Lipidosterolic Extract of Serenoa Repens Modulates the Expression of Inflammation Related-Genes in Benign Prostatic Hyperplasia Epithelial and Stromal Cells


Abstract
:1. Introduction
2. Results
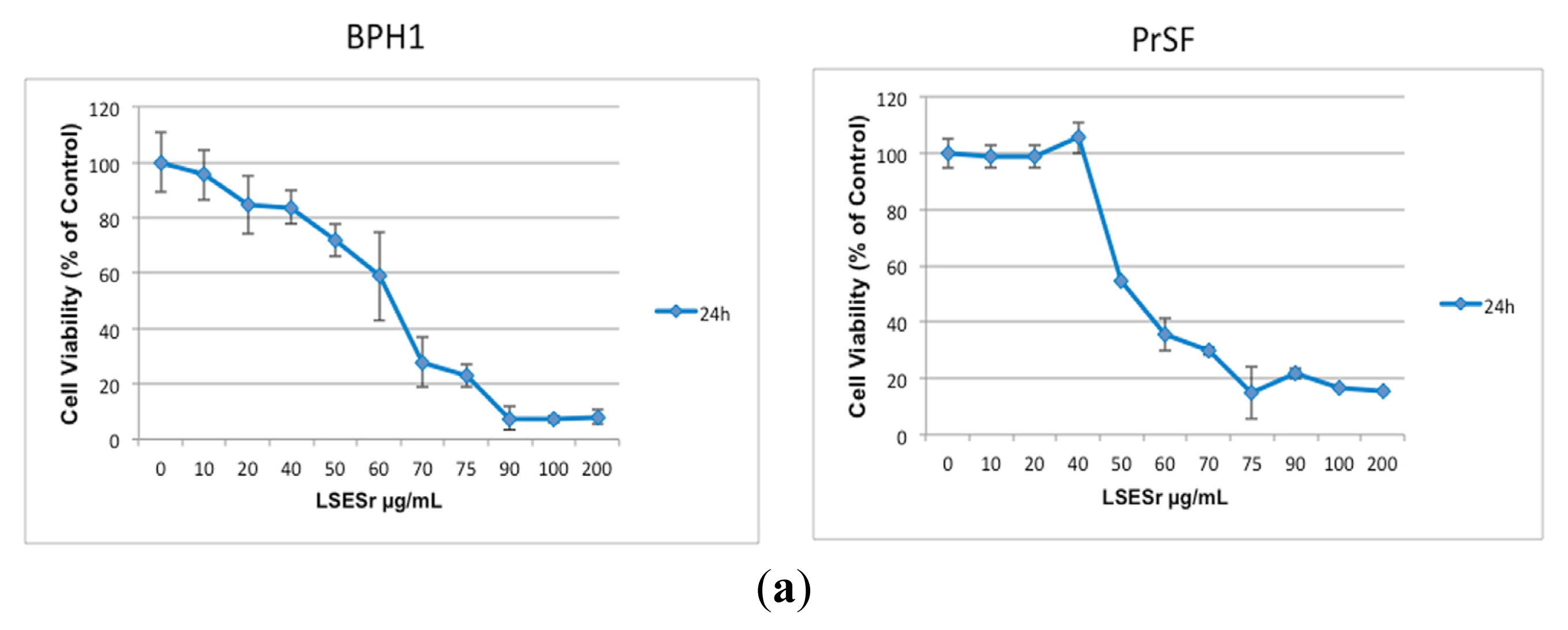
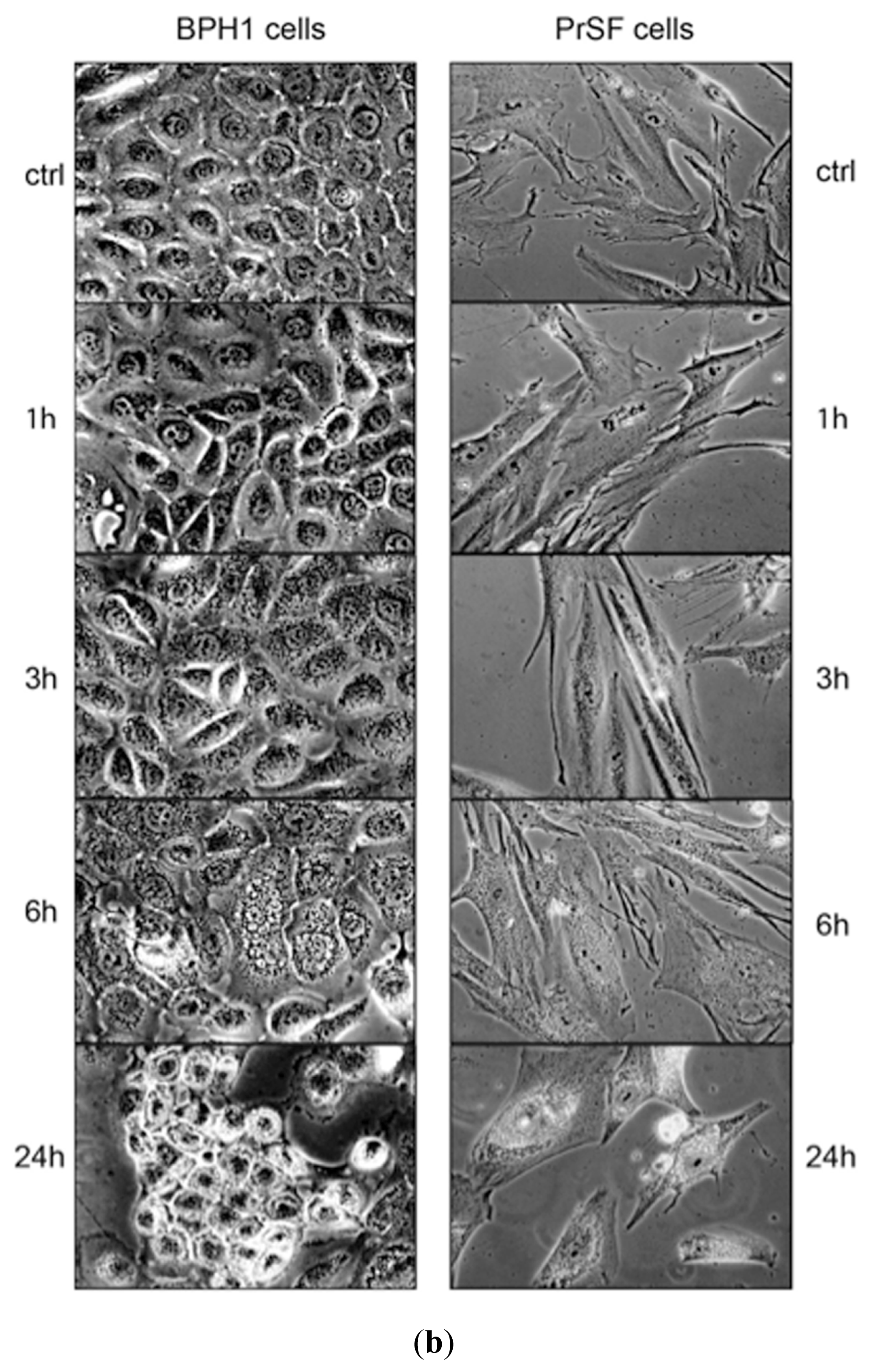
2.1. LSESr Affects Cell Viability in Cultured Human Epithelial and Stromal Cells Derived from Benign Prostatic Hyperplasia (BPH)
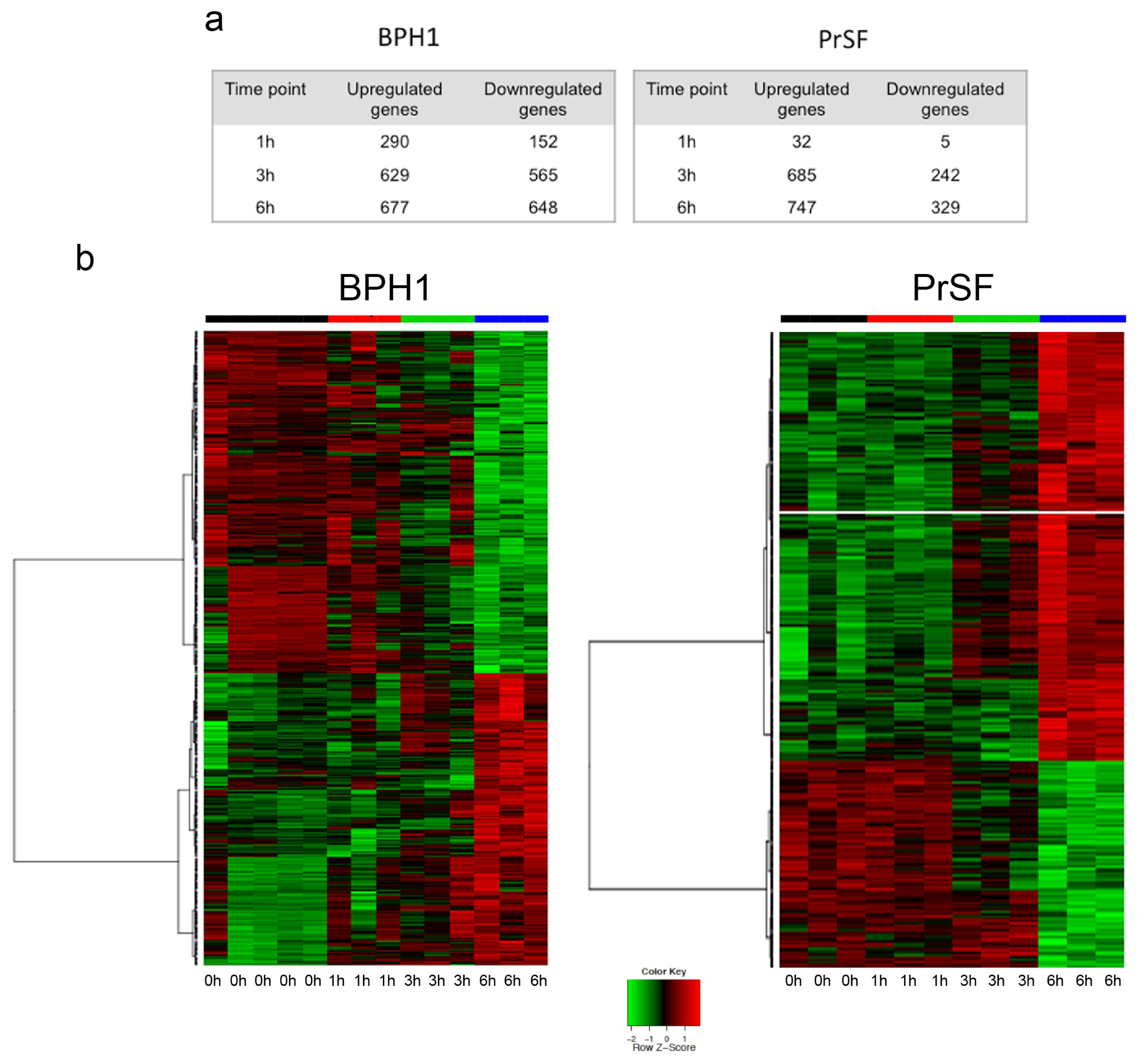
2.2. Gene Expression Profiling of BPH Epithelial and Stromal Cells Treated with LSESr
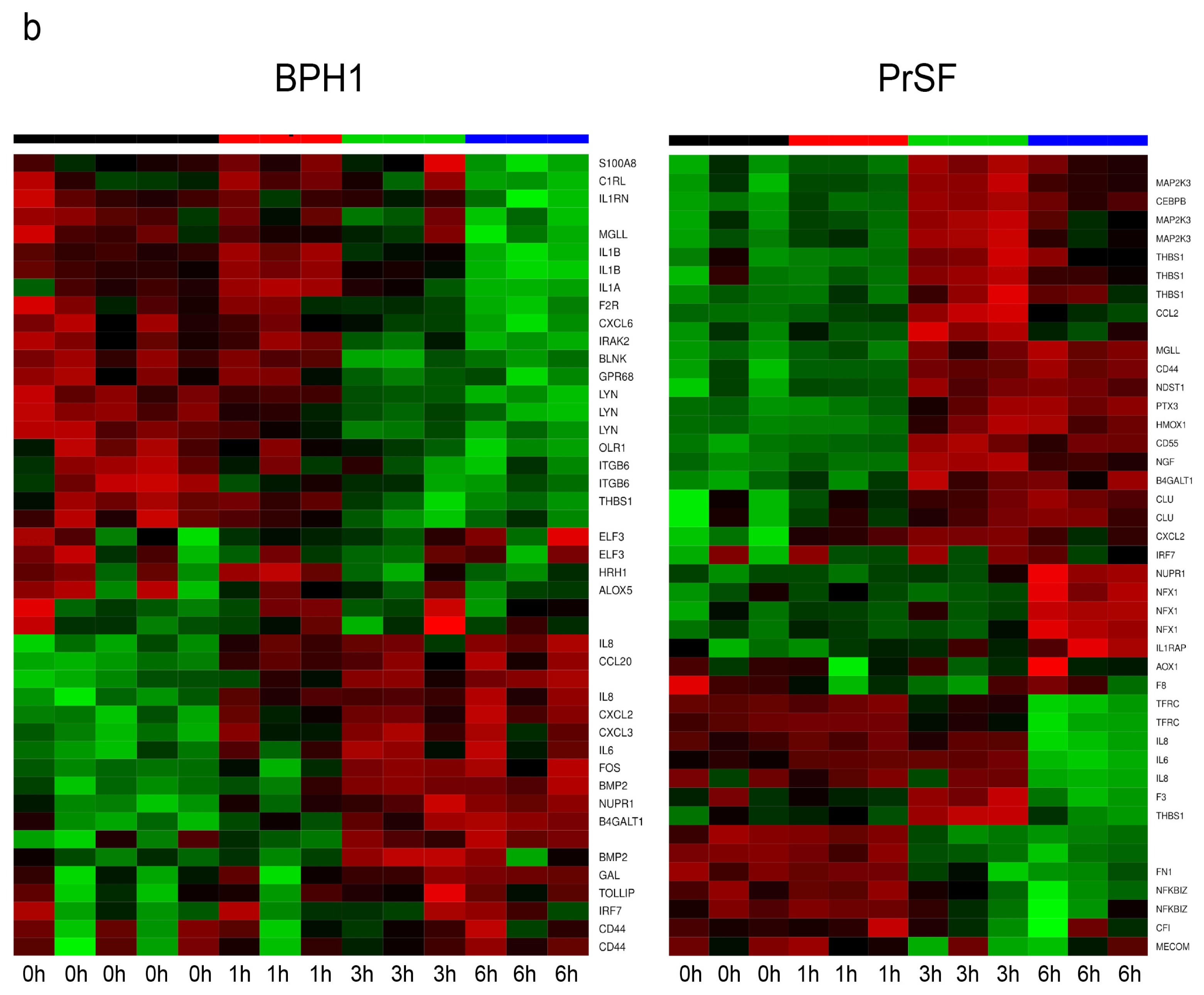
2.2.1. LSESr Induces Changes in Gene Expression Pattern in BPH Epithelial and Stromal Cells
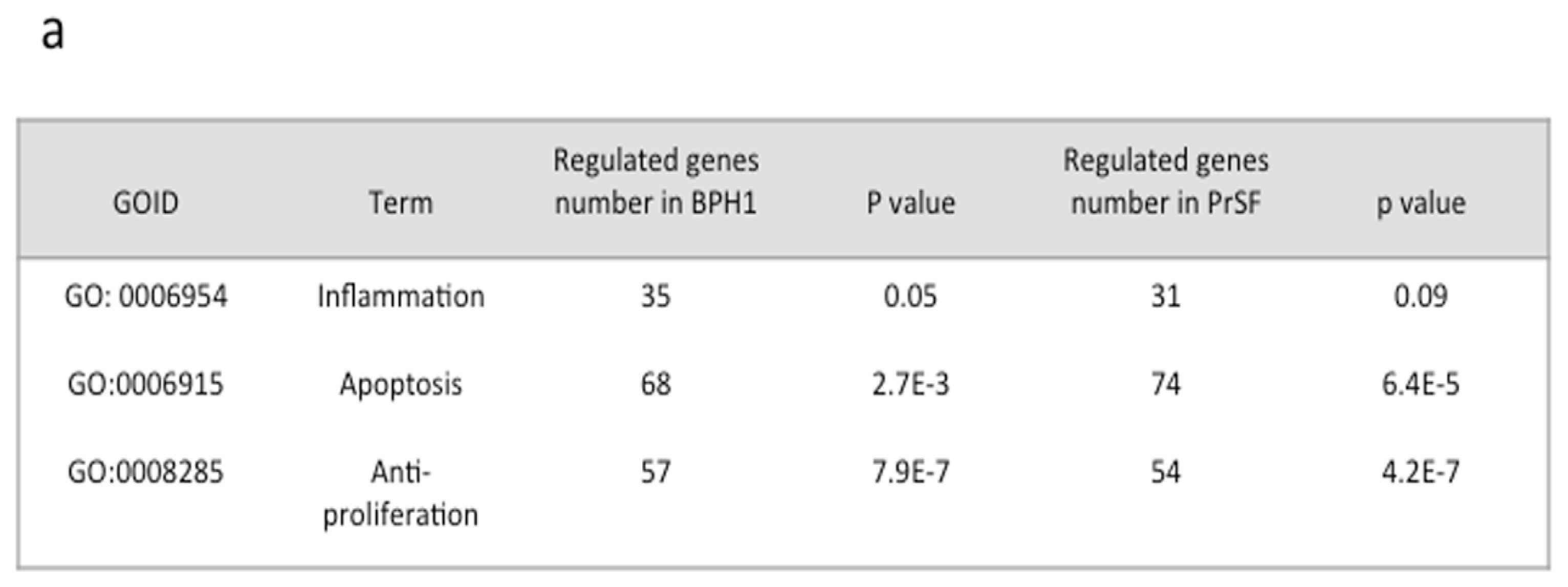
2.2.2. Changes in Apoptosis, Proliferation, and Inflammation Related Processes Revealed by Gene Ontology (GO) Analysis
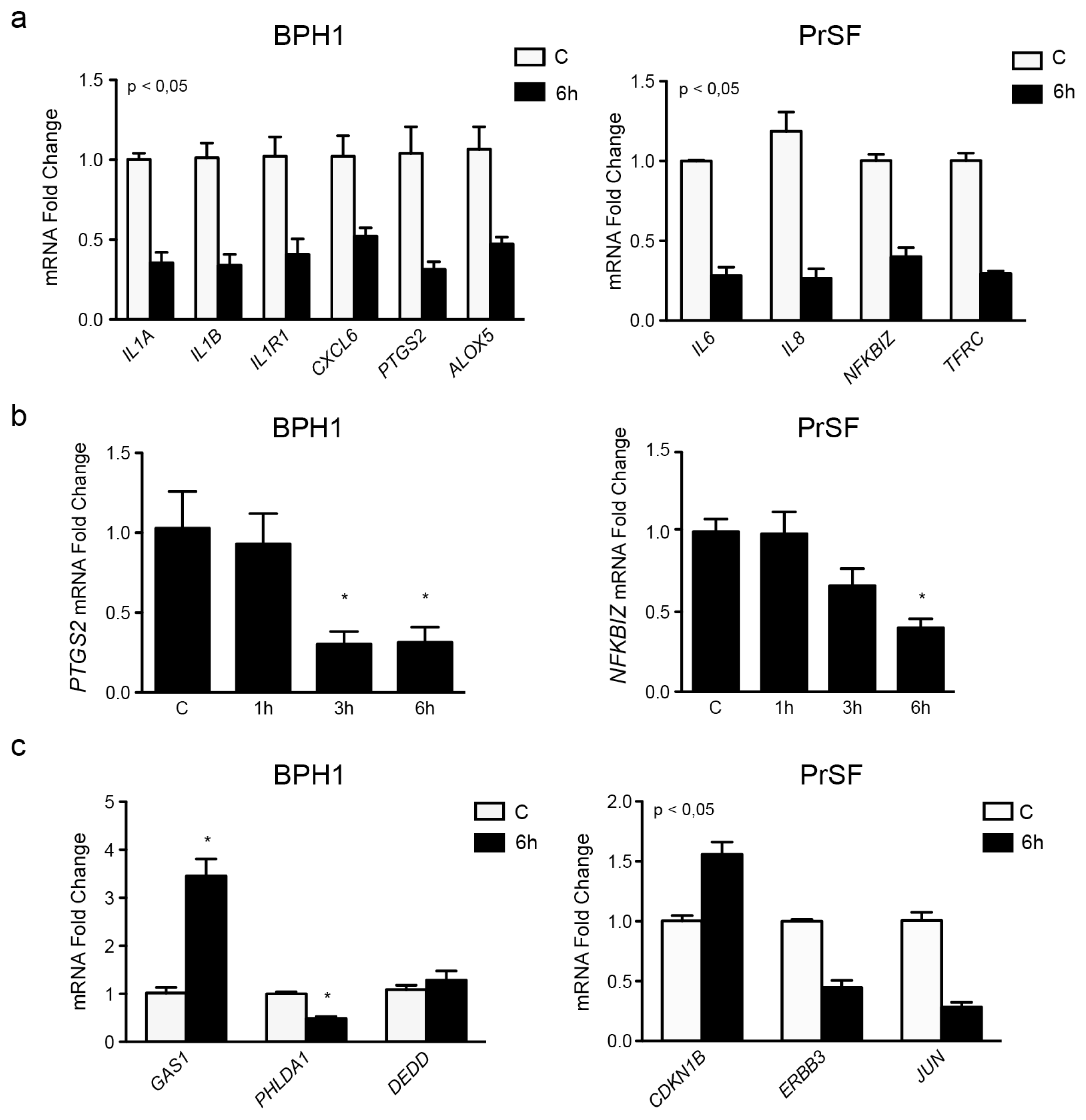
2.2.3. LSESr Perturbs Expression of Inflammation-Related Genes in BPH Epithelial and Stromal Cells
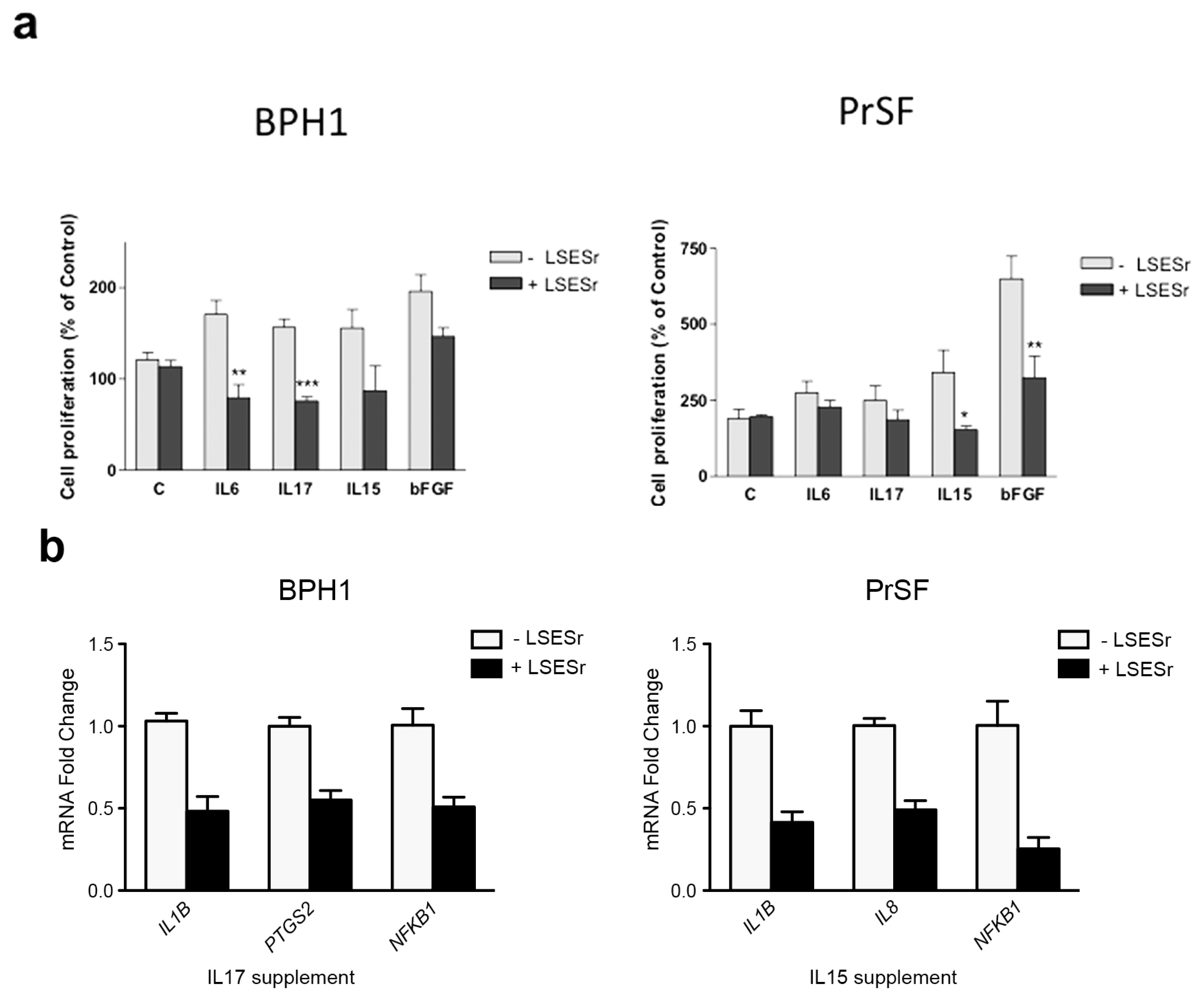
2.3. LSESr Impedes the Effects of Pro-Inflammatory Factors on Proliferation in BPH Epithelial and Stromal Cells
2.4. Discussion
3. Experimental Section
3.1. Cell Culture
3.2. Lipidosterolic Extract of Serenoa repens
3.3. Cell Viability Assay
3.4. Cell Proliferation Assay
3.5. Microarray Hybridization and Analysis
3.6. Quantitative Real-Time PCR (RT-PCR)
4. Conclusions
Supplementary Information
ijms-14-14301-s001.pdfAcknowledgments
Conflict of Interest
References
- Carter, H.B.; Coffey, D.S. The prostate: An increasing medical problem. Prostate 1990, 16, 39–48. [Google Scholar]
- Lee, K.L.; Peehl, D.M. Molecular and cellular pathogenesis of benign prostatic hyperplasia. J. Urol 2004, 172, 1784–1791. [Google Scholar]
- Kramer, G.; Mitteregger, D.; Marberger, M. Is benign prostatic hyperplasia (BPH) an immune inflammatory disease? Eur. Urol 2007, 51, 1202–1216. [Google Scholar]
- Robert, G.; Descazeaud, A.; Nicolaiew, N.; Terry, S.; Sirab, N.; Vacherot, F.; Maille, P.; Allory, Y.; de la Taille, A. Inflammation in benign prostatic hyperplasia: A 282 patients’ immunohistochemical analysis. Prostate 2009, 69, 1774–1780. [Google Scholar]
- Schenk, J.M.; Kristal, A.R.; Neuhouser, M.L.; Tangen, C.M.; White, E.; Lin, D.W.; Thompson, I.M. Serum adiponectin, C-peptide and leptin and risk of symptomatic benign prostatic hyperplasia: Results from the prostate cancer prevention trial. Prostate 2009, 69, 1303–1311. [Google Scholar]
- Hubmer, G. Inflammation of the hypertrophic prostate. Zeitschrift fur Urologie und Nephrologie 1968, 61, 801–804. [Google Scholar]
- Roehrborn, C.G. Combination medical therapy for lower urinary tract symptoms and benign prostatic hyperplasia. Rev. Urol 2005, 7, S43–S51. [Google Scholar]
- Nickel, J.C.; Roehrborn, C.G.; O’Leary, M.P.; Bostwick, D.G.; Somerville, M.C.; Rittmaster, R.S. The relationship between prostate inflammation and lower urinary tract symptoms: examination of baseline data from the REDUCE trial. European urology 2008, 54, 1379–1384. [Google Scholar]
- Minnery, C.H.; Getzenberg, R.H. Benign prostatic hyperplasia cell line viability and modulation of jm-27 by doxazosin and Ibuprofen. J. Urol 2005, 174, 375–379. [Google Scholar]
- Falahatkar, S.; Mokhtari, G.; Pourreza, F.; Asgari, S.A.; Kamran, A.N. Celecoxib for treatment of nocturia caused by benign prostatic hyperplasia: A prospective, randomized, double-blind, placebo-controlled study. Urology 2008, 72, 813–816. [Google Scholar]
- Raynaud, J.P.; Cousse, H.; Martin, P.M. Inhibition of type 1 and type 2 5alpha-reductase activity by free fatty acids, active ingredients of Permixon. J. Steroid Biochem. Mol. Biol 2002, 82, 233–239. [Google Scholar]
- Bayne, C.W.; Donnelly, F.; Ross, M.; Habib, F.K. Serenoa repens (Permixon): A 5alpha-reductase types I and II inhibitor-new evidence in a coculture model of BPH. Prostate 1999, 40, 232–241. [Google Scholar]
- Di Silverio, F.; Monti, S.; Sciarra, A.; Varasano, P.A.; Martini, C.; Lanzara, S.; D’Eramo, G.; di Nicola, S.; Toscano, V. Effects of long-term treatment with Serenoa repens (Permixon) on the concentrations and regional distribution of androgens and epidermal growth factor in benign prostatic hyperplasia. Prostate 1998, 37, 77–83. [Google Scholar]
- Di Silverio, F.; D’Eramo, G.; Lubrano, C.; Flammia, G.P.; Sciarra, A.; Palma, E.; Caponera, M.; Sciarra, F. Evidence that Serenoa repens extract displays an antiestrogenic activity in prostatic tissue of benign prostatic hypertrophy patients. Eur. Urol 1992, 21, 309–314. [Google Scholar]
- Vacherot, F.; Azzouz, M.; Gil-Diez-De-Medina, S.; Colombel, M.; de La Taille, A.; Lefrere Belda, M.A.; Abbou, C.C.; Raynaud, J.P.; Chopin, D.K. Induction of apoptosis and inhibition of cell proliferation by the lipido-sterolic extract of Serenoa repens (LSESr, Permixon in benign prostatic hyperplasia. Prostate 2000, 45, 259–266. [Google Scholar]
- Paubert-Braquet, M.; Cousse, H.; Raynaud, J.P.; Mencia-Huerta, J.M.; Braquet, P. Effect of the lipidosterolic extract of Serenoa repens (Permixon) and its major components on basic fibroblast growth factor-induced proliferation of cultures of human prostate biopsies. Eur. Urol 1998, 33, 340–347. [Google Scholar]
- Vela-Navarrete, R.; Escribano-Burgos, M.; Farre, A.L.; Garcia-Cardoso, J.; Manzarbeitia, F.; Carrasco, C. Serenoa repens treatment modifies bax/bcl-2 index expression and caspase-3 activity in prostatic tissue from patients with benign prostatic hyperplasia. J. Urol 2005, 173, 507–510. [Google Scholar]
- Paubert-Braquet, M.; Mencia Huerta, J.M.; Cousse, H.; Braquet, P. Effect of the lipidic lipidosterolic extract of Serenoa repens (Permixon) on the ionophore A23187-stimulated production of leukotriene B4 (LTB4) from human polymorphonuclear neutrophils. Prostaglandins Leukot. Essent. Fat. Acids 1997, 57, 299–304. [Google Scholar]
- Bonvissuto, G.; Minutoli, L.; Morgia, G.; Bitto, A.; Polito, F.; Irrera, N.; Marini, H.; Squadrito, F.; Altavilla, D. Effect of Serenoa repens, lycopene, and selenium on proinflammatory phenotype activation: An in vitro and in vivo comparison study. Urology 2011, 77, 248, e249–e216.. [Google Scholar]
- Latil, A.; Libon, C.; Templier, M.; Junquero, D.; Lantoine-Adam, F.; Nguyen, T. Hexanic lipidosterolic extract of Serenoa repens inhibits the expression of two key inflammatory mediators, MCP-1/CCL2 and VCAM-1, in vitro. BJU Int 2012, 110, E301–E307. [Google Scholar]
- Vela Navarrete, R.; Garcia Cardoso, J.V.; Barat, A.; Manzarbeitia, F.; Lopez Farre, A. BPH and inflammation: pharmacological effects of Permixon on histological and molecular inflammatory markers. Results of a double blind pilot clinical assay. Eur. Urol 2003, 44, 549–555. [Google Scholar]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar]
- Bostanci, Y.; Kazzazi, A.; Momtahen, S.; Laze, J.; Djavan, B. Correlation between benign prostatic hyperplasia and inflammation. Curr. Opin. Urol 2013, 23, 5–10. [Google Scholar]
- Di Silverio, F.; Bosman, C.; Salvatori, M.; Albanesi, L.; Proietti Pannunzi, L.; Ciccariello, M.; Cardi, A.; Salvatori, G.; Sciarra, A. Combination therapy with rofecoxib and finasteride in the treatment of men with lower urinary tract symptoms (LUTS) and benign prostatic hyperplasia (BPH). Eur. Urol 2005, 47, 72–78, ; discussion 78–79.. [Google Scholar]
- Addla, S.K.; Adeyoju, A.B.; Neilson, D.; O’Reilly, P. Diclofenac for treatment of nocturia caused by nocturnal polyuria: A prospective, randomised, double-blind, placebo-controlled crossover study. Eur. Urol 2006, 49, 720–725. [Google Scholar]
- Steiner, G.E.; Newman, M.E.; Paikl, D.; Stix, U.; Memaran-Dagda, N.; Lee, C.; Marberger, M.J. Expression and function of pro-inflammatory interleukin IL-17 and IL-17 receptor in normal, benign hyperplastic, and malignant prostate. Prostate 2003, 56, 171–182. [Google Scholar]
- Liu, L.; Li, Q.; Han, P.; Li, X.; Zeng, H.; Zhu, Y.; Wei, Q. Evaluation of interleukin-8 in expressed prostatic secretion as a reliable biomarker of inflammation in benign prostatic hyperplasia. Urology 2009, 74, 340–344. [Google Scholar]
- Schenk, J.M.; Kristal, A.R.; Neuhouser, M.L.; Tangen, C.M.; White, E.; Lin, D.W.; Kratz, M.; Thompson, I.M. Biomarkers of systemic inflammation and risk of incident, symptomatic benign prostatic hyperplasia: Results from the prostate cancer prevention trial. Am. J. Epidemiol 2010, 171, 571–582. [Google Scholar]
- Penna, G.; Mondaini, N.; Amuchastegui, S.; Degli Innocenti, S.; Carini, M.; Giubilei, G.; Fibbi, B.; Colli, E.; Maggi, M.; Adorini, L. Seminal plasma cytokines and chemokines in prostate inflammation: interleukin 8 as a predictive biomarker in chronic prostatitis/chronic pelvic pain syndrome and benign prostatic hyperplasia. Eur. Urol 2007, 51, 524–533, ; discussion 533.. [Google Scholar]
- Begley, L.A.; Kasina, S.; MacDonald, J.; Macoska, J.A. The inflammatory microenvironment of the aging prostate facilitates cellular proliferation and hypertrophy. Cytokine 2008, 43, 194–199. [Google Scholar]
- Ryschich, E.; Huszty, G.; Knaebel, H.P.; Hartel, M.; Buchler, M.W.; Schmidt, J. Transferrin receptor is a marker of malignant phenotype in human pancreatic cancer and in neuroendocrine carcinoma of the pancreas. Eur J. Cancer 2004, 40, 1418–1422. [Google Scholar]
- O’Donnell, K.A.; Yu, D.; Zeller, K.I.; Kim, J.W.; Racke, F.; Thomas-Tikhonenko, A.; Dang, C.V. Activation of transferrin receptor 1 by c-Myc enhances cellular proliferation and tumorigenesis. Mol. Cell. Biol 2006, 26, 2373–2386. [Google Scholar]
- Wada, S.; Noguchi, T.; Takeno, S.; Kawahara, K. PIK3CA and TFRC located in 3q are new prognostic factors in esophageal squamous cell carcinoma. Ann. Surg. Oncol 2006, 13, 961–966. [Google Scholar]
- Song, J.Y.; Lee, J.K.; Lee, N.W.; Jung, H.H.; Kim, S.H.; Lee, K.W. Microarray analysis of normal cervix, carcinoma in situ, and invasive cervical cancer: Identification of candidate genes in pathogenesis of invasion in cervical cancer. Int. J. Gynecol. Cancer Off. J. Int. Gynecol. Cancer Soc 2008, 18, 1051–1059. [Google Scholar]
- Schlomm, T.; Hellwinkel, O.J.; Buness, A.; Ruschhaupt, M.; Lubke, A.M.; Chun, F.K.; Simon, R.; Budaus, L.; Erbersdobler, A.; Graefen, M.; et al. Molecular cancer phenotype in normal prostate tissue. Eur. Urol 2009, 55, 885–890. [Google Scholar]
- Essaghir, A.; Demoulin, J.B. A minimal connected network of transcription factors regulated in human tumors and its application to the quest for universal cancer biomarkers. PLoS One 2012, 7, e39666. [Google Scholar]
- Tacchini, L.; Gammella, E.; de Ponti, C.; Recalcati, S.; Cairo, G. Role of HIF-1 and NF-kappaB transcription factors in the modulation of transferrin receptor by inflammatory and anti-inflammatory signals. J. Biol. Chem 2008, 283, 20674–20686. [Google Scholar]
- DiDonato, J.A.; Mercurio, F.; Karin, M. NF-κB and the link between inflammation and cancer. Immunol. Rev 2012, 246, 379–400. [Google Scholar]
- Pasparakis, M. Role of NF-κB in epithelial biology. Immunol. Rev 2012, 246, 346–358. [Google Scholar]
- Paule, B.; Terry, S.; Kheuang, L.; Soyeux, P.; Vacherot, F.; de la Taille, A. The NF-κB/IL-6 pathway in metastatic androgen-independent prostate cancer: New therapeutic approaches? World J. Urol 2007, 25, 477–489. [Google Scholar]
- Blindauer, C.A.; Leszczyszyn, O.I. Metallothioneins: Unparalleled diversity in structures and functions for metal ion homeostasis and more. Nat. Product Rep 2010, 27, 720–741. [Google Scholar]
- Swindell, W.R. Metallothionein and the biology of aging. Ageing Res. Rev 2011, 10, 132–145. [Google Scholar]
- Paubert-Braquet, M.; Richardson, F.O.; Servent-Saez, N.; Gordon, W.C.; Monge, M.C.; Bazan, N.G.; Authie, D.; Braquet, P. Effect of Serenoa repens extract (Permixon) on estradiol/testosterone-induced experimental prostate enlargement in the rat. Pharmacol. Res. Off. J. Ital. Pharmacol. Soc 1996, 34, 171–179. [Google Scholar]
- Kim, J.; Yanagihara, Y.; Kikugawa, T.; Ji, M.; Tanji, N.; Masayoshi, Y.; Freeman, M.R. A signaling network in phenylephrine-induced benign prostatic hyperplasia. Endocrinology 2009, 150, 3576–3583. [Google Scholar]
- Rick, F.G.; Schally, A.V.; Block, N.L.; Nadji, M.; Szepeshazi, K.; Zarandi, M.; Vidaurre, I.; Perez, R.; Halmos, G.; Szalontay, L. Antagonists of growth hormone-releasing hormone (GHRH) reduce prostate size in experimental benign prostatic hyperplasia. Proc. Natl. Acad. Sci. USA 2011, 108, 3755–3760. [Google Scholar]
- Castro, P.; Xia, C.; Gomez, L.; Lamb, D.J.; Ittmann, M. Interleukin-8 expression is increased in senescent prostatic epithelial cells and promotes the development of benign prostatic hyperplasia. Prostate 2004, 60, 153–159. [Google Scholar]
- Reiner, A.; Yekutieli, D.; Benjamini, Y. Identifying differentially expressed genes using false discovery rate controlling procedures. Bioinformatics 2003, 19, 368–375. [Google Scholar]
- Database for Annotation, Visualization and Integrated Discovery (DAVID). Available online: http://david.abcc.ncifcrf.gov (on accessed 4 August 2011).
- The Gene Ontology Website. Available online: http://www.geneontology.org (on accessed 4 August 2011).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BPH1 cells | ||||
|---|---|---|---|---|
| Gene | Description | FC (MA *) | FC (qPCR **) | Representative GO TERMS |
| CXCL6 | chemokine (C-X-C motif) ligand 6 | −1.67 | −0.94 | inflammatory response, immune response, chemotaxis, defense response |
| IL1A | interleukin 1, alpha | −1.02 | −1.49 | inflammatory response, immune response, defense response, cell proliferation |
| IL1B | interleukin 1, beta | −1.13 | −1.56 | inflammatory response, immune response, defense response, cell proliferation, apoptosis |
| IL1R1 | interleukin 1 receptor | −0.78 | −1.29 | immune response, defense response, cytokine-mediated signaling pathway |
| PTGS2 | prostaglandin-endoperoxide synthase 2 | −1.21 | −1.67 | fatty acid metabolic process, infllammatory response, cell poliferation, apoptosis |
| ALOX5 | arachidonate 5-lipoxygenase | −0.58 | −1.08 | fatty acid metabolic process, defense response, inflammatory response |
| PHLDA1 | pleckstrin homology-like domain, family A, member 1 | −1.10 | −1.04 | apoptosis, cell death, glycoprotein metabolic process |
| GAS1 | growth arrest-specific 1 | 1.45 | 1.79 | cell proliferation, cell cycle arrest, apoptosis, cell morphogenesis |
| DEDD | death effector domain containing | 0.77 | 0.36 | apoptosis, cell death, transcription |
| CDKN1B | cyclin-dependent kinase inhibitor 1B (p27, Kip1) | 0.50 | NS | cyclin-dependent protein kinase inhibitor, apoptosis, proliferation, cell cycle arrest |
| PrSF cells | ||||
| IL6 | interleukin 6 (interferon, beta 2) | −1.69 | −1.83 | inflammatory response, immune response, proliferation, chemotaxis, defense response |
| IL8 | interleukin 8 | −1.81 | −1.91 | inflammatory response, immune response, proliferation, chemotaxis, defense response |
| NFkBIZ | nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, zeta | −1.11 | −1.32 | inflammatory response, regulation of transcription, defense response |
| TFRC | transferrin receptor (p90, CD71) | −1.23 | −1.77 | endocytosis, vesicle-mediated transport, inflammatory response, defense response |
| ERBB3 | v-erb-b2 erythroblastic leukemia viral oncogene homolog 3 (avian) | −0.97 | −1.16 | cell surface receptor linked signal transduction, apoptosis, proliferation |
| JUN | jun oncogene | −0.76 | −1.81 | apoptosis, cell death, proliferation, transcription |
| CDKN1B | cyclin-dependent kinase inhibitor 1B (p27, Kip1) | 0.84 | 0.64 | cyclin-dependent protein kinase inhibitor, apoptosis, proliferation, cell cycle arrest |
| GAS1 | growth arrest-specific 1 | 0.73 | NS | cell proliferation, cell cycle arrest, apoptosis, cell morphogenesis |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sirab, N.; Robert, G.; Fasolo, V.; Descazeaud, A.; Vacherot, F.; Taille, A.D.l.; Terry, S. Lipidosterolic Extract of Serenoa Repens Modulates the Expression of Inflammation Related-Genes in Benign Prostatic Hyperplasia Epithelial and Stromal Cells. Int. J. Mol. Sci. 2013, 14, 14301-14320. https://doi.org/10.3390/ijms140714301
Sirab N, Robert G, Fasolo V, Descazeaud A, Vacherot F, Taille ADl, Terry S. Lipidosterolic Extract of Serenoa Repens Modulates the Expression of Inflammation Related-Genes in Benign Prostatic Hyperplasia Epithelial and Stromal Cells. International Journal of Molecular Sciences. 2013; 14(7):14301-14320. https://doi.org/10.3390/ijms140714301
Chicago/Turabian StyleSirab, Nanor, Grégoire Robert, Virginie Fasolo, Aurélien Descazeaud, Francis Vacherot, Alexandre De la Taille, and Stéphane Terry. 2013. "Lipidosterolic Extract of Serenoa Repens Modulates the Expression of Inflammation Related-Genes in Benign Prostatic Hyperplasia Epithelial and Stromal Cells" International Journal of Molecular Sciences 14, no. 7: 14301-14320. https://doi.org/10.3390/ijms140714301