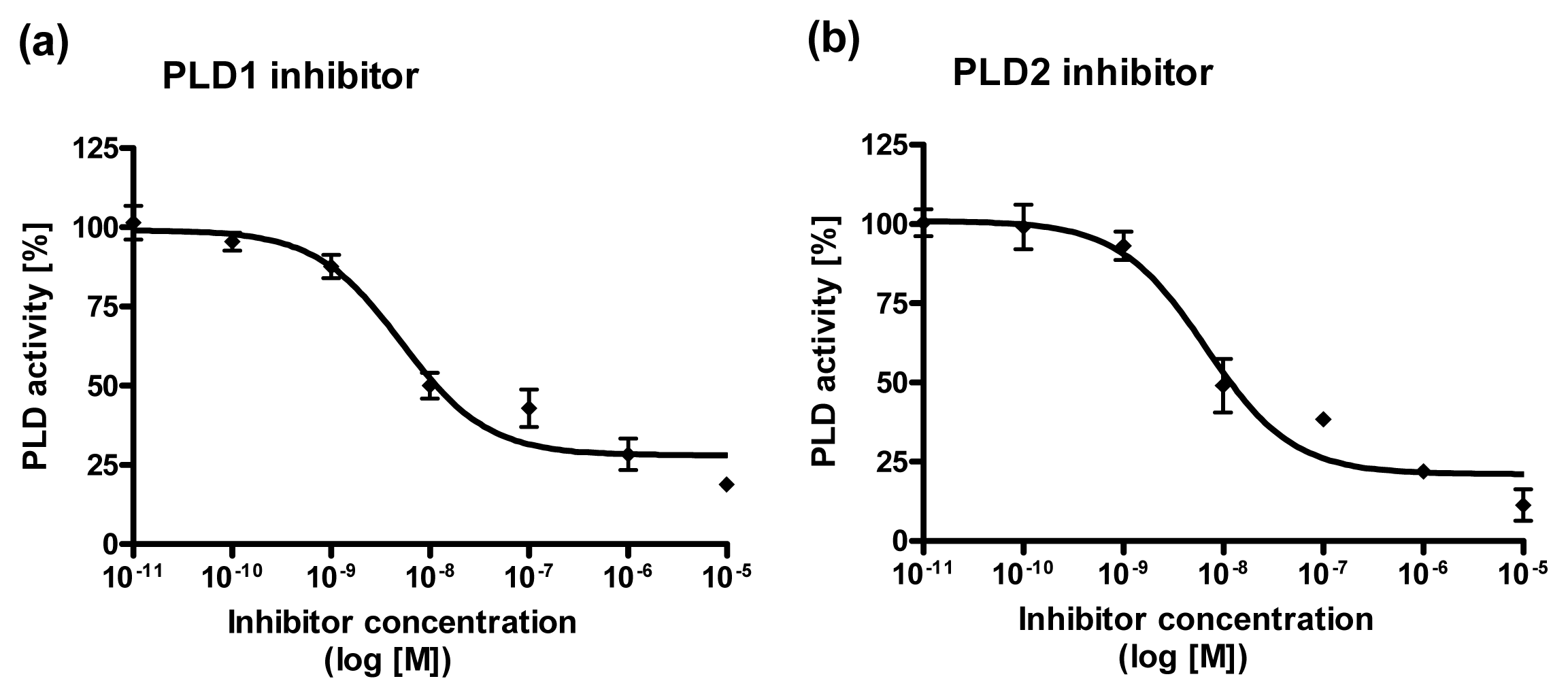
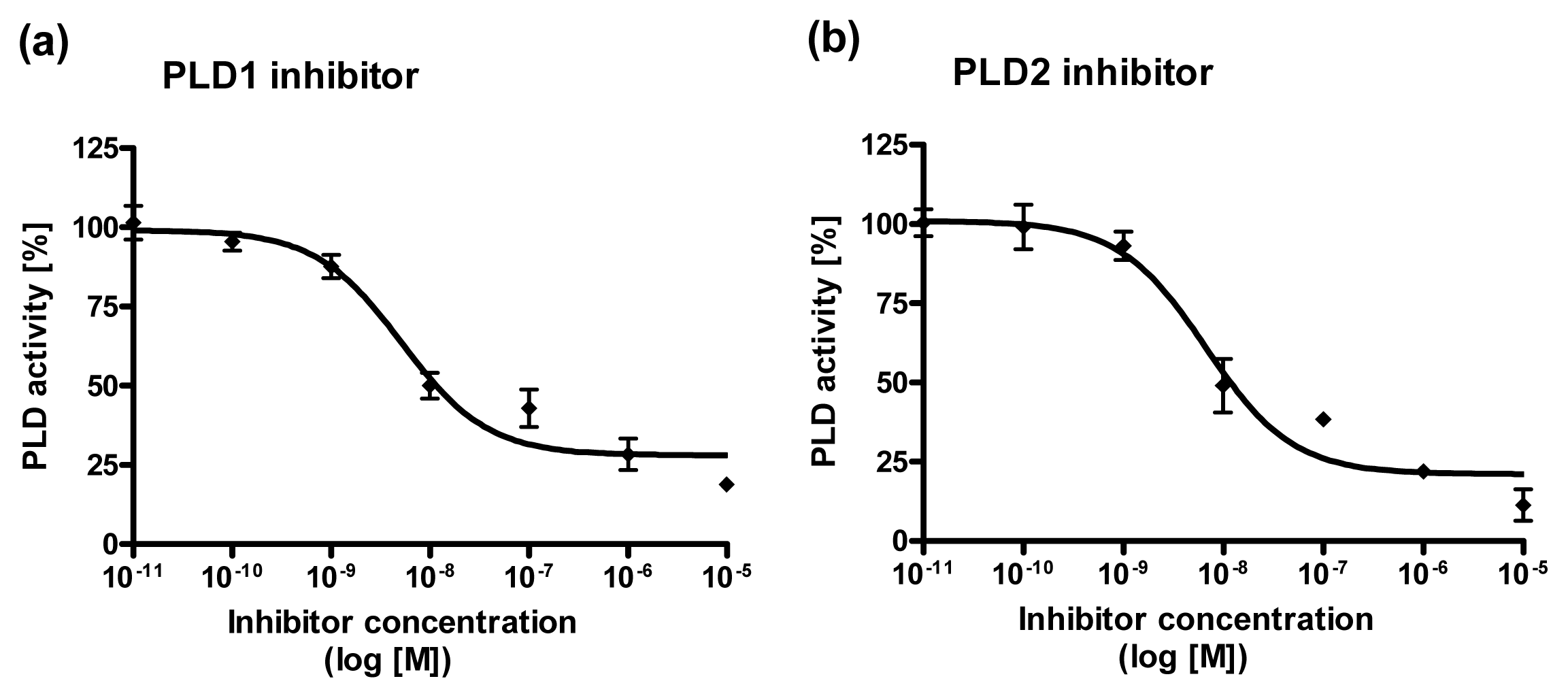
2.4. Dose Response Curves for PLD1 and PLD2 Inhibitors
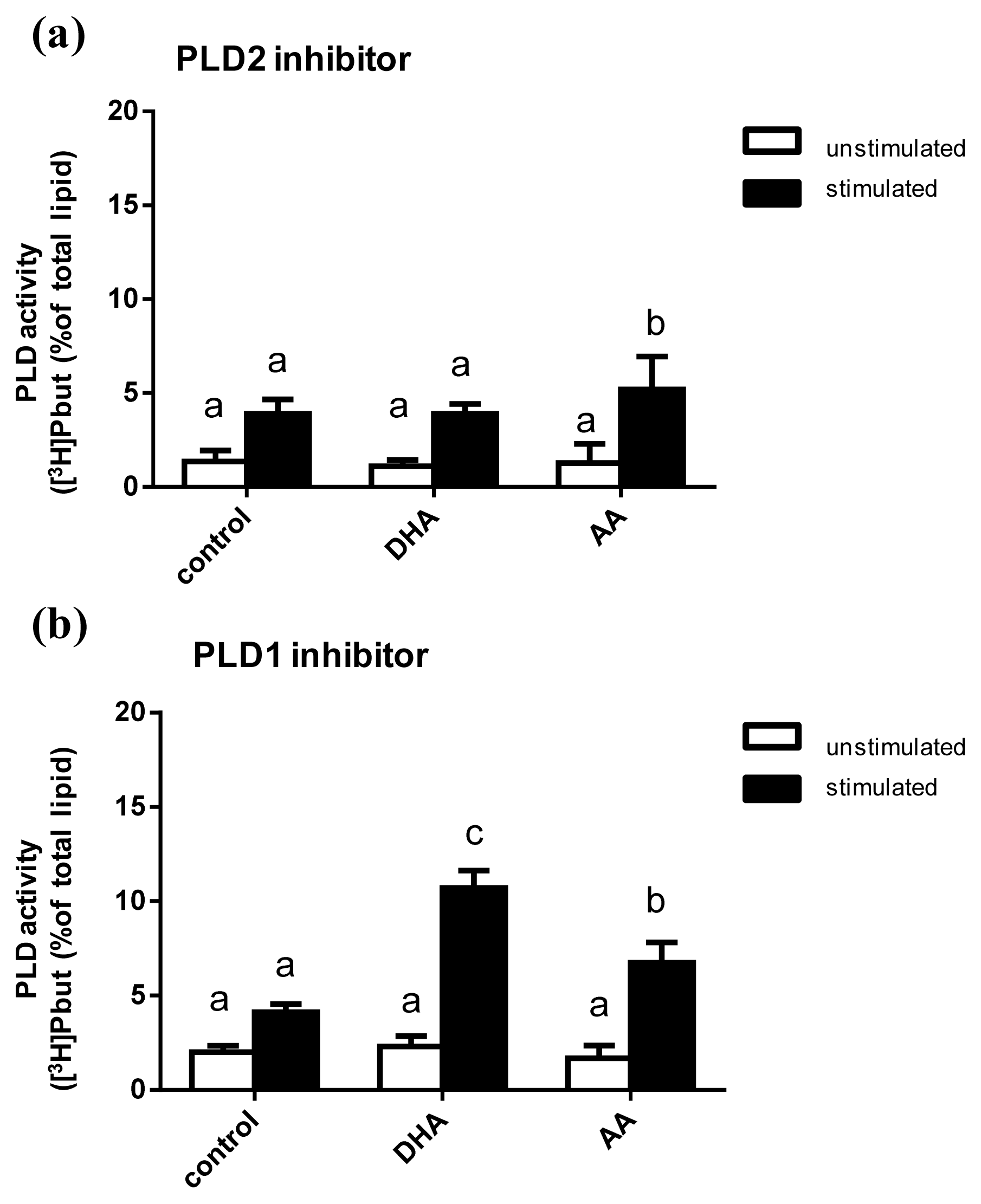
2.5. Influence of PUFA Supplementation on the Activity of PLD Isoforms
3. Experimental Section
3.1. Materials
3.2. Cell Culture
3.3. Transient Transfection
3.4. Confocal Microscopy
3.5. Measurement of PLD Activity
3.6. Selective Inhibition of PLD Isforms
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Basiouni, S.; Stockel, K.; Fuhrmann, H.; Schumann, J. Polyunsaturated fatty acid supplements modulate mast cell membrane microdomain composition. Cell. Immunol 2012, 275, 42–46. [Google Scholar]
- Gueck, T.; Seidel, A.; Fuhrmann, H. Effects of essential fatty acids on mediators of mast cells in culture. Prostaglandins Leukot. Essent. Fat. Acids 2003, 68, 317–322. [Google Scholar]
- Gueck, T.; Seidel, A.; Baumann, D.; Meister, A.; Fuhrmann, H. Alterations of mast cell mediator production and release by gamma-linolenic and docosahexaenoic acid. Vet. Dermatol 2004, 15, 309–314. [Google Scholar]
- Selvy, P.E.; Lavieri, R.R.; Lindsley, C.W.; Brown, H.A. Phospholipase D: Enzymology, functionality, and chemical modulation. Chem. Rev 2011, 111, 6064–6119. [Google Scholar]
- Exton, J.H. New developments in phospholipase D. J. Biol. Chem 1997, 272, 15579–15582. [Google Scholar]
- Brown, F.D.; Thompson, N.; Saqib, K.M.; Clark, J.M.; Powner, D.; Thompson, N.T.; Solari, R.; Wakelam, M.J.O. Phospholipase D1 localises to secretory granules and lysosomes and is plasma-membrane translocated on cellular stimulation. Curr. Biol 1998, 8, 835–838. [Google Scholar]
- Choi, W.S.; Kim, Y.M.; Combs, C.; Frohman, M.A.; Beaven, M.A. Phospholipases D1 and D2 regulate different phases of exocytosis in mast cells. J. Immunol 2002, 168, 5682–5689. [Google Scholar]
- Colley, W.C.; Sung, T.C.; Roll, R.; Jenco, J.; Hammond, S.M.; Altshuller, Y.; BarSagi, D.; Morris, A.J.; Frohman, M.A. Phospholipase D2, a distinct phospholipase D isoform with novel regulatory properties that provokes cytoskeletal reorganization. Curr. Biol 1997, 7, 191–201. [Google Scholar]
- Gemeinhardt, A.; Alfalah, M.; Guck, T.; Naim, H.Y.; Fuhrmann, H. The influence of linoleic and linolenic acid on the activity and intracellular localisation of phospholipase D in COS-1 cells. Biol. Chem 2009, 390, 253–258. [Google Scholar]
- Bechoua, S.; Dubois, M.; Nemoz, G.; Lagarde, M.; Prigent, A.F. Docosahexaenoic acid lowers phosphatidate level in human activated lymphocytes despite phospholipase D activation. J. Lipid Res 1998, 39, 873–883. [Google Scholar]
- Diaz, O.; Berquand, A.; Dubois, M.; di Agostino, S.; Sette, C.; Bourgoin, S.; Lagarde, M.; Nemoz, G.; Prigent, A.F. The mechanism of docosahexaenoic acid-induced phospholipase D activation in human lymphocytes involves exclusion of the enzyme from lipid rafts. J. Biol. Chem 2002, 277, 39368–39378. [Google Scholar]
- Grenier, S.; Flamand, N.; Pelletier, J.; Naccache, P.H.; Borgeat, P.; Bourgoin, S.G. Arachidonic acid activates phospholipase D in human neutrophils; essential role of endogenous leukotriene B-4 and inhibition by adenosine A(2A) receptor engagement. J. Leukoc. Biol 2003, 73, 530–539. [Google Scholar]
- Askari, B.; Carroll, M.A.; Capparelli, M.; Kramer, F.; Gerrity, R.G.; Bornfeldt, K.E. Oleate and linoleate enhance the growth-promoting effects of insulin-like growth factor-I through a phospholipase D-dependent pathway in arterial smooth muscle cells. J. Biol. Chem 2002, 277, 36338–36344. [Google Scholar]
- Wang, Y.T.; Oram, J.F. Unsaturated fatty acids phosphorylate and destabilize ABCA1 through a phospholipase D2 pathway. J. Biol. Chem 2005, 280, 35896–35903. [Google Scholar]
- Chahdi, A.; Choi, W.S.; Kim, Y.M.; Beaven, M.A. Mastoparan selectively activates phospholipase D2 in cell membranes. J. Biol. Chem 2003, 278, 12039–12045. [Google Scholar]
- Sarri, E.; Pardo, R.; Fensome-Green, A.; Cockcroft, S. Endogenous phospholipase D2 localizes to the plasma membrane of RBL-2H3 mast cells and can be distinguished from ADP ribosylation factor-stimulated phospholipase D1 activity by its specific sensitivity to oleic acid. Biochem. J 2003, 369, 319–329. [Google Scholar]
- Emoto, M.; Klarlund, J.K.; Waters, S.B.; Hu, V.; Buxton, J.M.; Chawla, A.; Czech, M.P. A role for phospholipase D in GLUT4 glucose transporter translocation. J. Biol. Chem 2000, 275, 7144–7151. [Google Scholar]
- Whatmore, J.; Morgan, C.P.; Cunningham, E.; Collison, K.S.; Willison, K.R.; Cockcroft, S. ADP-ribosylation factor 1-regulated phospholipase D activity is localized at the plasma membrane and intracellular organelles in HL60 cells. Biochem. J 1996, 320, 785–794. [Google Scholar]
- Ross, E.M.; Higashijima, T. Regulation of G-protein activation by mastoparans and other cationic peptides. Methods Enzymol 1994, 237, 26–37. [Google Scholar]
- Higashijima, T.; Burnier, J.; Ross, E.M. Regulation of Gi and Go by mastoparan, related amphiphilic peptides, and hydrophobic amines—Mechanism and structural determinants of activity. J. Biol. Chem 1990, 265, 14176–14186. [Google Scholar]
- Dedos, S.G.; Wicher, D.; Kaltofen, S.; Birkenbeil, H. Different Ca2+ signalling cascades manifested by mastoparan in the prothoracic glands of the tobacco hornworm, Manduca sexta, and the silkworm, Bombyx mori. Arch. Insect Biochem. Physiol 2007, 65, 52–64. [Google Scholar]
- Nakahata, N.; Ishimoto, H.; Mizuno, K.; Ohizumi, Y.; Nakanishi, H. Dual effects of mastoparan on intracellular free Ca2+ concentrations in human astrocytoma-cells. Br. J. Pharmacol 1994, 112, 299–303. [Google Scholar]
- Tobio, A.; Alfonso, A.; Botana, L.M. c-kit mutations and PKC crosstalks: PKC translocates to nucleous only in cells HMC560,816. J. Cell. Biochem 2011, 112, 2637–2651. [Google Scholar]
- Oka, M.; Kageshita, T.; Ono, T.; Goto, A.; Kuroki, T.; Ichihashi, M. Protein kinase C alpha associates with phospholipase D-1 and enhances basal phospholipase D activity in a protein phosphorylationindependent manner in human melanoma cells. J. Investig. Dermatol 2003, 121, 69–76. [Google Scholar]
- Lee, S.; Park, J.B.; Kim, J.H.; Kim, Y.; Kim, J.H.; Shin, K.J.; Lee, J.S.; Ha, S.H.; Suh, P.G.; Ryu, S.H. Actin directly interacts with phospholipase D, inhibiting its activity. J. Biol. Chem 2001, 276, 28252–28260. [Google Scholar]
- Garbi, M.; Rubinstein, S.; Lax, Y.; Breitbart, H. Activation of protein kinase C alpha in the lysophosphatidic acid-induced bovine sperm acrosome reaction and phospholipase D1 regulation. Biol. Reprod 2000, 63, 1271–1277. [Google Scholar]
- Hu, T.H.; Exton, J.H. Mechanisms of regulation of phospholipase D1 by protein kinase Ca. J. Biol. Chem 2003, 278, 2348–2355. [Google Scholar]
- Mochly-Rosen, D.; Henrich, C.J.; Cheever, L.; Khaner, H.; Simpson, P.C. A protein kinase C isozyme is translocated to cytoskeletal elements on activation. Cell Regul 1990, 1, 693–706. [Google Scholar]
- Nair, S.S.D.; Leitch, J.; Garg, M.L. N-3 polyunsaturated fatty acid supplementation alters inositol phosphate metabolism and protein kinase C activity in adult porcine cardiac myocytes. J. Nutr. Biochem 2001, 12, 7–13. [Google Scholar]
- Denys, A.; Hichami, A.; Khan, N.A. n-3PUFAs modulate T-cell activation via protein kinase C-alpha and -epsilon and the NF-kappa B signaling pathway. J. Lipid Res 2005, 46, 752–758. [Google Scholar]
- Khan, W.A.; Blobe, G.C.; Hannun, Y.A. Arachidonic-acid and free fatty-acids as 2nd-messengers and the role of protein-kinase-C. Cell. Signal 1995, 7, 171–184. [Google Scholar]
- Lopez-Nicolas, R.; Lopez-Andreo, M.J.; Marin-Vicente, C.; Gomez-Fernandez, J.C.; Corbalan-Garcia, S. Molecular mechanisms of PKCalpha localization and activation by arachidonic acid. The C2 domain also plays a role. J Mol. Biol 2006, 357, 1105–1120. [Google Scholar]
- Divecha, N.; Roefs, M.; Halstead, J.R.; D’Andrea, S.; Fernandez-Borga, M.; Oomen, L.; Saqib, K.M.; Wakelam, M.J.; D’Santos, C. Interaction of the type Ialpha PIPkinase with phospholipase D: A role for the local generation of phosphatidylinositol 4, 5-bisphosphate in the regulation of PLD2 activity. EMBO J 2000, 19, 5440–5449. [Google Scholar]
- Lavieri, R.R.; Scott, S.A.; Selvy, P.E.; Kim, K.; Jadhav, S.; Morrison, R.I.; Daniels, J.S.; Brown, H.A.; Lindsley, C.W. Design, synthesis, and biological evaluation of halogenated N-(2-(4-Oxo-1-phenyl-1,3,8-triazaspiro[4.5]decan-8-yl)ethyl)benzamides: Discovery of an isoform-selective small molecule phospholipase D2 inhibitor. J. Med. Chem 2010, 53, 6706–6719. [Google Scholar]
- Scott, S.A.; Selvy, P.E.; Buck, J.R.; Cho, H.P.; Criswell, T.L.; Thomas, A.L.; Armstrong, M.D.; Arteaga, C.L.; Lindsley, C.W.; Brown, H.A. Design of isoform-selective phospholipase D inhibitors that modulate cancer cell invasiveness. Nat. Chem. Biol 2009, 5, 108–117. [Google Scholar]
- Araki, S.; Ito, M.; Kureishi, Y.; Feng, J.H.; Machida, H.; Isaka, N.; Amano, M.; Kaibuchi, K.; Hartshorne, D.J.; Nakano, T. Arachidonic acid-induced Ca2+ sensitization of smooth muscle contraction through activation of Rho-kinase. Pflugers Arch.-Eur. J. Physiol 2001, 441, 596–603. [Google Scholar]
- Fu, X.; Gong, M.C.; Jia, T.; Somlyo, A.V.; Somlyo, A.P. The effects of the Rho-kinase inhibitor Y-27632 on arachidonic acid-, GTPgammaS-, and phorbol ester-induced Ca2+-sensitization of smooth muscle. FEBS Lett 1998, 440, 183–187. [Google Scholar]
- Young, C.; Gean, P.W.; Chiou, L.C.; Shen, Y.Z. Docosahexaenoic acid inhibits synaptic transmission and epileptiform activity in the rat hippocampus. Synapse 2000, 37, 90–94. [Google Scholar]
- Yi, L.; Zhang, Q.Y.; Mi, M.T. Role of Rho GTPase in inhibiting metastatic ability of human prostate cancer cell line PC-3 by omega-3 polyunsaturated fatty acid. Ai Zheng 2007, 26, 1281–1286. [Google Scholar]
- Duraisamy, Y.; Lambert, D.; O’Neill, C.A.; Padfield, P.J. Differential incorporation of docosahexaenoic acid into distinct cholesterol-rich membrane raft domains. Biochem. Biophys. Res. Commun 2007, 360, 885–890. [Google Scholar]
- Devinney, R.; Gold, W.M. Establishment of 2 Dog mastocytoma cell-lines in continuous culture. Am. J. Respir. Cell Mol. Biol 1990, 3, 413–420. [Google Scholar]
- Lazarus, S.C.; Mccabe, L.J.; Nadel, J.A.; Gold, W.M.; Leikauf, G.D. Effects of mast cell-derived mediators on epithelial-cells in canine trachea. Am. J. Physiol 1986, 251, C387–C394. [Google Scholar]
- Basiouni, S.; Fuhrmann, H.; Schumann, J. High-efficiency transfection of suspension cell lines. Biotechniques 2012, 1–4. [Google Scholar] [CrossRef]


© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Basiouni, S.; Fuhrmann, H.; Schumann, J. The Influence of Polyunsaturated Fatty Acids on the Phospholipase D Isoforms Trafficking and Activity in Mast Cells. Int. J. Mol. Sci. 2013, 14, 9005-9017. https://doi.org/10.3390/ijms14059005
Basiouni S, Fuhrmann H, Schumann J. The Influence of Polyunsaturated Fatty Acids on the Phospholipase D Isoforms Trafficking and Activity in Mast Cells. International Journal of Molecular Sciences. 2013; 14(5):9005-9017. https://doi.org/10.3390/ijms14059005
Chicago/Turabian StyleBasiouni, Shereen, Herbert Fuhrmann, and Julia Schumann. 2013. "The Influence of Polyunsaturated Fatty Acids on the Phospholipase D Isoforms Trafficking and Activity in Mast Cells" International Journal of Molecular Sciences 14, no. 5: 9005-9017. https://doi.org/10.3390/ijms14059005