Eviprostat Activates cAMP Signaling Pathway and Suppresses Bladder Smooth Muscle Cell Proliferation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
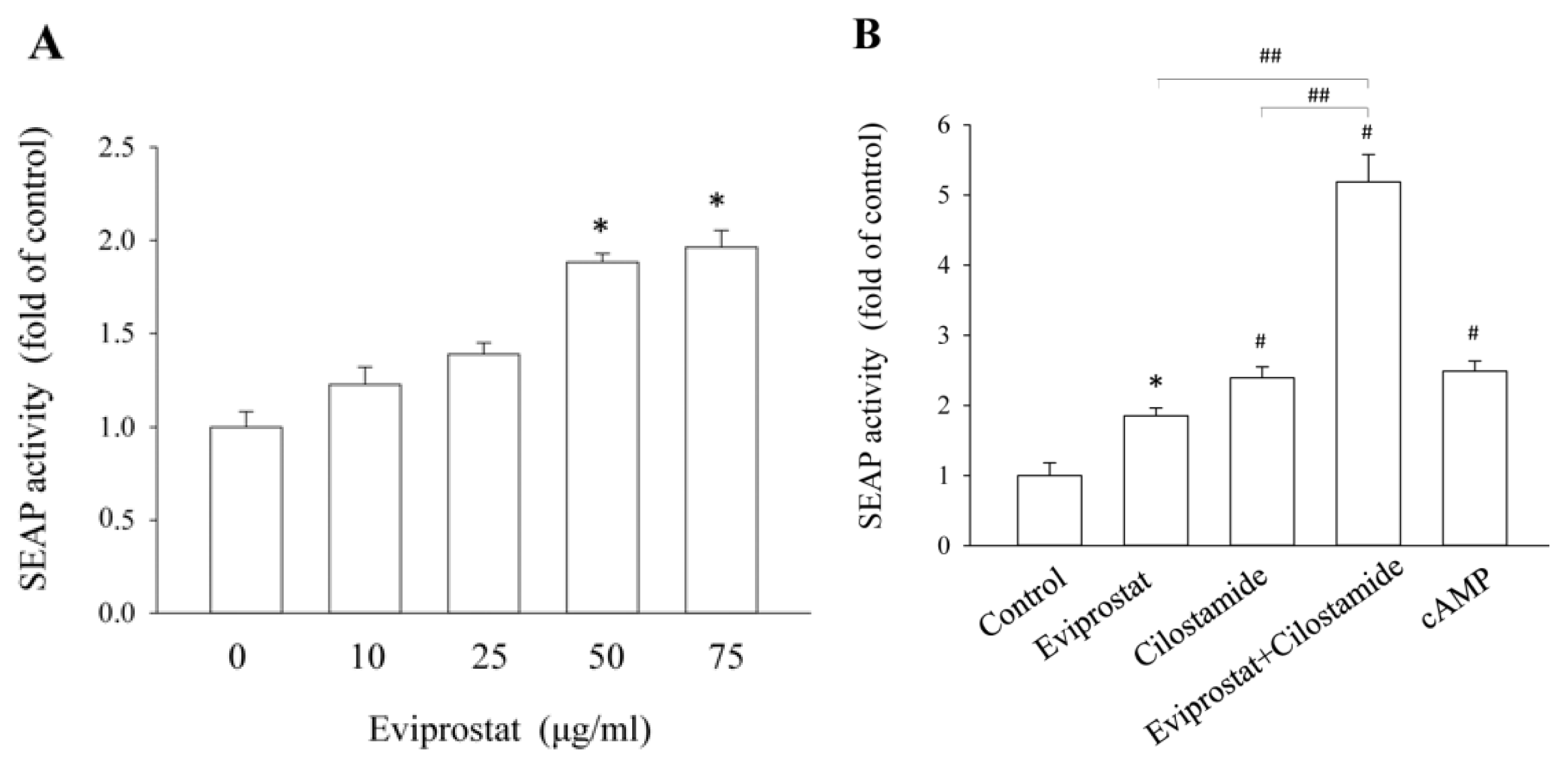
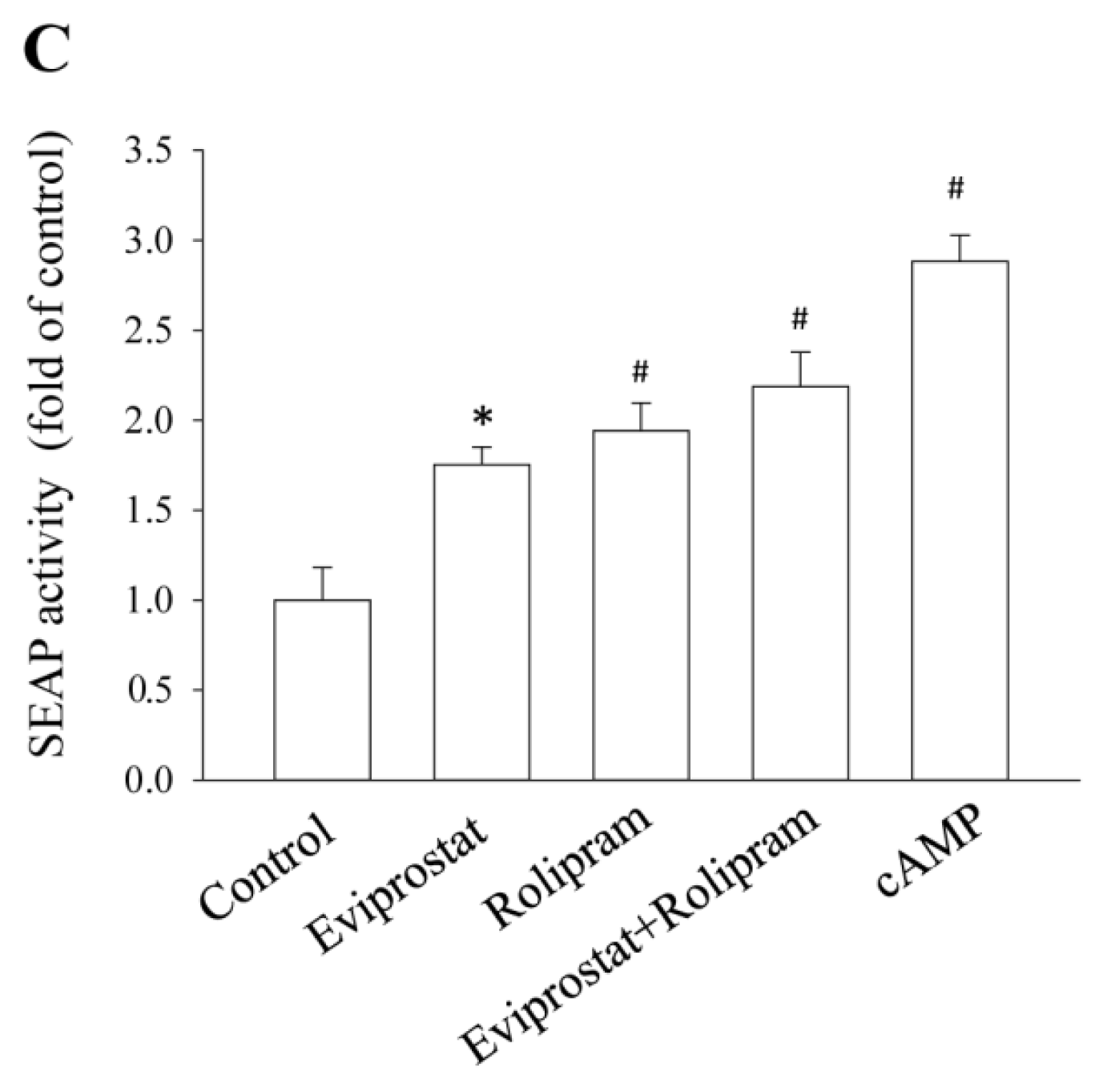
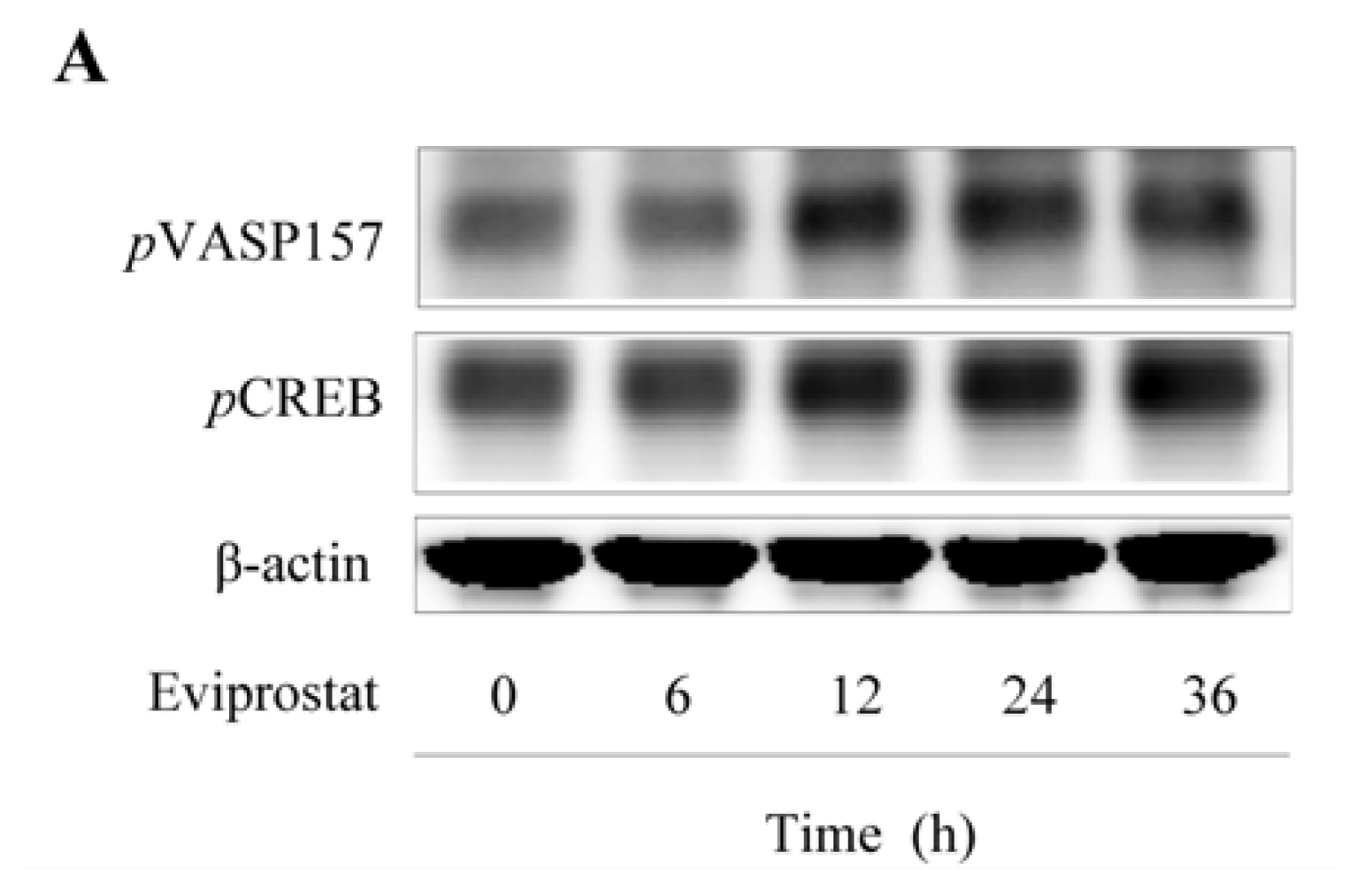
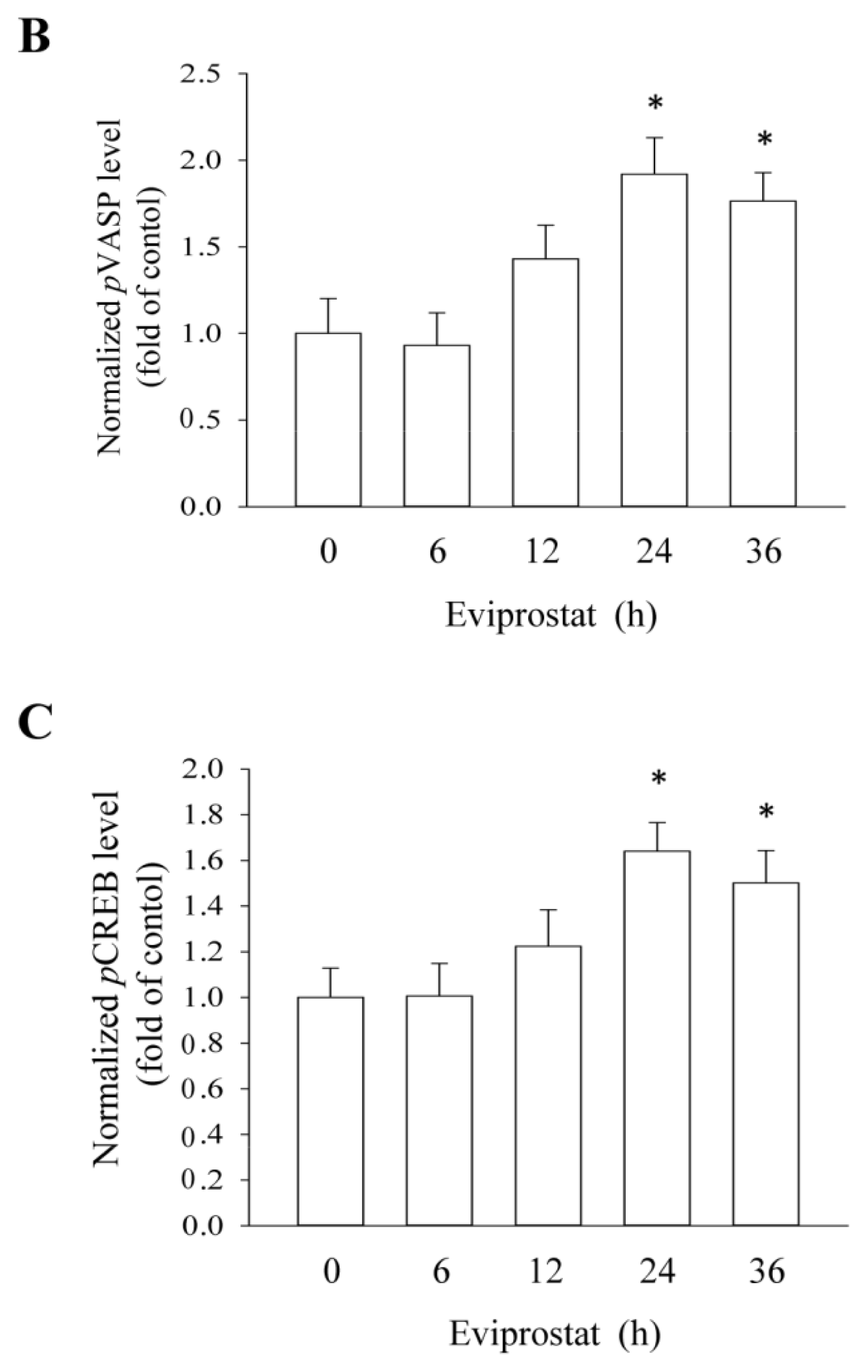
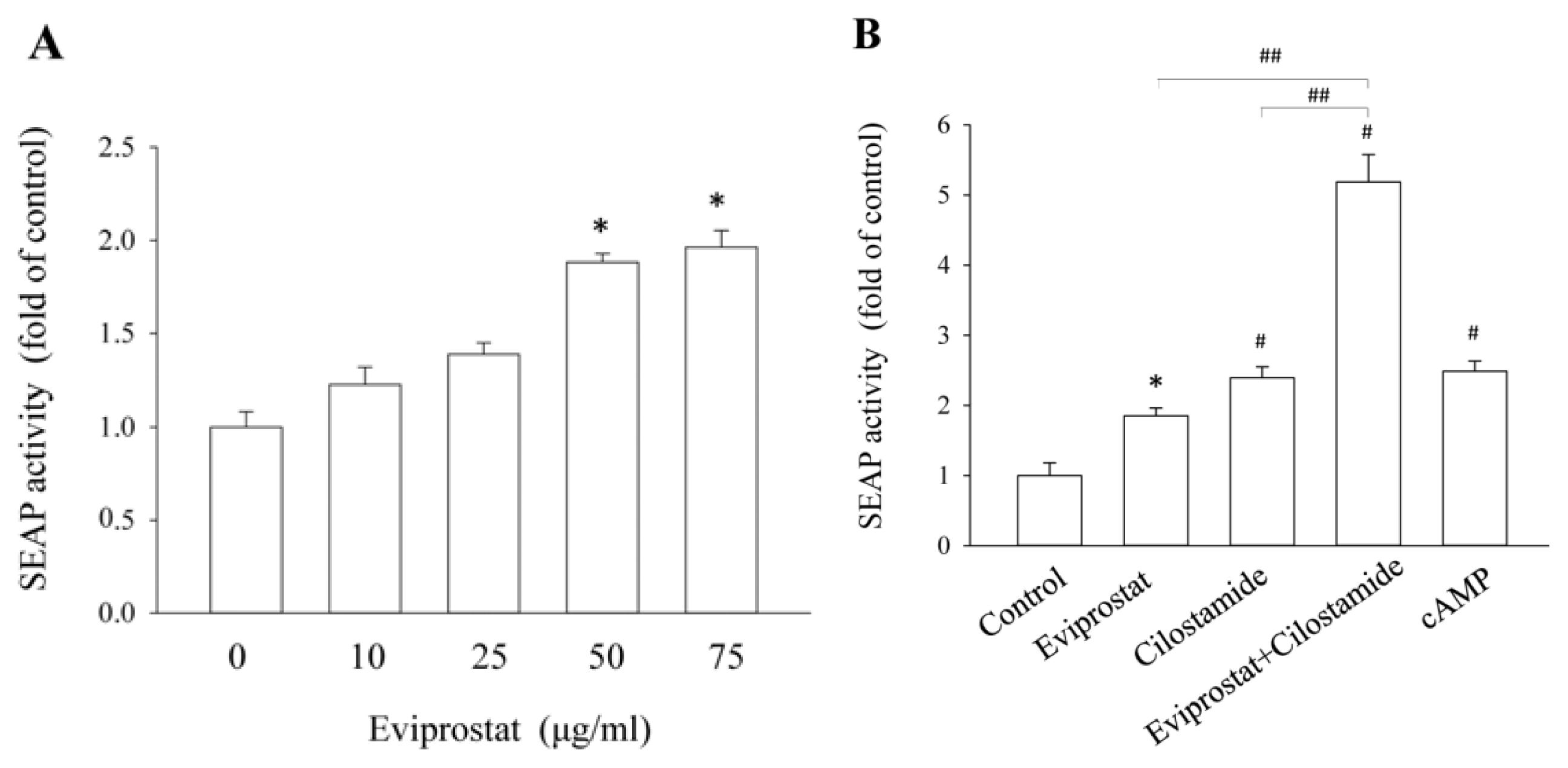
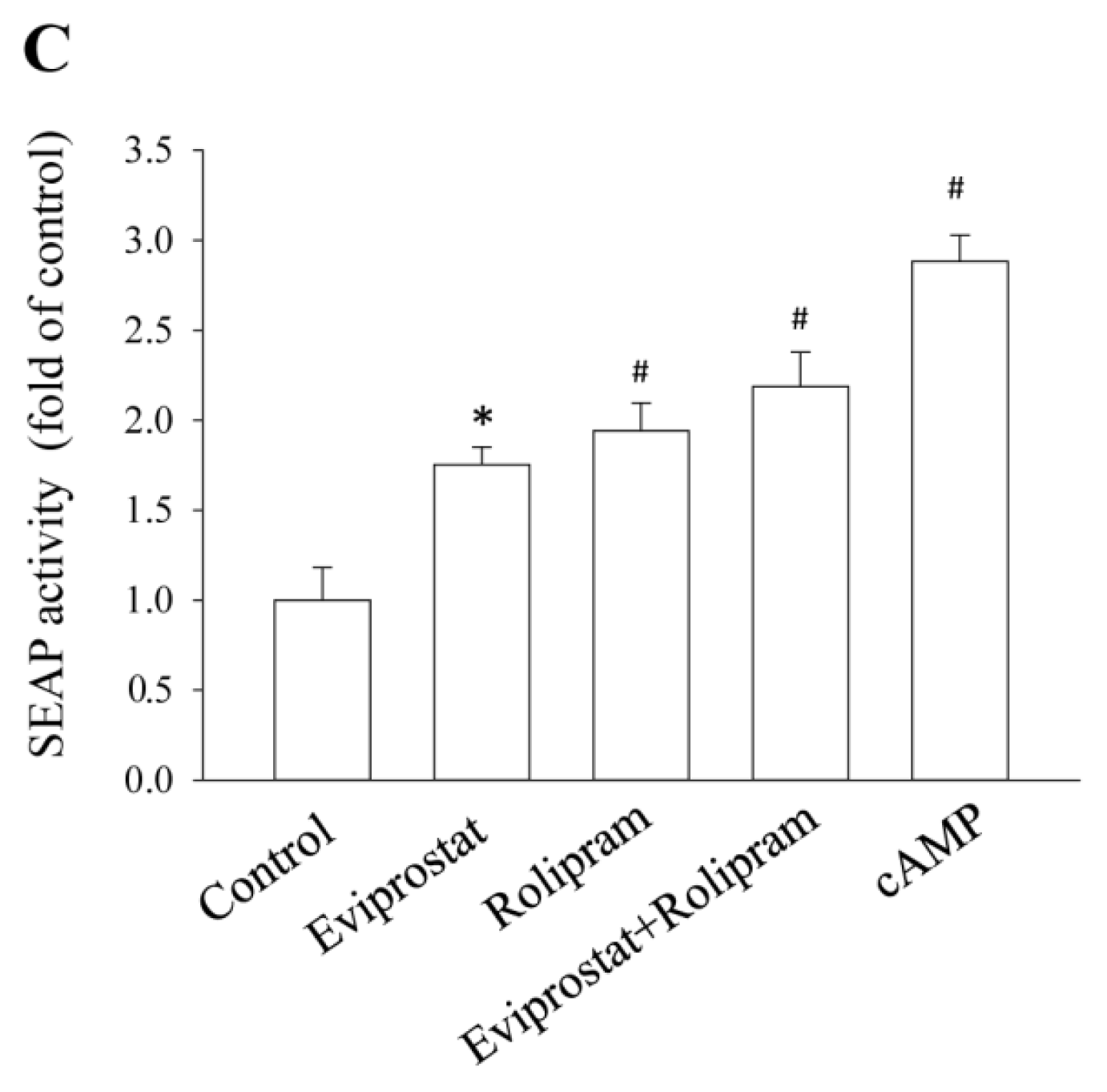
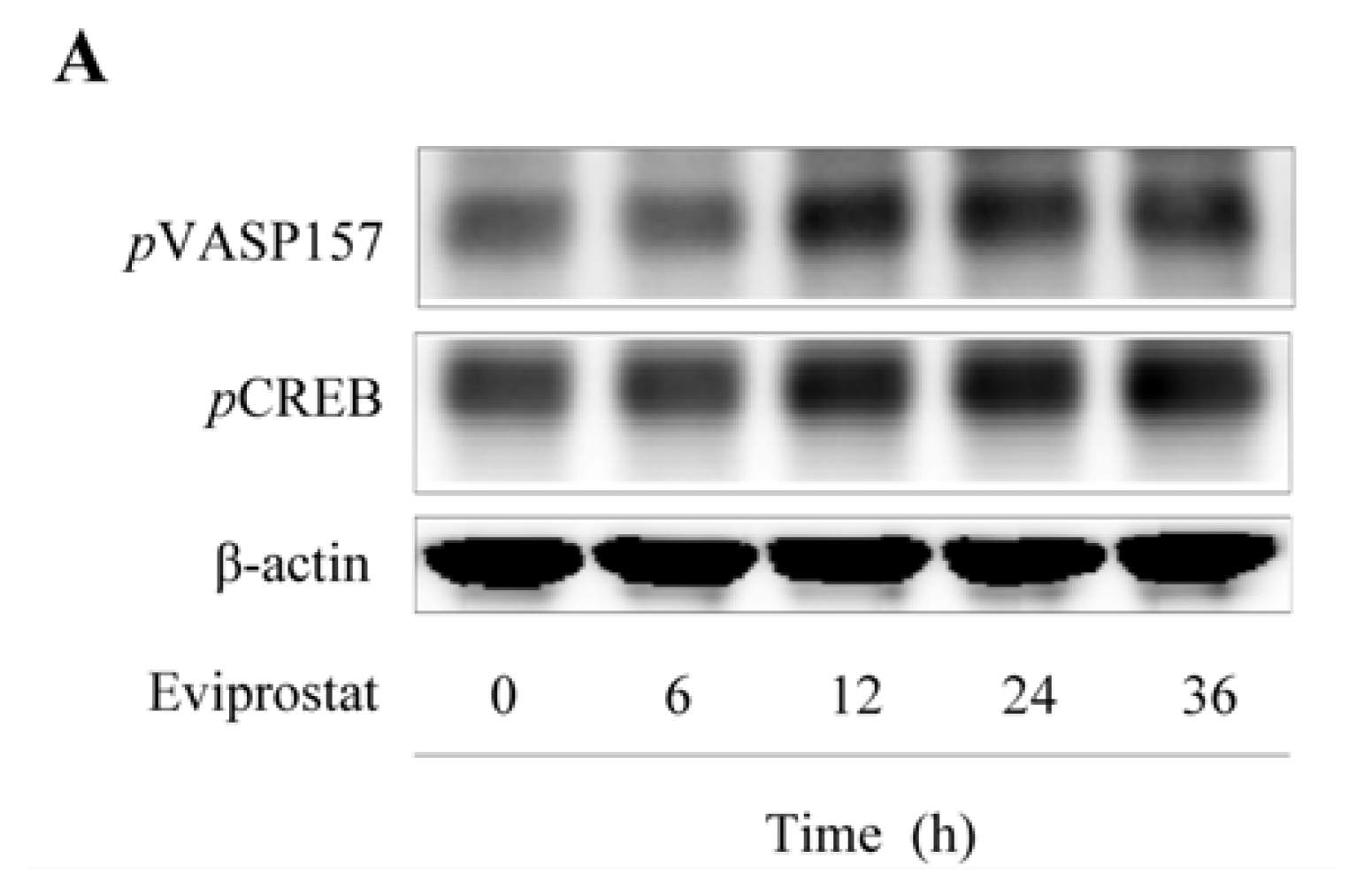
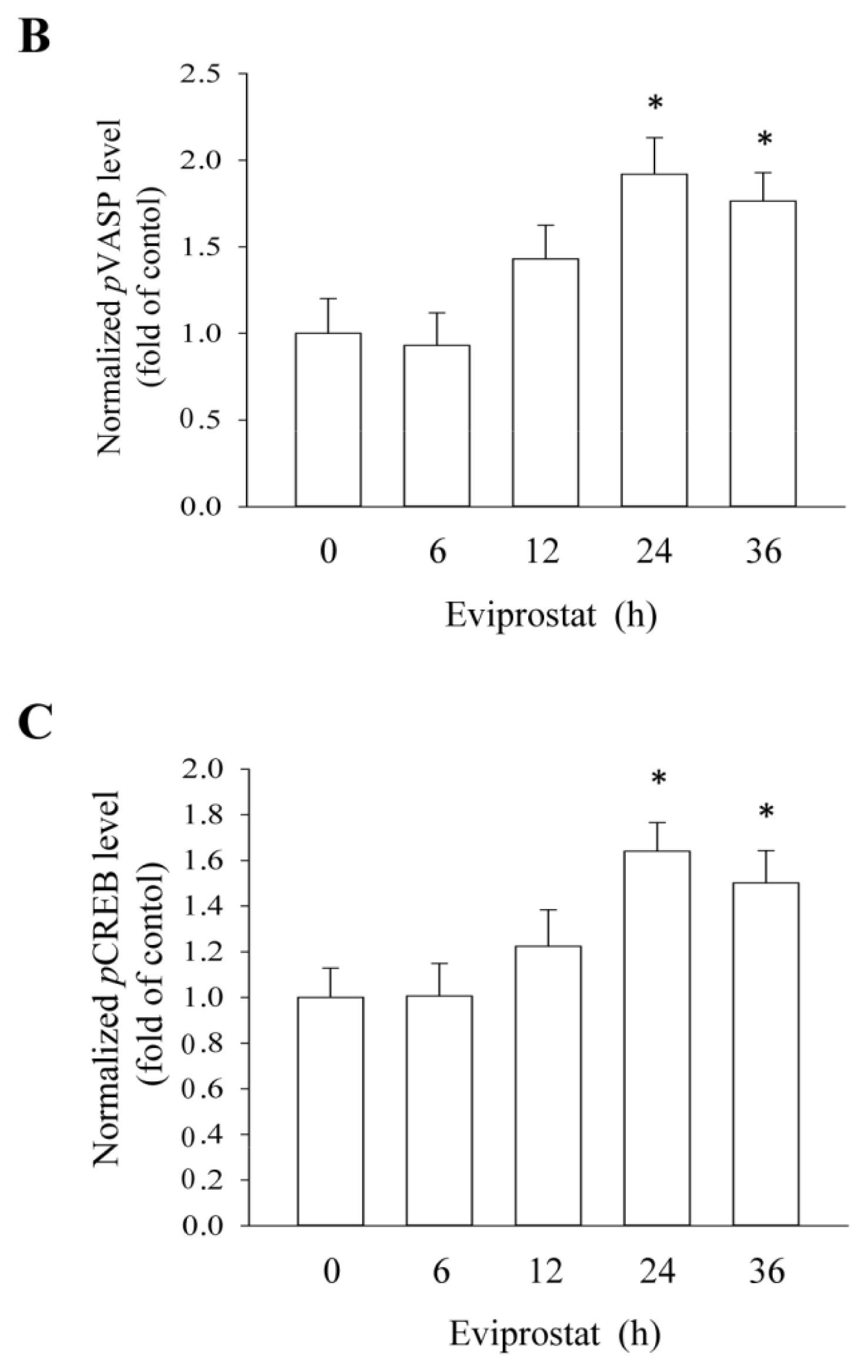
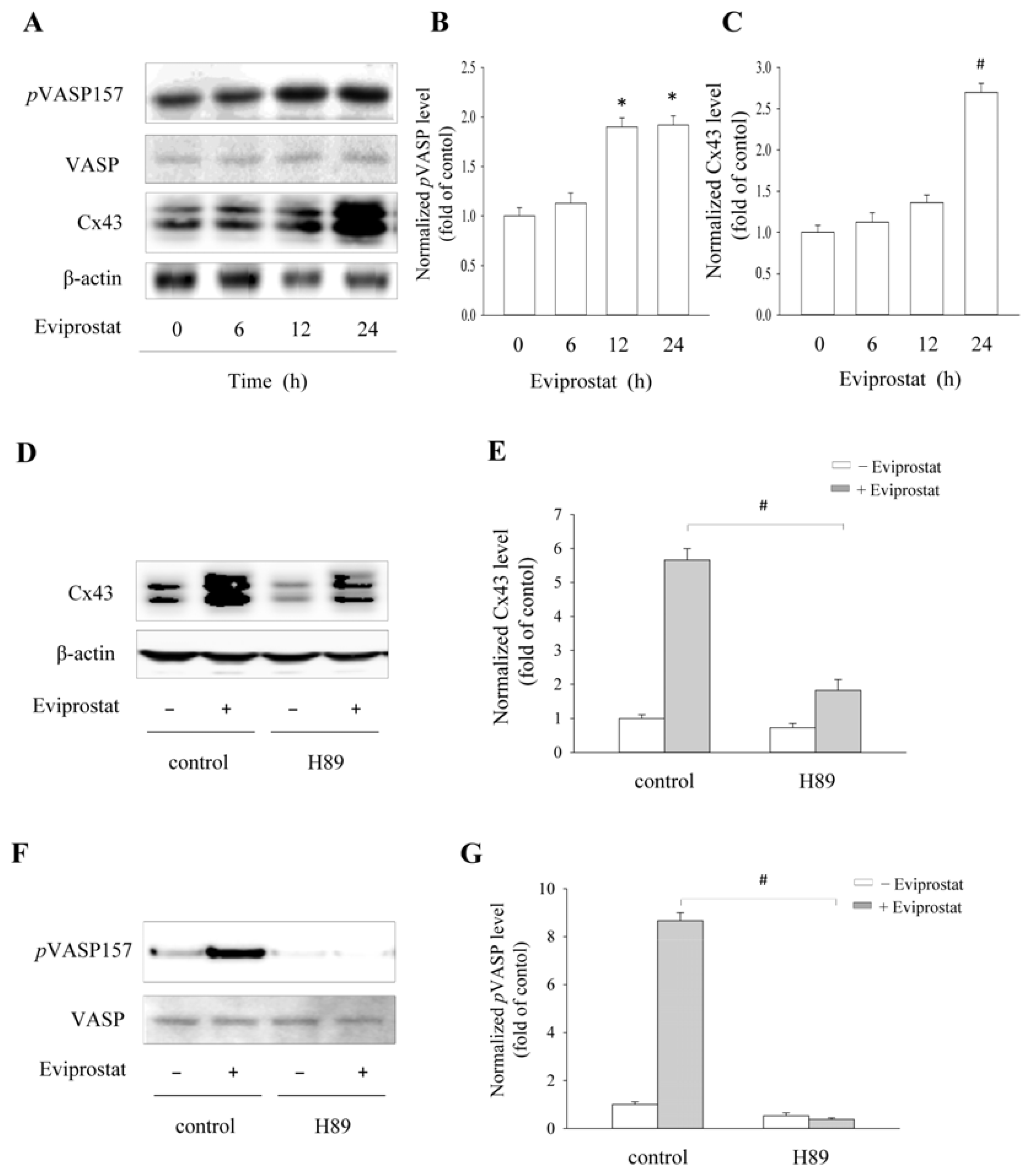
2.1. Eviprostat Activates cAMP Signaling Pathway
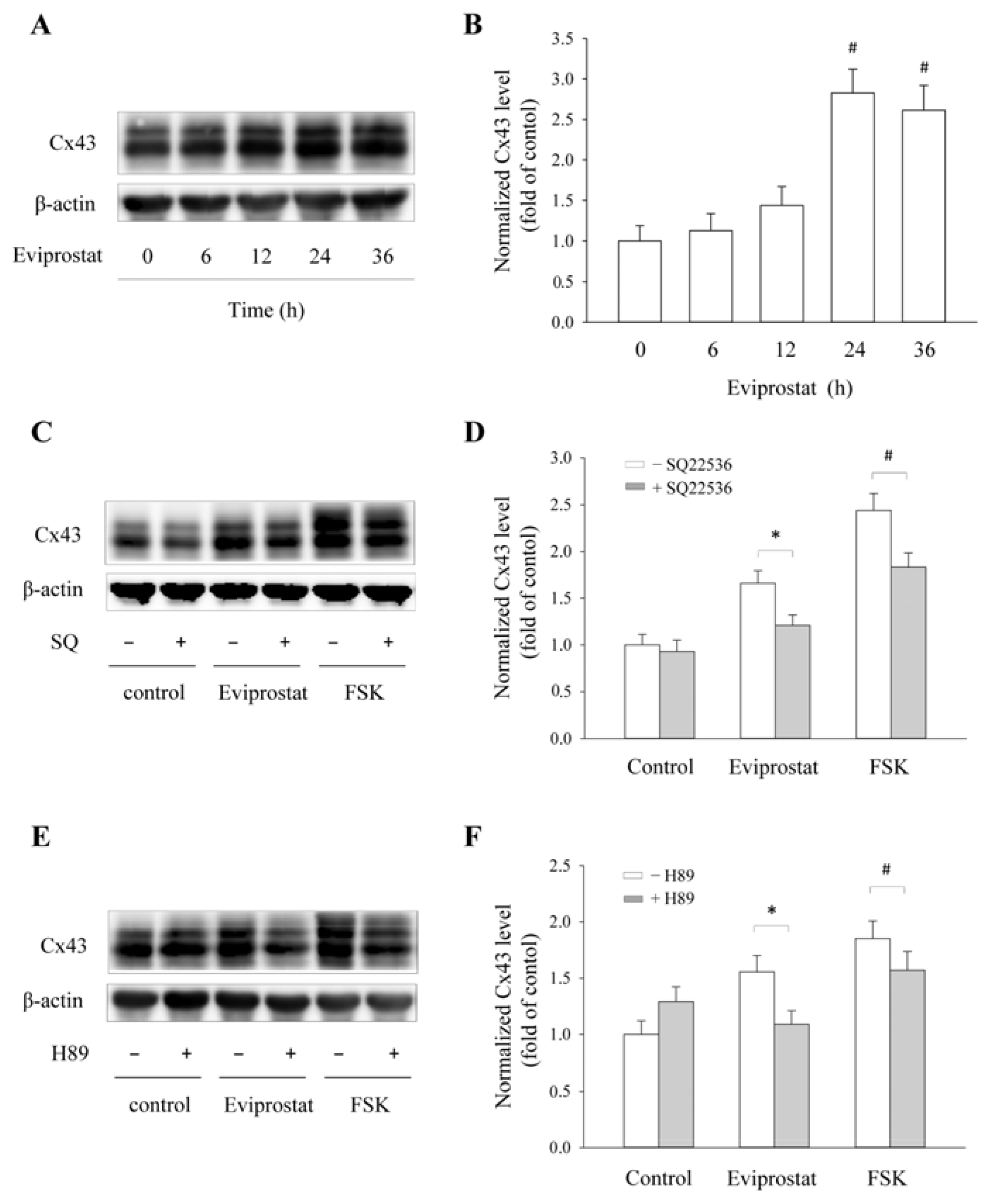
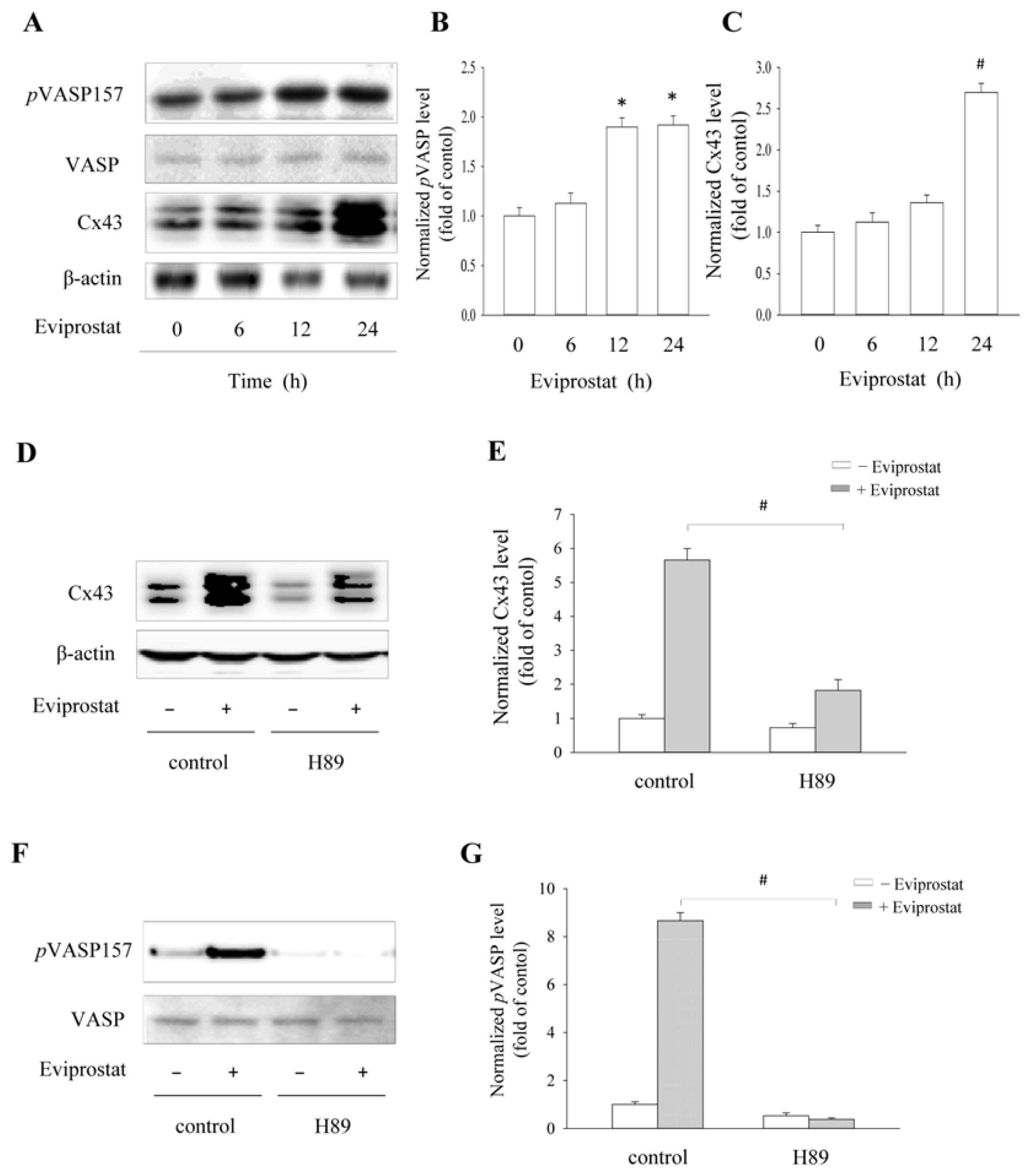
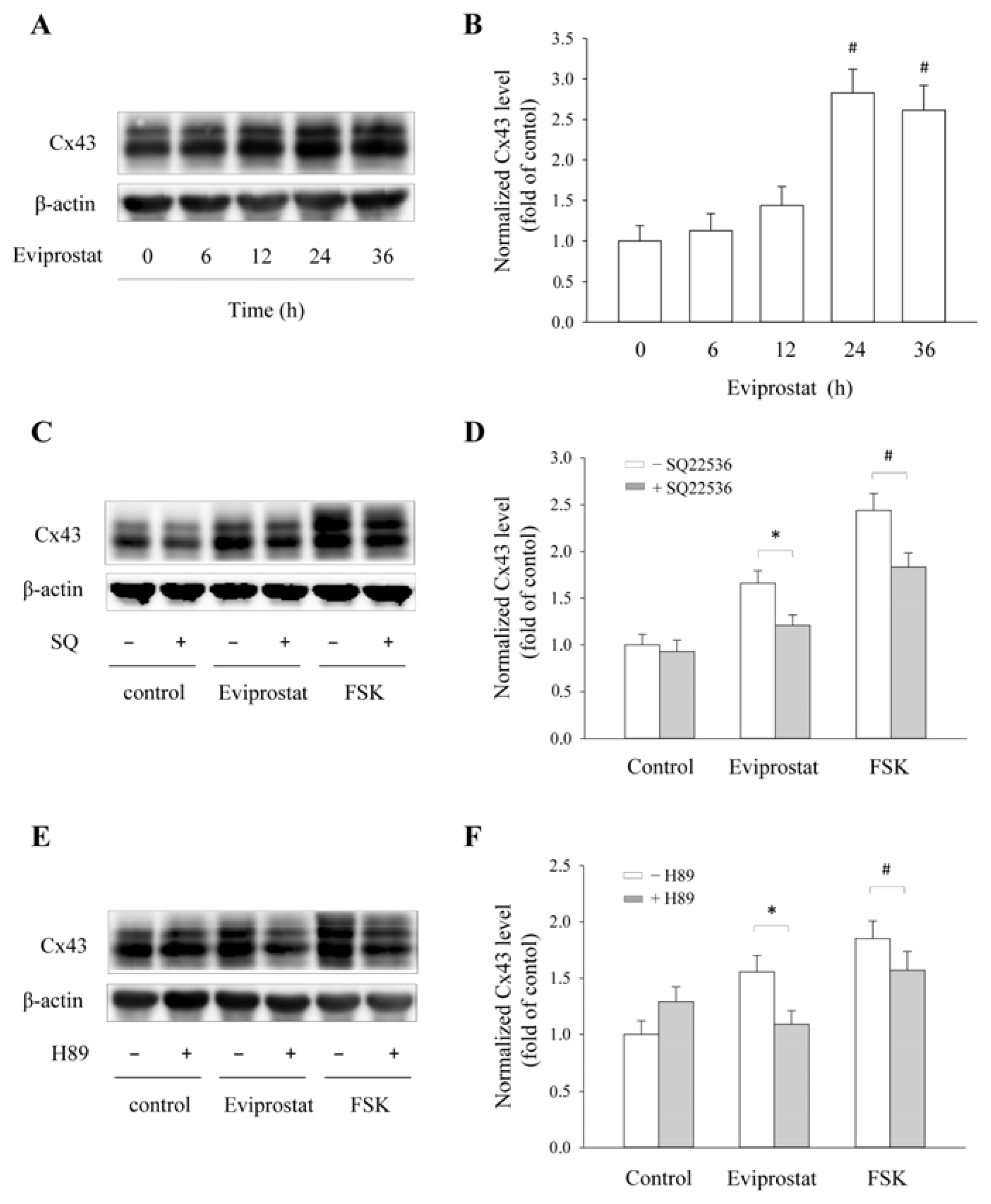
2.2. Eviprostat Elevates Cx43 Expression via cAMP Signaling Pathway
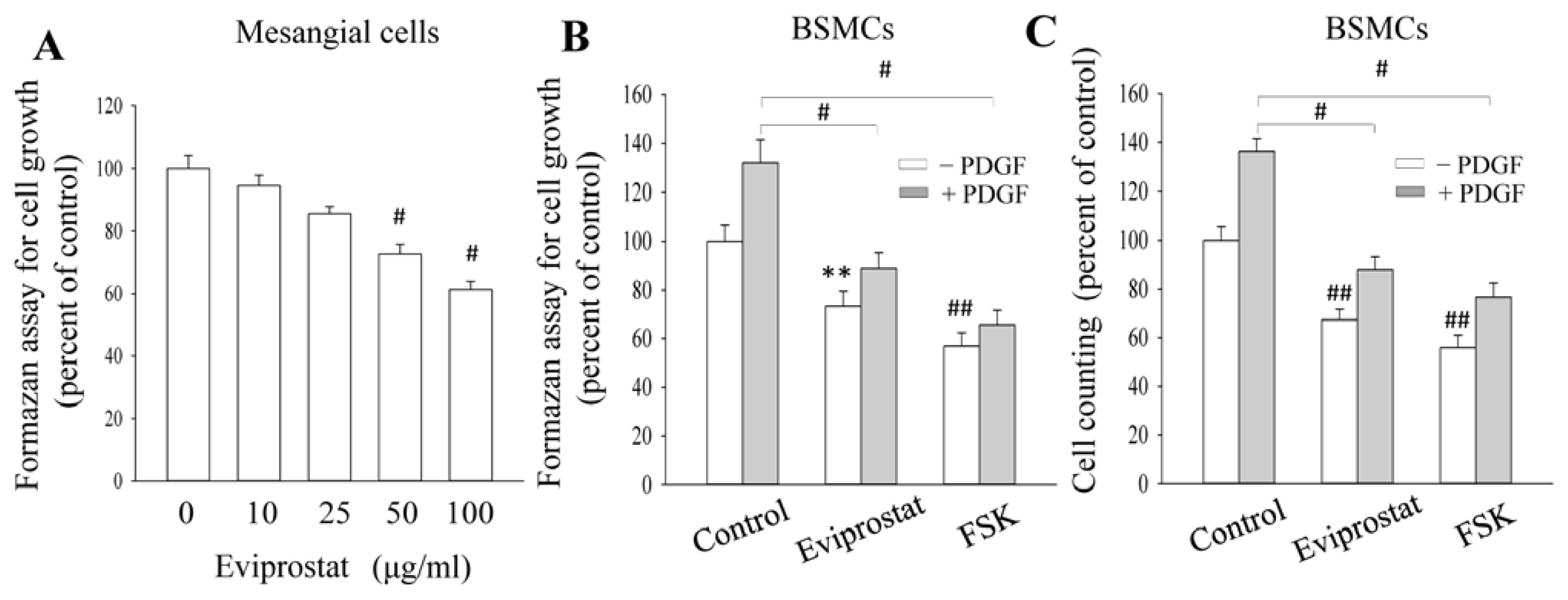
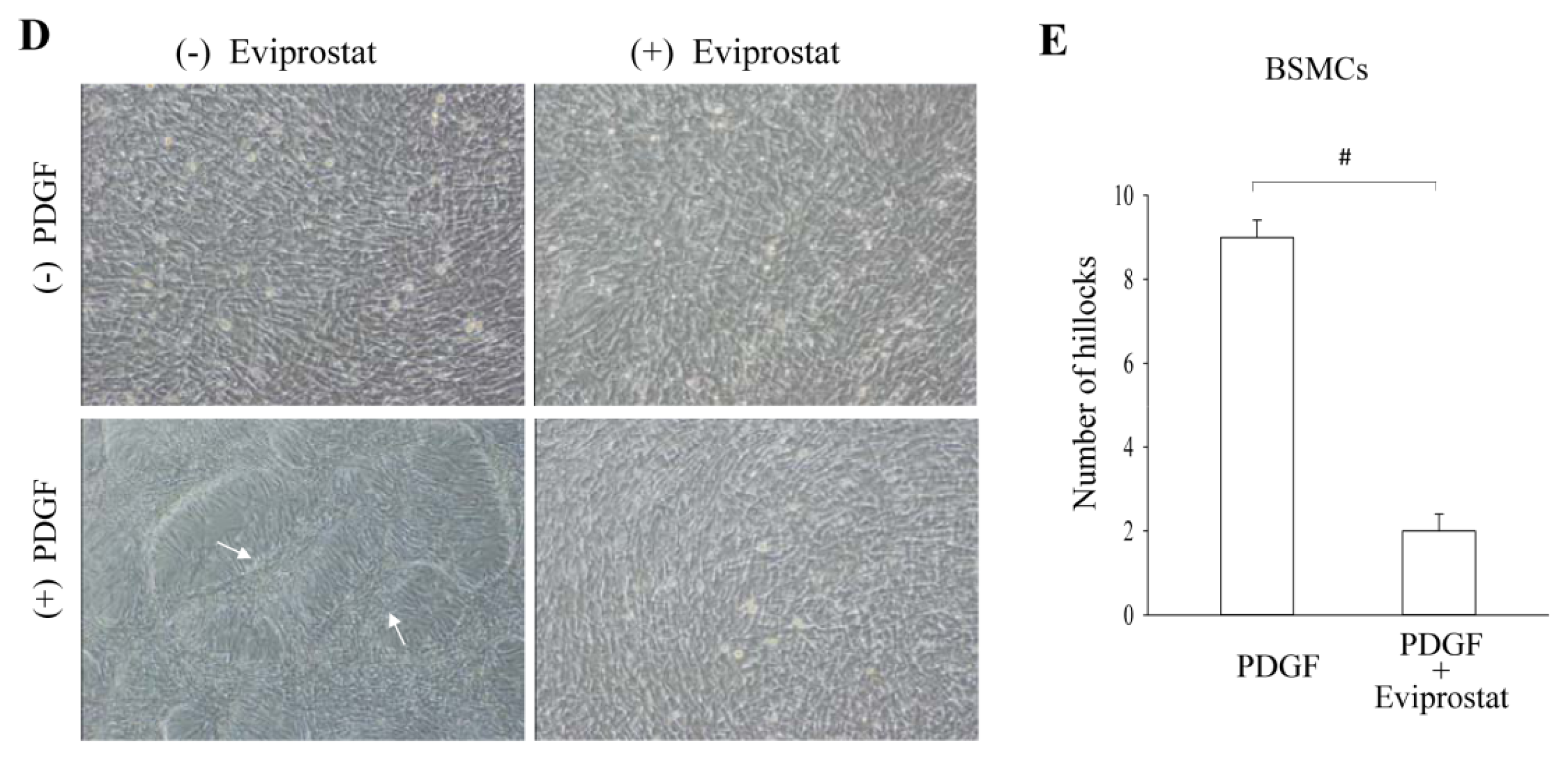
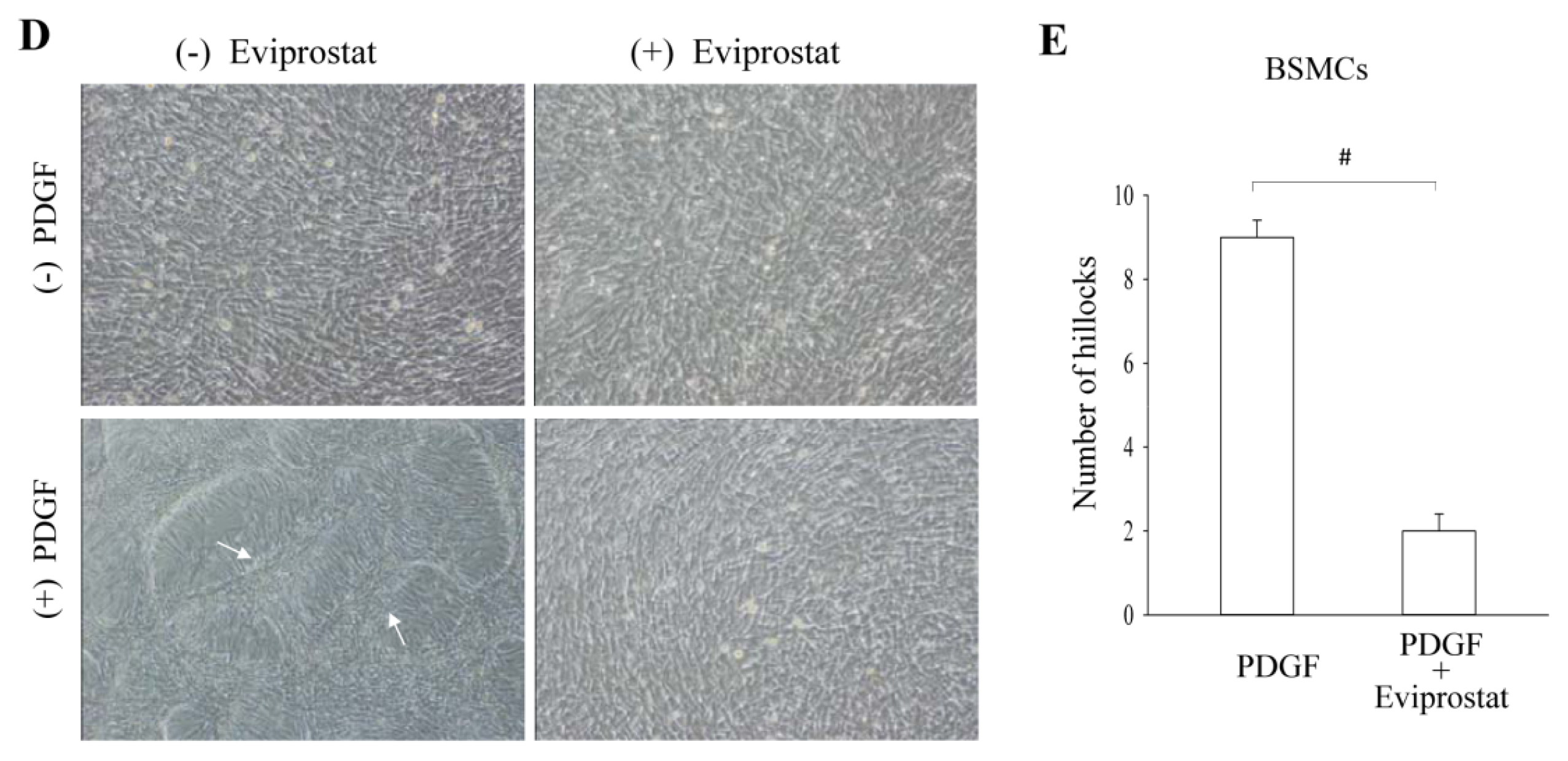
2.3. Eviprostat Suppresses PDGF-Induced Cellular Proliferation
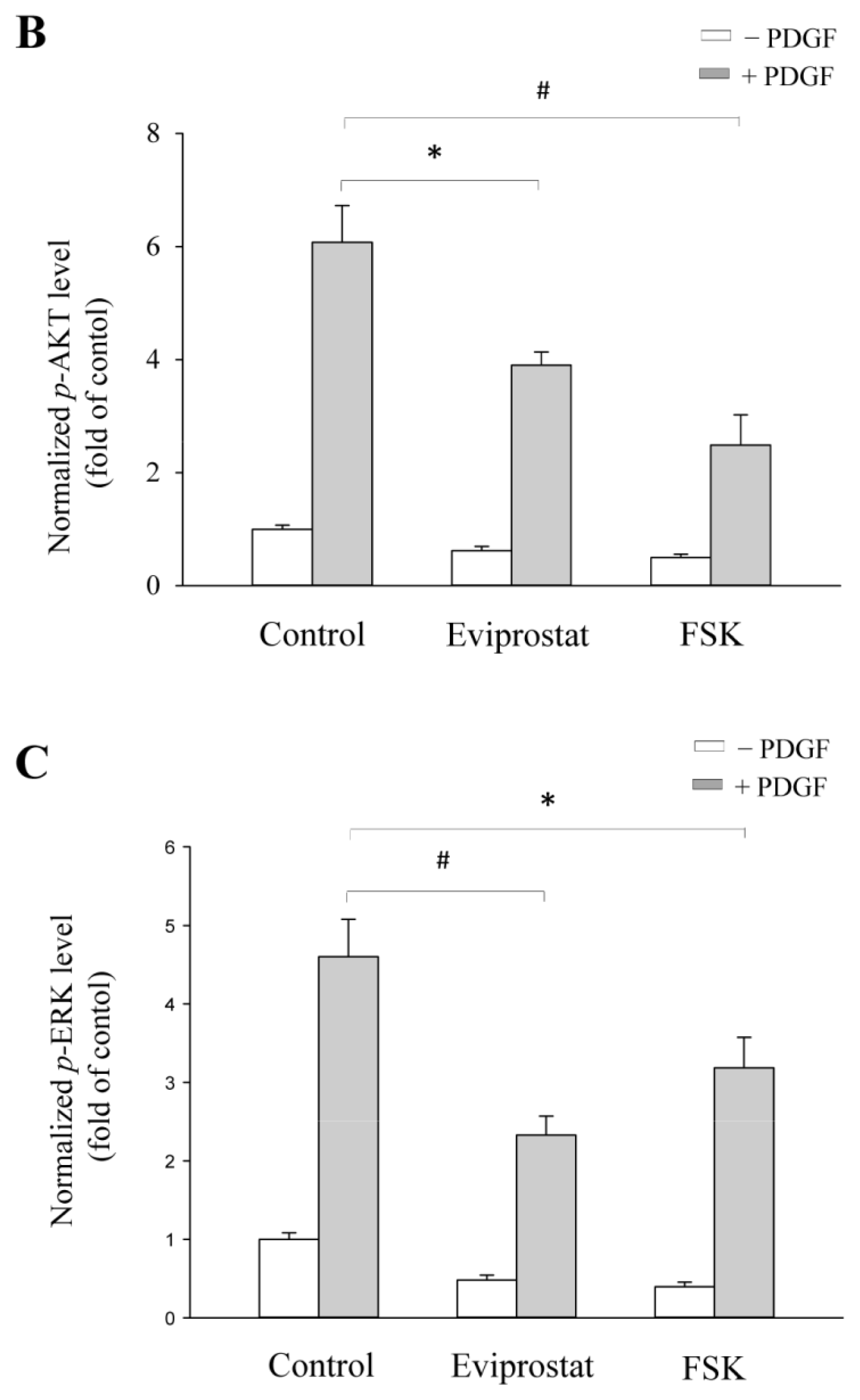
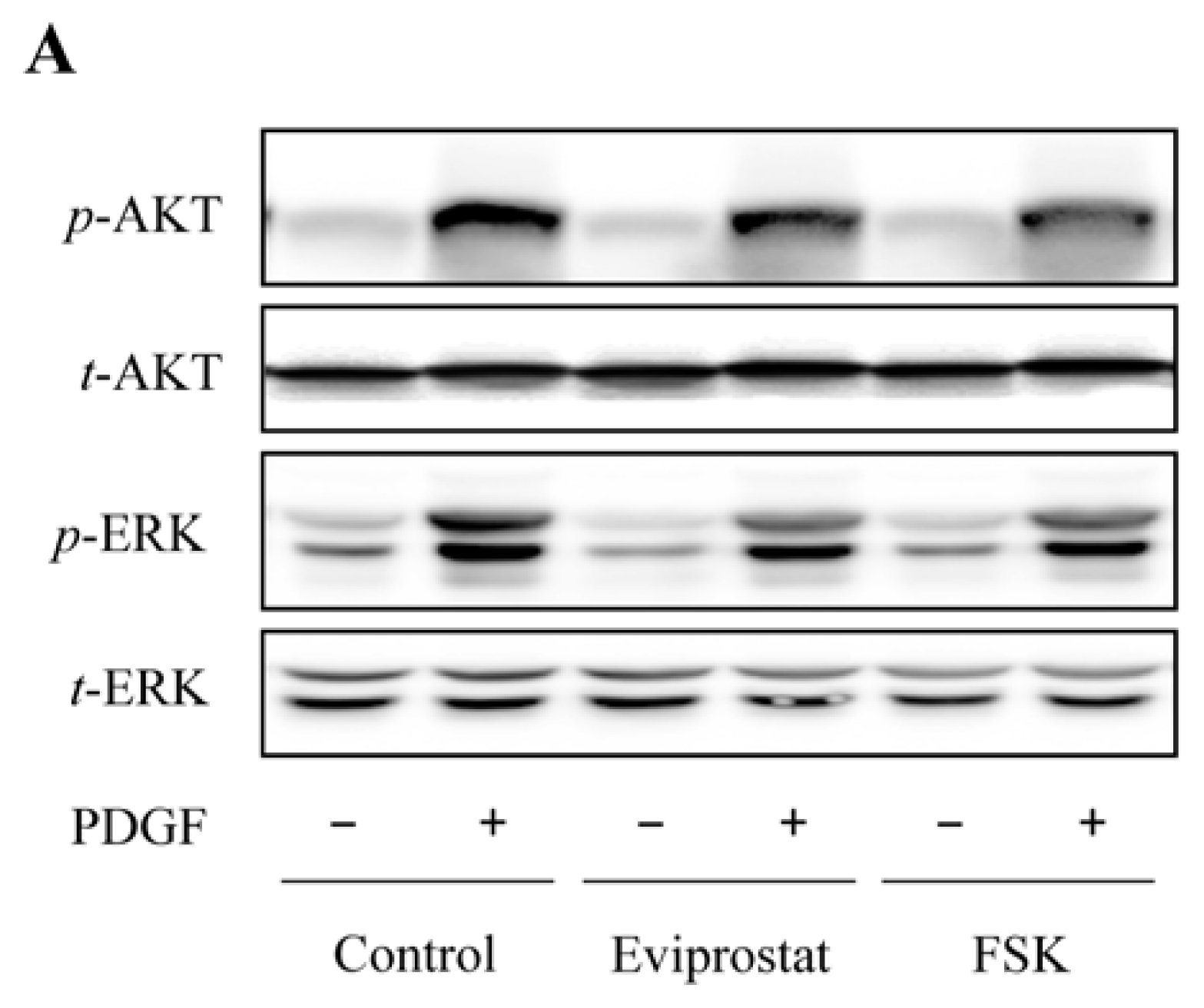
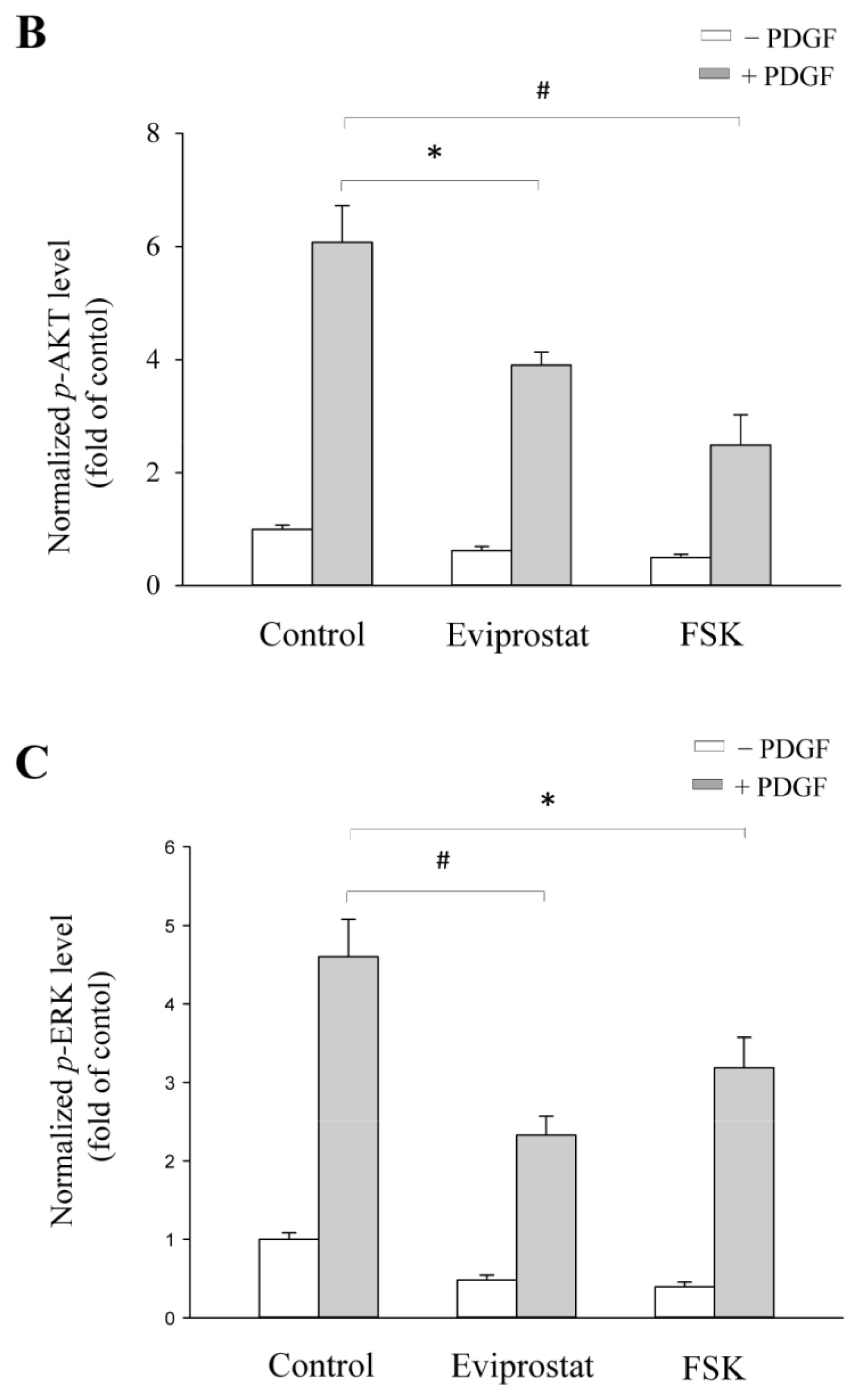
2.4. Eviprostat Suppresses PDGF-Induced Activation of PI3K and MAPK
2.5. Eviprostat Increases Cx43 Levels via cAMP Signaling Pathway in BSMCs
2.6. Discussion
3. Experimental Section
3.1. Reagents
3.2. Establishment of Reporter Cells
3.3. Bladder Smooth Muscle Cell Culture
3.4. Western Blot Analysis
3.5. SEAP Assay
3.6. Formazan Assay
3.7. Cell Counting
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Berry, S.J.; Coffey, D.S.; Walsh, P.C.; Ewing, L.L. The development of human benign prostatic hyperplasia with age. J. Urol 1984, 132, 474–479. [Google Scholar]
- Lee, K.L.; Peehl, D.M. Molecular and cellular pathogenesis of benign prostatic hyperplasia. J. Urol 2004, 172, 1784–1791. [Google Scholar]
- Ishigooka, M.; Hashimoto, T.; Hayami, S.; Tomaru, M.; Nakada, T.; Mitobe, K. Clinical and retrospective evaluation of Eviprostat: A non-hormonal and non-neuropharmacological agent for benign prostatic hyperplasia. Int. Urol. Nephrol 1995, 27, 61–66. [Google Scholar]
- Oka, M.; Tachibana, M.; Noda, K.; Inoue, N.; Tanaka, M.; Kuwabara, K. Relevance of anti-reactive oxygen species activity to anti-inflammatory activity of components of eviprostat, a phytotherapeutic agent for benign prostatic hyperplasia. Phytomedicine 2007, 14, 465–472. [Google Scholar]
- Do Monte, F.H.; dos Santos, J.G., Jr; Russi, M.; Lanziotti, V.M.; Leal, L.K.; Cunha, G.M. Antinociceptive and anti-inflammatory properties of the hydroalcoholic extract of stems from Equisetum arvense L. in mice. Pharmacol. Res 2004, 49, 239–243. [Google Scholar]
- Tasken, K.; Stokka, A.J. The molecular machinery for cAMP-dependent immunomodulation in T-cells. Biochem. Soc. Trans 2006, 34, 476–479. [Google Scholar]
- Andersson, K.E.; Arner, A. Urinary bladder contraction and relaxation: Physiology and pathophysiology. Physiol. Rev 2004, 84, 935–986. [Google Scholar]
- Misra, U.K.; Pizzo, S.V. Coordinate regulation of forskolin-induced cellular proliferation in macrophages by protein kinase A/cAMP-response element-binding protein (CREB) and Epac1-Rap1 signaling: Effects of silencing CREB gene expression on Akt activation. J. Biol. Chem 2005, 280, 38276–38289. [Google Scholar]
- Ritchie, R.H.; Rosenkranz, A.C.; Huynh, L.P.; Stephenson, T.; Kaye, D.M.; Dusting, G.J. Activation of IP prostanoid receptors prevents cardiomyocyte hypertrophy via cAMP-dependent signaling. Am. J. Physiol. Heart Circ. Physiol 2004, 287, H1179–H1185. [Google Scholar]
- Lee, S.R.; Hong, C.H.; Choi, Y.D.; Kim, J.H. Increased urinary nerve growth factor as a predictor of persistent detrusor overactivity after bladder outlet obstruction relief in a rat model. J. Urol 2010, 183, 2440–2444. [Google Scholar]
- Monga, M.; Gabal-Shehab, L.L.; Stein, P. Urinary transforming growth factor-beta1 levels correlate with bladder outlet obstruction. Int. J. Urol 2001, 8, 487–489. [Google Scholar]
- Li, K.; Yao, J.; Sawada, N.; Kitamura, M.; Andersson, K.E.; Takeda, M. β-Catenin signaling contributes to platelet derived growth factor elicited bladder smooth muscle cell contraction through up-regulation of Cx43 expression. J. Urol 2012, 188, 307–315. [Google Scholar]
- Yao, J.; Kitamura, M.; Zhu, Y.; Meng, Y.; Kasai, A.; Hiramatsu, N.; Morioka, T.; Takeda, M.; Oite, T. Synergistic effects of PDGF-BB and cAMP-elevating agents on expression of connexin43 in mesangial cells. Am. J. Physiol. Renal. Physiol 2006, 290, F1083–F1093. [Google Scholar]
- Adam, R.M.; Roth, J.A.; Cheng, H.L.; Rice, D.C.; Khoury, J.; Bauer, S.B.; Peters, C.A.; Freeman, M.R. Signaling through PI3K/Akt mediates stretch and PDGF-BB-dependent DNA synthesis in bladder smooth muscle cells. J. Urol 2003, 169, 2388–2393. [Google Scholar]
- Tagaya, M.; Oka, M.; Ueda, M.; Takagaki, K.; Tanaka, M.; Ohgi, T.; Yano, J. Eviprostat suppresses proinflammatory gene expression in the prostate of rats with partial bladder-outlet obstruction: A genome-wide DNA microarray analysis. Cytokine 2009, 47, 185–193. [Google Scholar]
- Yao, J.; Hiramatsu, N.; Zhu, Y.; Morioka, T.; Takeda, M.; Oite, T.; Kitamura, M. Nitric oxide-mediated regulation of connexin43 expression and gap junctional intercellular communication in mesangial cells. J. Am. Soc. Nephrol 2005, 16, 58–67. [Google Scholar]
- Yao, J.; Zhu, Y.; Sun, W.; Sawada, N.; Hiramatsu, N.; Takeda, M.; Kitamura, M. Irsogladine maleate potentiates the effects of nitric oxide on activation of cAMP signalling pathways and suppression of mesangial cell mitogenesis. Br. J. Pharmacol 2007, 151, 457–466. [Google Scholar]
- Zhu, Y.; Yao, J.; Meng, Y.; Kasai, A.; Hiramatsu, N.; Hayakawa, K.; Miida, T.; Takeda, M.; Okada, M.; Kitamura, M. Profiling of functional phosphodiesterase in mesangial cells using a CRE-SEAP-based reporting system. Br. J. Pharmacol 2006, 148, 833–844. [Google Scholar]
- Butt, E.; Abel, K.; Krieger, M.; Palm, D.; Hoppe, V.; Hoppe, J.; Walter, U. cAMP- and cGMP-dependent protein kinase phosphorylation sites of the focal adhesion vasodilator-stimulated phosphoprotein (VASP) in vitro and in intact human platelets. J. Biol. Chem 1994, 269, 14509–14517. [Google Scholar]
- Saez, J.C.; Berthoud, V.M.; Branes, M.C.; Martinez, A.D.; Beyer, E.C. Plasma membrane channels formed by connexins: Their regulation and functions. Physiol. Rev 2003, 83, 1359–1400. [Google Scholar]
- Li, K.; Yao, J.; Shi, L.; Sawada, N.; Chi, Y.; Yan, Q.; Matsue, H.; Kitamura, M.; Takeda, M. Reciprocal regulation between proinflammatory cytokine-induced inducible NO synthase (iNOS) and connexin43 in bladder smooth muscle cells. J. Biol. Chem 2011, 286, 41552–41562. [Google Scholar]
- Clark, J.G.; Kostal, K.M.; Marino, B.A. Modulation of collagen production following bleomycin-induced pulmonary fibrosis in hamsters. Presence of a factor in lung that increases fibroblast prostaglandin E2 and cAMP and suppresses fibroblast proliferation and collagen production. J. Biol. Chem 1982, 257, 8098–8105. [Google Scholar]
- Kitamura, M. The antioxidant N-acetylcysteine induces mesangial cells to create three-dimensional cytoarchitecture that underlies cellular differentiation. J. Am. Soc. Nephrol 1999, 10, 746–751. [Google Scholar]
- Das, R.; Esposito, V.; Abu-Abed, M.; Anand, G.S.; Taylor, S.S.; Melacini, G. cAMP activation of PKA defines an ancient signaling mechanism. Proc. Natl. Acad. Sci. USA 2007, 104, 93–98. [Google Scholar]
- Kamenetsky, M.; Middelhaufe, S.; Bank, E.M.; Levin, L.R.; Buck, J.; Steegborn, C. Molecular details of cAMP generation in mammalian cells: A tale of two systems. J. Mol. Biol 2006, 362, 623–639. [Google Scholar]
- Stehr, M.; Adam, R.M.; Khoury, J.; Zhuang, L.; Solomon, K.R.; Peters, C.A.; Freeman, M.R. Platelet derived growth factor-BB is a potent mitogen for rat ureteral and human bladder smooth muscle cells: Dependence on lipid rafts for cell signaling. J. Urol 2003, 169, 1165–1170. [Google Scholar]
- Sterzel, R.B.; Lovett, D.H.; Foellmer, H.G.; Perfetto, M.; Biemesderfer, D.; Kashgarian, M. Mesangial cell hillocks. Nodular foci of exaggerated growth of cells and matrix in prolonged culture. Am. J. Pathol 1986, 125, 130–140. [Google Scholar]
- Wall, E.A.; Zavzavadjian, J.R.; Chang, M.S.; Randhawa, B.; Zhu, X.; Hsueh, R.C.; Liu, J.; Driver, A.; Bao, X.R.; Sternweis, P.C.; et al. Suppression of LPS-induced TNF-alpha production in macrophages by cAMP is mediated by PKA-AKAP95-p105. Sci. Signal 2009, 2, ra28. [Google Scholar]
- Sawada, N.; Yao, J.; Hiramatsu, N.; Hayakawa, K.; Araki, I.; Takeda, M.; Kitamura, M. Involvement of hypoxia-triggered endoplasmic reticulum stress in outlet obstruction-induced apoptosis in the urinary bladder. Lab. Invest 2008, 88, 553–563. [Google Scholar]
- Fang, X.; Huang, T.; Zhu, Y.; Yan, Q.; Chi, Y.; Jiang, J.X.; Wang, P.; Matsue, H.; Kitamura, M.; Yao, J. Connexin43 hemichannels contribute to cadmium-induced oxidative stress and cell injury. Antioxid. Redox. Signal 2011, 14, 2427–2439. [Google Scholar]

© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, K.; Yao, J.; Chi, Y.; Sawada, N.; Araki, I.; Kitamura, M.; Takeda, M. Eviprostat Activates cAMP Signaling Pathway and Suppresses Bladder Smooth Muscle Cell Proliferation. Int. J. Mol. Sci. 2013, 14, 12107-12122. https://doi.org/10.3390/ijms140612107
Li K, Yao J, Chi Y, Sawada N, Araki I, Kitamura M, Takeda M. Eviprostat Activates cAMP Signaling Pathway and Suppresses Bladder Smooth Muscle Cell Proliferation. International Journal of Molecular Sciences. 2013; 14(6):12107-12122. https://doi.org/10.3390/ijms140612107
Chicago/Turabian StyleLi, Kai, Jian Yao, Yuan Chi, Norifumi Sawada, Isao Araki, Masanori Kitamura, and Masayuki Takeda. 2013. "Eviprostat Activates cAMP Signaling Pathway and Suppresses Bladder Smooth Muscle Cell Proliferation" International Journal of Molecular Sciences 14, no. 6: 12107-12122. https://doi.org/10.3390/ijms140612107