Biosynthetic Pathway and Health Benefits of Fucoxanthin, an Algae-Specific Xanthophyll in Brown Seaweeds

Abstract
:1. Introduction
2. Presence of Fucoxanthin in Brown Seaweed
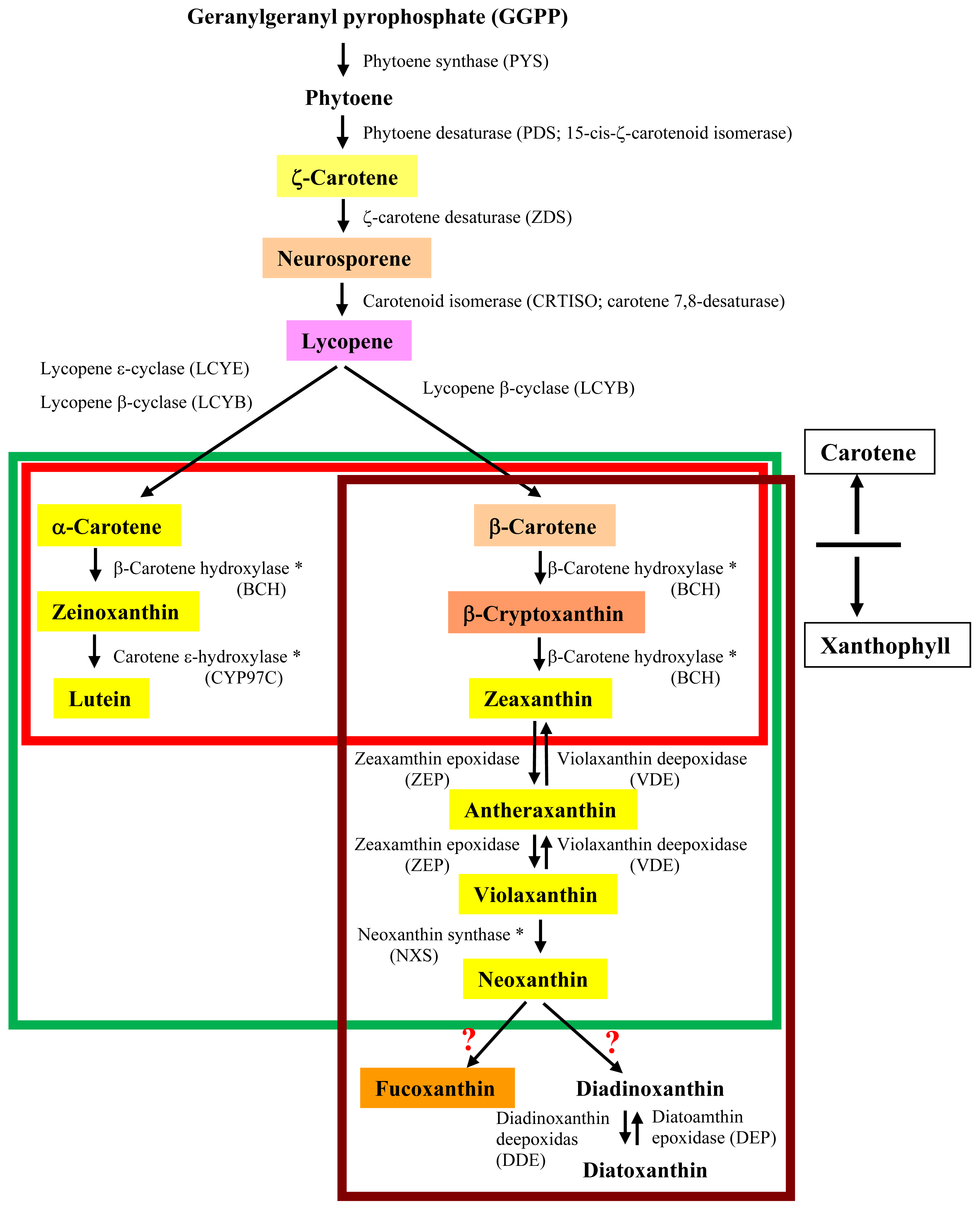
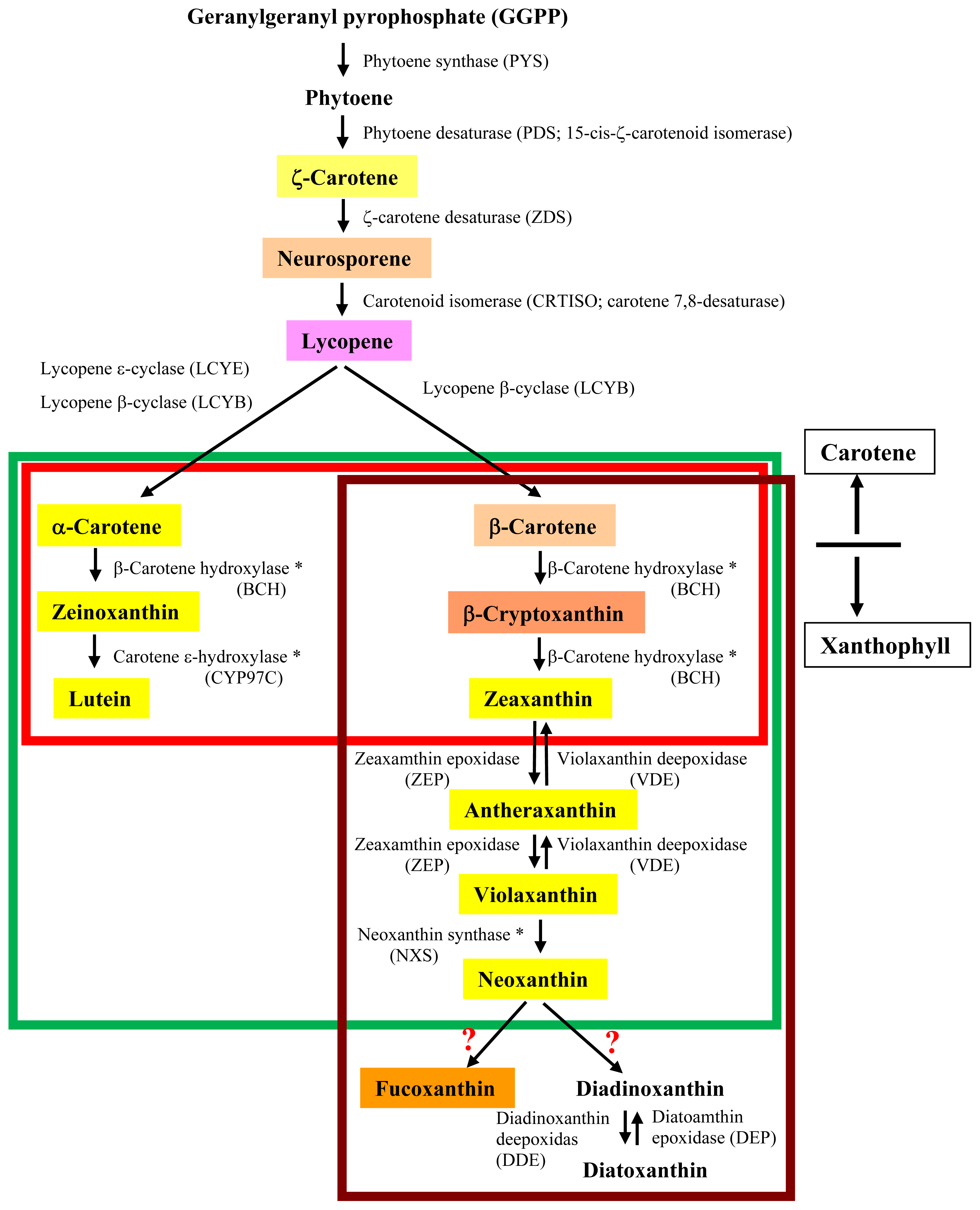
3. Putative Biosynthetic Pathway of Fucoxanthin in Brown Seaweeds
3.1. Proposed Pathways for Fucoxanthin Biosynthesis Based on Knowledge in Diatoms
3.2. Unique Features of Carotenogenic Genes
3.3. The Absence of Genes Encoding β-Carotenoid Hydroxylase and Neoxanthin Synthase
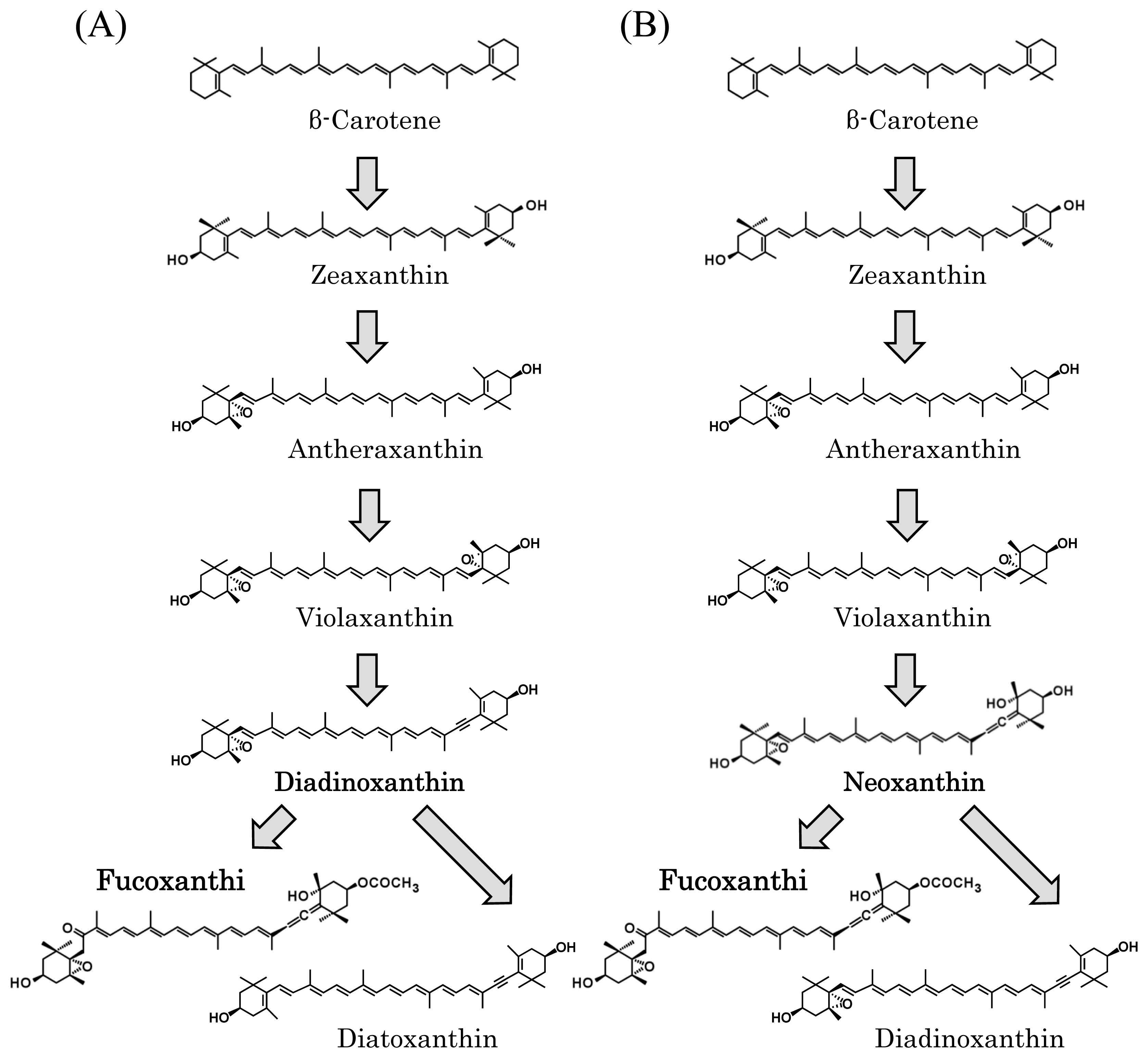
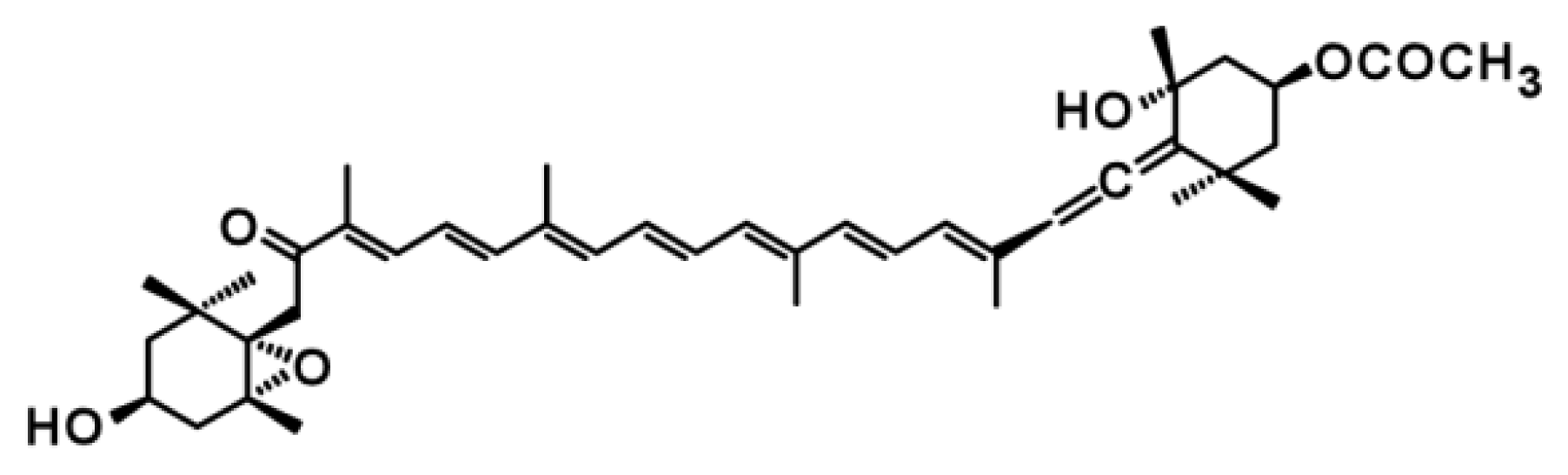
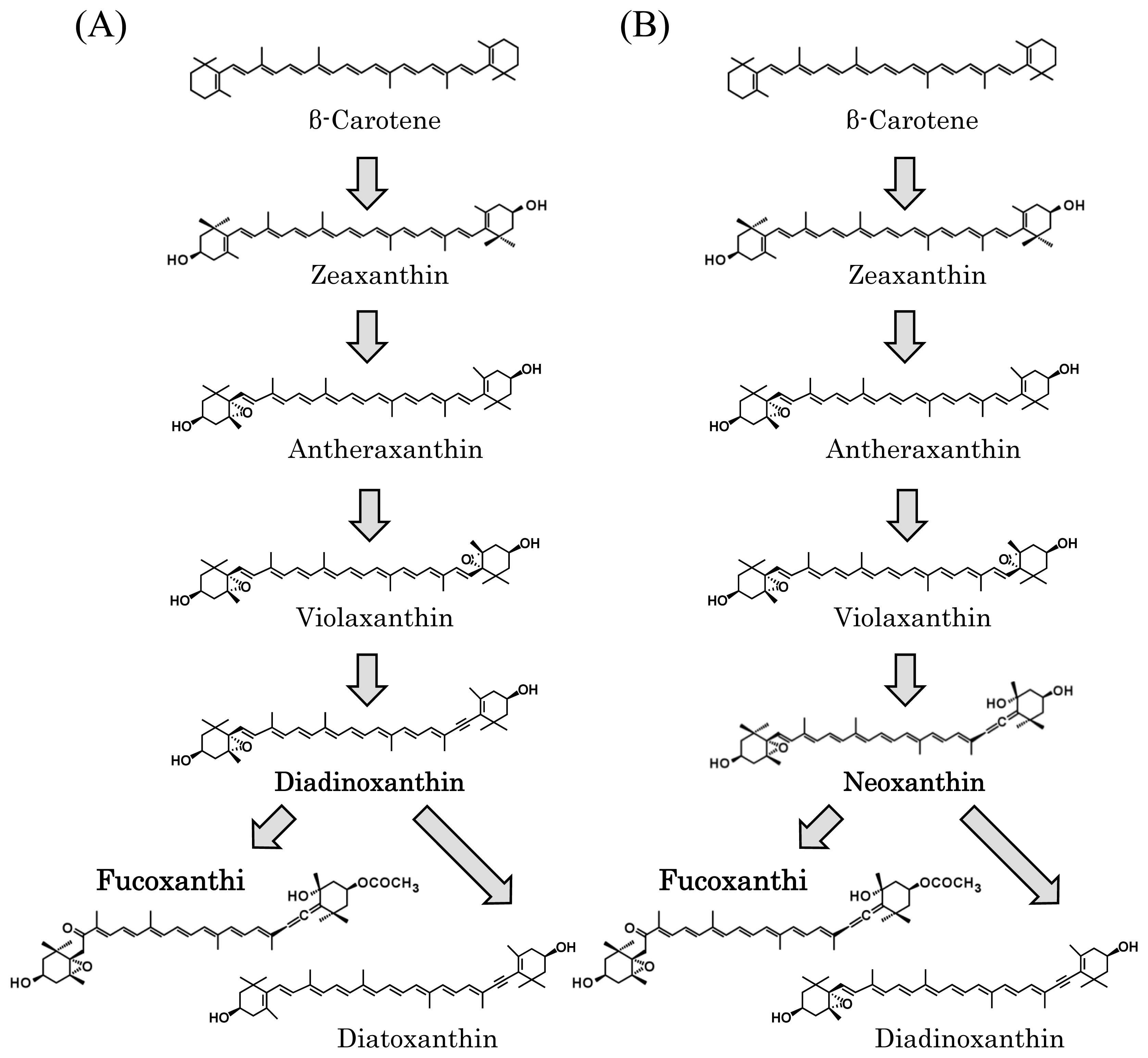
3.4. Unknown Ketolase Involved in Fucoxanthin Biosynthesis
3.5. Toward Resolution of the Fucoxanthin Biosynthetic Pathway in Brown Seaweeds
4. Health Benefits of Fucoxanthin
4.1. Antioxidant Activity
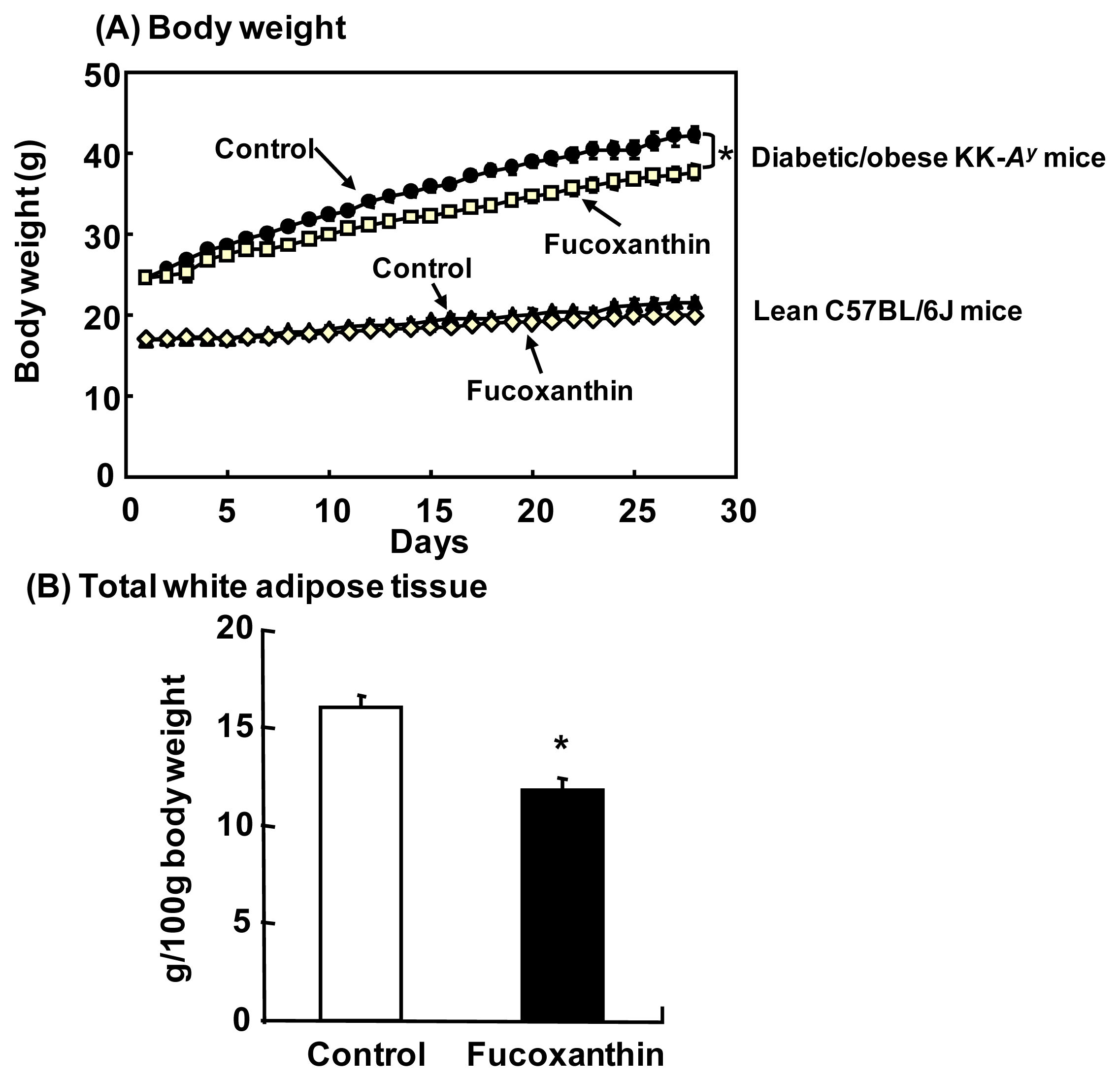
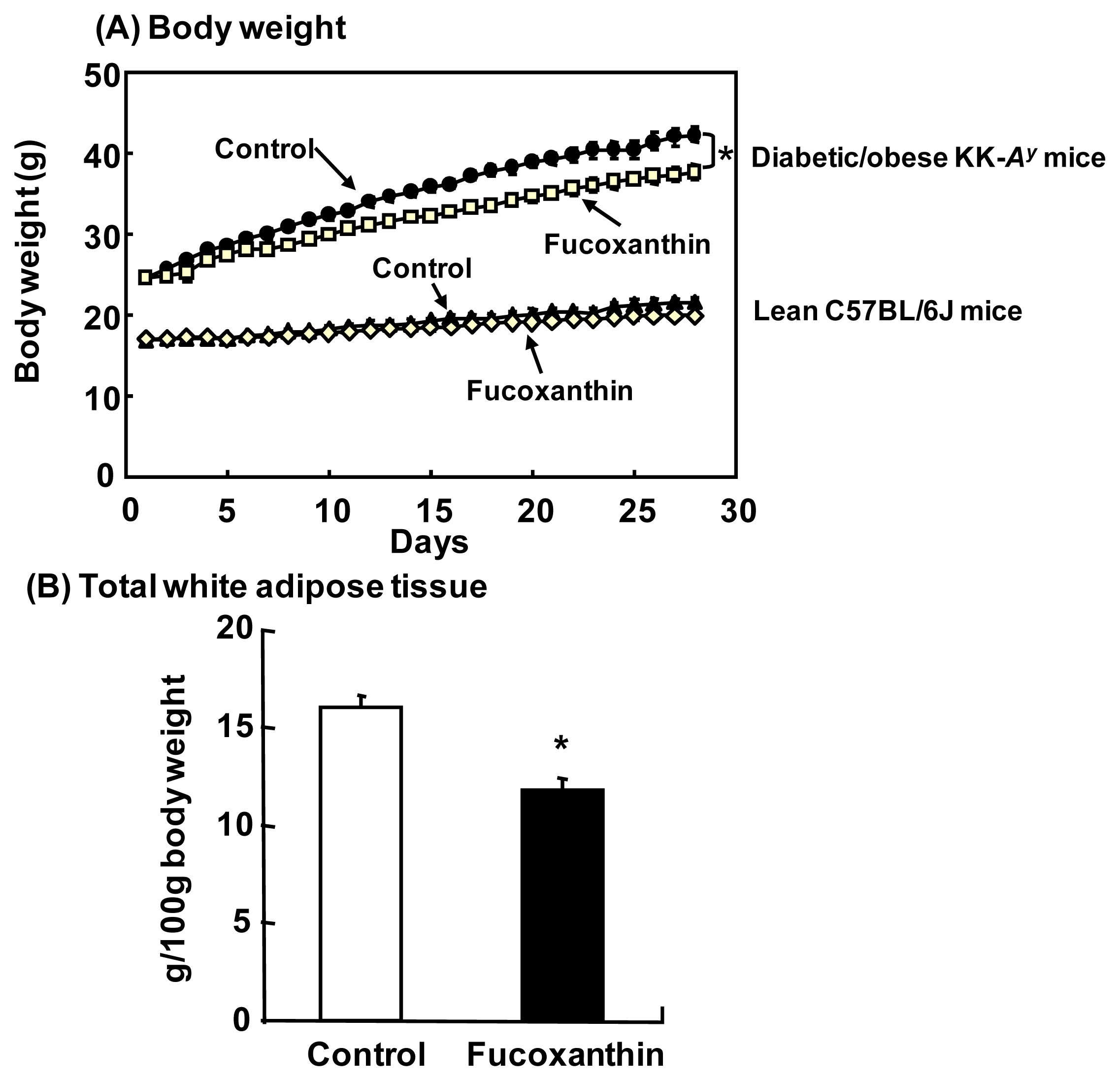
4.2. Anti-Obesity and Anti-Diabetic Effects in Animals
4.3. Anti-Cancer Effects
4.4. Future Perspectives
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Farré, G.; Sanahuja, G.; Naqvi, S.; Bai, C.; Capell, T.; Zhu, C.; Cristou, P. Travel advice on the road to carotenoids in plants. Plant Sci 2010, 179, 28–48. [Google Scholar]
- Takaichi, S. Carotenoids in algae: Distributions, biosynthesis and functions. Mar. Drugs 2011, 9, 1101–1108. [Google Scholar]
- Bartley, G.; Scolinik, P.A. Plant carotenoids: Pigments for photoprotection, visual attraction, and human health. Plant Cell 1995, 7, 1027–1038. [Google Scholar]
- Vishnevetsky, M.; Ovadis, M.; Vainstein, A. Carrotenoid sequestration in plants: The role of carotenoid-associated proteins. Trends Plant Sci 1999, 4, 232–235. [Google Scholar]
- Milborrow, B.V. The pathway of biosynthesis of abscisic acid in vascular plants: A review of the present state of knowledge of ABA biosynthesis. J. Exp. Bot 2001, 52, 1145–1164. [Google Scholar]
- Seo, M.; Koshiba, T. Complex regulation of ABA biosynthesis in plants. Trends Plant Sci 2002, 7, 41–48. [Google Scholar]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta 2012, 1819, 86–96. [Google Scholar]
- Umehara, M.; Hanada, A.; Yoshida, S.; Akiyama, K.; Arite, T.; Takeda-Kamiya, N.; Magome, H.; Kamiya, Y.; Shirasu, K.; Yoneyama, K.; et al. Inhibition of shoot branching by new terpenoid plant hormones. Nature 2008, 455, 195–200. [Google Scholar]
- Gomez-Roldan, V.; Fermas, S.; Brewer, P.B.; Puech-Pagès, V.; Dun, E.A.; Pillot, J.P.; Letisse, F.; Matusova, R.; Danoun, S.; Portais, J.C.; Bouwmeester, H.; Bécard, G.; Beveridge, C.A.; Rameau, C.; Rochange, S.F. Strigolactone inhibition of shoot branching. Nature 2008, 455, 189–194. [Google Scholar]
- Seto, Y.; Kameoka, H.; Yamaguchi, S.; Kyozuka, J. Recent advances in strigolactone research: chemical and biological aspectes. Planr Cell Physiol 2012, 53, 1843–1853. [Google Scholar]
- Nagao, A. Absorption and metabolism of dietary carotenoids. Biofactors 2011, 37, 83–87. [Google Scholar]
- Miyashita, K.; Nishikawa, S.; Beppu, F.; Tsukui, T.; Abe, M.; Hosokawa, M. The allenic carotenoid fucoxanthin, a novel marine nutraceutical from brown seaweeds. J. Sci. Food Agric 2011, 91, 1166–1174. [Google Scholar]
- Peng, J.; Yuan, J.P.; Wu, C.F.; Wang, J.H. Fucoxanthin, a marine carotenoid present in brown seaweeds and diatoms: Metabolism and bioactivities relevant to human health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar]
- Bertrand, M. Carotenoid biosynthesis in diatoms. Photosynth. Res 2010, 106, 89–102. [Google Scholar]
- Coesel, S.; Oborník, M.; Varela, J.; Falciatore, A.; Bowler, C. Evolutionary origins and functions of the carotenoid biosynthesis pathway in marine diatoms. PLoS One 2008, 3, e2896. [Google Scholar]
- Dambek, M.; Eilers, U.; Breitenbach, J.; Steiger, S.; Büchel, C.; Sandmann, G. Biosynthesis of fucoxanthin and diadinoxanthin and function of initial pathway genes in Phaeodactylum tricornutum. J. Exp. Bot 2012, 63, 5607–5612. [Google Scholar]
- Chan, C.X.; Blouin, N.A.; Zhuang, Y.; Zauner, S.; Prochnik, S.E.; Lindquist, E.; Lin, S.; Benning, C.; Lohr, M.; Yarish, C.; et al. Porphyra (Bangiophyceae) transcriptomes insights into red algal development and metabolism. J. Phycol 2012, 48, 1328–1342. [Google Scholar]
- Cunningham, F.X., Jr; Lee, H.; Gantt, E. Carotenod biosynthesis in the primitive red alga Cyanidioschyzon merolae. Eukaryot. Cell 2007, 6, 5330545. [Google Scholar]
- Schubert, N.; García-Mendoza, E.; Pacheco-Ruiz, I. Carotenoid composition of marine red algae. J. Phycol 2006, 42, 1208–1216. [Google Scholar]
- Kakinuma, M.; Shibahara, N.; Ikeda, H.; Maegawa, M.; Amano, H. Thermal stress response of a sterile mutant of Ulva pertusa (Chlorophyta). Fish. Sci 2001, 68, 287–294. [Google Scholar]
- Goss, R.; Jakob, T. Regulation and function of xanthophyll cycle-dependent photoprotection in algae. Photosynth. Res 2010, 106, 103–122. [Google Scholar]
- Lohr, M.; Wilhelm, C. Algae displaying the diadinoxanthin cycle also possess the violaxanthin cycle. Proc. Natl. Acad. Sci. USA 1999, 96, 8784–8789. [Google Scholar]
- Cock, J.M.; Sterck, L.; Rouzé, P.; Scornet, D.; Allen, A.E.; Amoutzias, G.; Anthouard, V.; Artiguenave, F.; Aury, J.M.; Badger, J.H.; et al. The Ectocarpus genome and the independent evolution of multicellularity in brown algae. Nature 2010, 465, 617–621. [Google Scholar]
- Armbrust, E.V.; Berges, J.A.; Bowler, C.; Green, B.R.; Martinez, D.; Putnam, N.H.; Zhou, S.; Allen, A.E.; Apt, K.E.; Bechner, M.; et al. The genome of the diatom Thalassiosira pseudonana: ecology, evolution, and metabolism. Science 2004, 306, 79–86. [Google Scholar]
- Bowler, C.; Allen, A.E.; Badger, J.H.; Grimwood, J.; Jabbari, K.; Kuo, A.; Maheswari, U.; Martens, C.; Maumus, F.; Otillar, R.P.; et al. The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 2008, 456, 239–244. [Google Scholar]
- North, H.M.; de Almeida, A.; Boutin, J.-P.; Frey, A.; To, A.; Botran, L.; Sotta, B.; Marion-Poll, A. The Arabidopsis ABA-deficient mutant aba4 demonstrates that the major route for stress-induced ABA accumulation is via neoxanthin isomers. Plant J 2007, 50, 810–824. [Google Scholar]
- Zhong, Y.J.; Huang, J.C.; Liu, J.; Li, Y.; Jiang, Y.; Xu, Z.F.; Sandmann, G.; Chen, F. Functional characterization of various algal carotenoid ketolases reveals that ketolating zeaxanthin efficiently is essential for high production of astaxanthin in transgenic Arabidopsis. J. Exp. Bot 2011, 62, 3659–3669. [Google Scholar]
- Huang, J.; Zhong, Y.; Sandmann, G.; Liu, J.; Chen, F. Cloning and selection of carotenoid ketolase genes for the engineering of high-yield astaxanthin in plants. Planta 2012, 236, 691–699. [Google Scholar]
- Lohr, M.; Im, C.S.; Grossman, A.R. Genome-based examination of chlorophyll and carotenoid biosynthesis in Chlamydomonas reinhardtii. Plant Physiol 2005, 138, 490–515. [Google Scholar]
- Tran, P.T.; Sharifi, M.N.; Poddar, S.; Dent, R.M.; Niyogi, K.K. Intragenic enhancers and suppressors of phytoene desaturase mutations in Chlamydomonas reinhardtii. PLoS One 2012, 7, e42196. [Google Scholar]
- Biomarkers for Antioxidant Defense and Oxidative Damage: Principle and Practical Applications; Aldini, G.; Yeum, K.J.; Niki, E.; Russell, R.M. (Eds.) Wiley-Blackwell Publishing: Ames, IA, USA; p. 2010.
- Stahl, W.; Sies, H. Photoprotection by dietary carotenoids: Concept, mechanism, evidence and future development. Mol. Nutr. Food Res 2012, 56, 287–295. [Google Scholar]
- Yan, X.; Chuda, Y.; Suzuki, M.; Nagata, T. Fucoxanthin as the major antioxidant in Hijikia fusiformis, a common edible seaweed. Biosci. Biotechnol. Biochem 1999, 63, 605–607. [Google Scholar]
- Sachindra, N.M.; Sato, E.; Maeda, H.; Hosokawa, M.; Niwano, Y.; Kohno, M.; Miyashita, K. Radical scavenging and singlet oxygen quenchingactivity of marine carotenoid fucoxanthin and its metabokites. J. Agric. Food Chem 2007, 55, 8516–8522. [Google Scholar]
- Nomura, T.; Kikuchi, M.; Kubodera, A.; Kawakami, Y. Proton-donative antioxidant activity of fucoxanthin with 1,1-diphenyl-2-picrylhydrazyl (DPPH). Biochem. Mol. Biol. Int 1997, 42, 361–370. [Google Scholar]
- Friedman, J.M. Obesity in the new millennium. Nature 2000, 404, 632–634. [Google Scholar]
- Matsuzawa, Y. The metabolic syndrome and adipocytokines. FEBS Lett 2006, 580, 2917–2922. [Google Scholar]
- Hosokawa, M.; Miyashita, T.; Nishikawa, S.; Emi, S.; Tsukui, T.; Beppu, F.; Okada, T.; Miyashita, K. Fucoxanthin regulates adipocytokine mRNA expression in white adipose tissue of diabetic/obese KK-Ay mice. Arch. Biochem. Biophys 2010, 504, 17–25. [Google Scholar]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Funayama, K.; Miyashita, K. Fucoxanthin from edible seaweed, Undaria pinnatifida, shows antiobesity effect through UCP1 expression in white adipose tissues. Biochem. Biophys. Res. Commun 2005, 332, 392–397. [Google Scholar]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev 2004, 84, 277–359. [Google Scholar]
- Pico, C.; Bonet, M.L.; Palou, A. Stimulation of uncoupling protein synthesis in white adipose tissue of mice treated with the beta 3-adrenergic agonist CGP-12177. Cell Mol. Life Sci 1998, 54, 191–195. [Google Scholar]
- Saito, M.; Okamatsu-Ogura, Y.; Matsushita, M.; Watanabe, K.; Yoneshiro, T.; Nio-Kobayashi, J.; Iwanaga, T.; Miyagawa, M.; Kameya, T.; Nakada, K.; et al. High incidence of metabolically active brown adipose tissue in healthy adult humans: Effects of cold exposure and adiposity. Diabetes 2009, 58, 1526–1531. [Google Scholar]
- Petrovic, N.; Walden, T.B.; Shabalina, I.G.; Timmons, J.A.; Cannon, B.; Nedergaard, J. Chronic peroxisome proliferator-activated receptor gamma (PPARgamma) activation of epididymally derived white adipocyte cultures reveals a population of thermogenically competent, UCP1-containing adipocytes molecularly distinct from classic brown adipocytes. J. Biol. Chem 2010, 285, 7153–7164. [Google Scholar]
- Wu, J.; Bostrom, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige adipocytes are a distinct type of thermogenic fat cell inmouse and human. Cell 2012, 150, 366–376. [Google Scholar]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Dietary combination of fucoxanthin and fish oil attenuates the weight gain of white adipose tissue and decreases blood glucose in obese/diabetic KK-Ay mice. J. Agric. Food Chem 2007, 55, 7701–7706. [Google Scholar]
- Woo, M.N.; Jeon, S.M.; Kim, H.J.; Lee, M.K.; Shin, S.K.; Shin, Y.C.; Park, Y.B.; Choi, M.S. Fucoxanthin supplementation improves plasma and hepatic lipid metabolism and blood glucose concentration in high-fat fed C57BL/6N mice. Chem. Biol. Interact 2010, 186, 316–322. [Google Scholar]
- Nishikawa, S.; Hosokawa, M.; Miyashita, K. Fucoxanthin promotes translocation and induction of glucose transporter 4 in skeletal muscles of diabetic/obese KK-Ay mice. Phytomedicine 2012, 19, 389–394. [Google Scholar]
- Abidov, M.; Ramazanov, Z.; Seifulla, R.; Grachev, S. The effect of Xanthigen in the weight management of obese premenopausal women with non-alcoholic fatty liver disease and normal liver fat. Diabetes Obes. Metab 2010, 12, 72–81. [Google Scholar]
- Sporn, M.B.; Suh, N. Chemoprevention of cancer. Carcinogenesis 2000, 21, 525–530. [Google Scholar]
- Levy, J.; Bosin, E.; Feldman, B.; Giat, Y.; Miinster, A.; Danilenko, M.; Sharoni, Y. Lycopene is a more potent inhibitor of human cancer cell proliferation than either α-carotene or β-carotene. Nutr. Cancer 1995, 24, 257–266. [Google Scholar]
- Rock, C.L. Carotenoid update. J. Am. Diet Assoc 2003, 103, 423–425. [Google Scholar]
- Heinonen, O.P.; Albanes, D. The effect of vitamin E and β carotene on the incidence of lung cancer and other cancers in male smokers. The α-Tocopherol, β Carotene Cancer Prevention Study Group. N. Engl. J. Med 1994, 330, 1029–1035. [Google Scholar]
- Hennekens, C.H.; Buring, J.E.; Manson, J.E.; Stampfer, M.; Rosner, B.; Cook, N.R.; Belanger, C.; LaMotte, F.; Gaziano, J.M.; Ridker, P.M.; et al. Lack of effect of long-term supplementation with β carotene on the incidence of malignant neoplasms and cardiovascular disease. N. Engl. J. Med 1996, 334, 1145–1149. [Google Scholar]
- Omenn, G.S.; Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Glass, A.; Keogh, J.P.; Meyskens, F.L.; Valanis, B.; Williams, J.H.; et al. Effects of a combination of β carotene and vitamin A on lung cancer and cardiovascular disease. N. Engl. J. Med 1996, 334, 1150–1155. [Google Scholar]
- Das, S.K.; Hashimoto, T.; Kanazawa, K. Growth inhibition of human hepatic carcinoma HepG2 cells by fucoxanthin is associated with down-regulation of cyclin D. Biochim. Biophys. Acta 2008, 1780, 743–749. [Google Scholar]
- Hosokawa, M.; Kudo, M.; Maeda, H.; Kohno, H.; Tanaka, T.; Miyashita, K. Fucoxanthin induces apoptosis and enhances the antiproliferative effect of the PPARgamma ligand, troglitazone, on colon cancer cells. Biochim. Biophys. Acta 2004, 1675, 113–119. [Google Scholar]
- Asai, A.; Sugawara, T.; Ono, H.; Nagao, A. Biotransformation of fucoxanthinol into amarouciaxanthin A in mice and HepG2 cells: Formation and cytotoxicity of fucoxanthin metabolites. Drug Metab. Dispos 2004, 32, 205–211. [Google Scholar]
- Konishi, I.; Hosokawa, M.; Sashima, T.; Kobayashi, H.; Miyashita, K. Halocynthiaxanthin and fucoxanthinol isolated from Halocynthia roretzi induce apoptosis in human leukemia, breast and colon cancer cells. Comp. Biochem. Physiol. C 2006, 142, 53–59. [Google Scholar]
- Kim, J.M.; Araki, S.; Kim, D.J.; Park, C.B.; Takasuka, N.; Baba-Toriyama, H.; Ota, T.; Nir, Z.; Khachik, F.; Shimidzu, N.; et al. Chemopreventive effects of carotenoids and curcumins on mouse colon carcinogenesis after 1,2-dimethylhydrazine initiation. Carcinogenesis 1998, 19, 81–85. [Google Scholar]
- Wang, J.; Chen, S.; Xu, S.; Yu, X.; Ma, D.; Hu, X.; Cao, X. In vivo induction of apoptosis by fucoxanthin, a marine carotenoid, associated with down-regulating STAT3/EGFR signaling in sarcoma 180 (S180) xenografts-bearing mice. Mar. Drugs 2012, 10, 2055–2068. [Google Scholar]
- Liu, C.L.; Huang, Y.S.; Hosokawa, M.; Miyashita, K.; Hu, M.L. Inhibition of proliferation of a hepatoma cell line by fucoxanthin in relation to cell cycle arrest and enhanced gap junctional intercellular communication. Chem. Biol. Interact 2009, 182, 165–172. [Google Scholar]
- Liu, C.L.; Chiu, Y.T.; Hu, M.L. Fucoxanthin enhances HO-1 and NQO1 expression in murine hepatic BNL CL.2 cells through activation of the Nrf2/ARE system partially by its pro-oxidant activity. J. Agric. Food Chem 2011, 59, 11344–11351. [Google Scholar]
- D’Orazio, N.; Gemello, E.; Gammone, M.A.; de Girolamo, M.; Ficoneri, C.; Riccioni, G. Fucoxantin: A treasure from the sea. Mar Drugs 2012, 10, 604–616. [Google Scholar]
- Beppu, F.; Niwano, Y.; Tsukui, T.; Hosokawa, M.; Miyashita, K. Single and repeated oral dose toxicity study of fucoxanthin (FX), a marine carotenoid, in mice. J. Toxicol. Sci 2009, 34, 501–510. [Google Scholar]
- Beppu, F.; Hosokawa, M.; Niwano, Y.; Miyashita, K. Effects of dietary fucoxanthin on cholesterol metabolism in diabetic/obese KK-Ay mice. Lipids Health Dis 2012, 11, 112. [Google Scholar]
- Hashimoto, T.; Ozaki, Y.; Mizuno, M.; Yoshida, M.; Nishitani, Y.; Azuma, T.; Komoto, A.; Maoka, T.; Tanino, Y.; Kanazawa, K. Pharmacokinetics of fucoxanthinol in human plasma after the oral administration of kombu extract. Br. J. Nutr 2012, 107, 1566–1569. [Google Scholar]
- Okuzumi, J.; Nishino, H.; Murakoshi, M.; Iwashima, A.; Tanaka, Y.; Yamane, T.; Fujita, Y.; Takahashi, T. Inhibitory effects of fucoxanthin, a natural carotenoid, on N-myc expression and cell cycle progression in human malignant tumor cells. Cancer Lett 1990, 55, 75–81. [Google Scholar]
- Hosokawa, M.; Wanezaki, S.; Miyauchi, K.; Kurihara, H.; Kohno, H.; Kawabata, J.; Odashima, S.; Takahashi, K. Apoptosis-inducing effect of fucoxanthin on human leukemia cell line HL-60. Food Sci. Technol. Res 1999, 19, 243–246. [Google Scholar]
- Kim, K.N.; Heo, S.J.; Kang, S.M.; Ahn, G.; Jeon, Y.J. Fucoxanthin induces apoptosis in human leukemia HL-60 cells through a ROS-mediated Bcl-xL pathway. Toxicol. in Vitro 2010, 24, 1648–1654. [Google Scholar]
- Ganesan, P.; Noda, K.; Manabe, Y.; Ohkubo, T.; Tanaka, Y.; Maoka, T.; Sugawara, T.; Hirata, T. Siphonaxanthin, a marine carotenoid from green algae, effectively induces apoptosis in human leukemia (HL-60) cells. Biochim. Biophys. Acta 2011, 1810, 497–503. [Google Scholar]
- Kotake-Nara, E.; Asai, A.; Nagao, A. Neoxanthin and fucoxanthin induce apoptosis in PC-3 human prostate cancer cells. Cancer Lett 2005, 220, 75–84. [Google Scholar]
- Satomi, Y.; Nishino, H. Fucoxanthin, a natural carotenoid, induces G1 arrest and GADD45 gene expression in human cancer cells. In Vivo 2007, 21, 305–309. [Google Scholar]
- Satomi, Y. Fucoxanthin induces GADD45A expression and G1 arrest with SAPK/JNK activation in LNCap human prostate cancer cells. Anticancer Res 2012, 32, 807–813. [Google Scholar]
- Yu, R.X.; Hu, X.M.; Xu, S.Q.; Jiang, Z.J.; Yang, W. Effects of fucoxanthin on proliferation and apoptosis in human gastric adenocarcinoma MGC-803 cells via JAK/STAT signal pathway. Eur. J. Pharmacol 2011, 657, 10–19. [Google Scholar]
- Zhang, Z.Y.; Zhang, P.J.; Hamada, M.; Takahashi, S.; Xing, G.Q.; Liu, J.Q.; Sugiura, N. Potential chemoprevention effect of dietary fucoxanthin on urinary bladder cancer EJ-1 cell line. Oncol. Rep 2008, 20, 1099–1103. [Google Scholar]
- Eid, S.Y.; Ei-Readi, M.Z.; Wink, M. Carotenoids reverse multidrug resistance in cancer cells by interfering with ABC-transporters. Phytomedicine 2012, 19, 977–987. [Google Scholar]
- Ishikawa, C.; Tafuku, S.; Kadekaru, T.; Sawada, S.; Tomita, M.; Okudaira, T.; Nakazato, T.; Toda, T.; Uchihara, J.N.; Taira, N.; et al. Antiadult T-cell leukemia effects of brown algae fucoxanthin and its deacetylated product fucoxanthinol. Int. J. Cancer 2008, 123, 2702–2712. [Google Scholar]
- Yamamoto, K.; Ishikawa, C.; Katano, H.; Yasumoto, T.; Mori, N. Fucoxanthin and its deacetylated product, fucoxanthinol, induce apoptosis of primary effusion lymphomas. Cancer Lett 2011, 300, 225–234. [Google Scholar]
- Nishino, H. Cancer prevention by carotenoids. Mutat. Res 1998, 402, 159–163. [Google Scholar]
- Okuzumi, J.; Takahashi, T.; Yamane, T.; Kitao, Y.; Inagake, M.; Ohya, K.; Tanaka, Y. Inhibitory effects of fucoxanthin, a natural carotenoid, on N-ethyl-N′-nitro-N-nitrosoguanidine-induced mouse duodenal carcinogenesis. Cancer Lett 1993, 68, 159–168. [Google Scholar]
- Shimoda, H.; Tanaka, J.; Shan, S.J.; Maoka, T. Anti-pigmentary activity of fucoxanthin and its influence on skin mRNA expression of melanogenic molecules. J. Pharm. Pharmacol 2010, 62, 1137–1145. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BCH | LTL | ZEP | VDE | VDL | VDR | NXS | |
|---|---|---|---|---|---|---|---|
| Brown algae | |||||||
| E. siliculosus | − | − | + | + | + | + | − |
| T. pseudonana | − | ++ | ++ | + | + | + | − |
| P. tricornutum | − | ++ | +++ | + | ++ | + | − |
| Red algae | |||||||
| C. merolae | + (Chl) | − | − | − | − | − | − |
| P. umbilicalis | − | − | + | − | − | − | − |
| P. purpurea | − | − | + | − | − | − | − |
| Green alga | |||||||
| C. reinhardtii | + (Partial) | − | + | − | − | + | − |
| Terrestrial plant | |||||||
| A. thaliana | − | +++++ | + | + | − | + | + |
| Carotenoid | Type of cancer | Mechanism | Target molecules | References |
|---|---|---|---|---|
| In vitro | ||||
| Fucoxanthin | GOTO (neuroblastoma) | G1 Cell cycle arrest | N-myc | [67] |
| HL-60 (leukemia) | Apoptosis induction | Caspase-3, 7, 9 | [68–70] | |
| Caco-2, HT29, DLD-1 (colon cancer) | Apoptosis induction | Bcl-2 | [56] | |
| PC-3, DU-145, LNCap (prostate cancer) | Apoptosis induction | Bcl-2, Bax, Caspase-3 | [71] | |
| DU-145, LNCap (prostate cancer) | G1 cell cycle arrest | GADD45A, SAPK/JNK | [72,73] | |
| HepG2 (hepato carcinoma) | G1 cell cycle arrest | Cyclin D | [55] | |
| SK-Hep-1 (hepato carcinoma) | G1 cell cycle arrest, apoptosis induction | Connexin 43, Connexin-32 | [61] | |
| MGC-803 (gastric adenocarcinoma) | G2/M cell cycle arrest, apoptosis induction | Cyclin B1, Survivin | [74] | |
| EJ-1 (urinary bladder cancer) | Apoptosis induction | Caspase-3 | [75] | |
| Caco-2 cell (colon cancer) | Enhancement on cytotoxicity of agents | MDR1 | [76] | |
| Fucoxanthinol | HL-60 (leukemia), MCF-7 (breast cancer), Caco-2 (colon cancer) | Apoptosis induction | Bcl-2 | [58] |
| PC-3 (prostate cancer) | Antiproliferative effect | [57] | ||
| T cell leukemia | Antiproliferative effect | [77] | ||
| BCBL-1, TY-1 (lymphoma) | G1 cell cycle arrest, apoptosis induction | NF-kB, AP-1, PI3kinase/Akt | [78] | |
| In vivo | ||||
| Fucoxanthin | Colon cancer | [69] | ||
| Liver tumorigenesis | [79] | |||
| Duodenal carcinogenesis | [80] | |||
| Sarcoma | Apoptosis induction | STAT3/EGFR | [60] | |
| Melanoma | Anti-melanogenesis | COX-2, p75NTR, EP1, MC1R | [81] | |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mikami, K.; Hosokawa, M. Biosynthetic Pathway and Health Benefits of Fucoxanthin, an Algae-Specific Xanthophyll in Brown Seaweeds. Int. J. Mol. Sci. 2013, 14, 13763-13781. https://doi.org/10.3390/ijms140713763
Mikami K, Hosokawa M. Biosynthetic Pathway and Health Benefits of Fucoxanthin, an Algae-Specific Xanthophyll in Brown Seaweeds. International Journal of Molecular Sciences. 2013; 14(7):13763-13781. https://doi.org/10.3390/ijms140713763
Chicago/Turabian StyleMikami, Koji, and Masashi Hosokawa. 2013. "Biosynthetic Pathway and Health Benefits of Fucoxanthin, an Algae-Specific Xanthophyll in Brown Seaweeds" International Journal of Molecular Sciences 14, no. 7: 13763-13781. https://doi.org/10.3390/ijms140713763