Involvement of miR-20a in Promoting Gastric Cancer Progression by Targeting Early Growth Response 2 (EGR2)

Abstract
:1. Introduction
2. Results
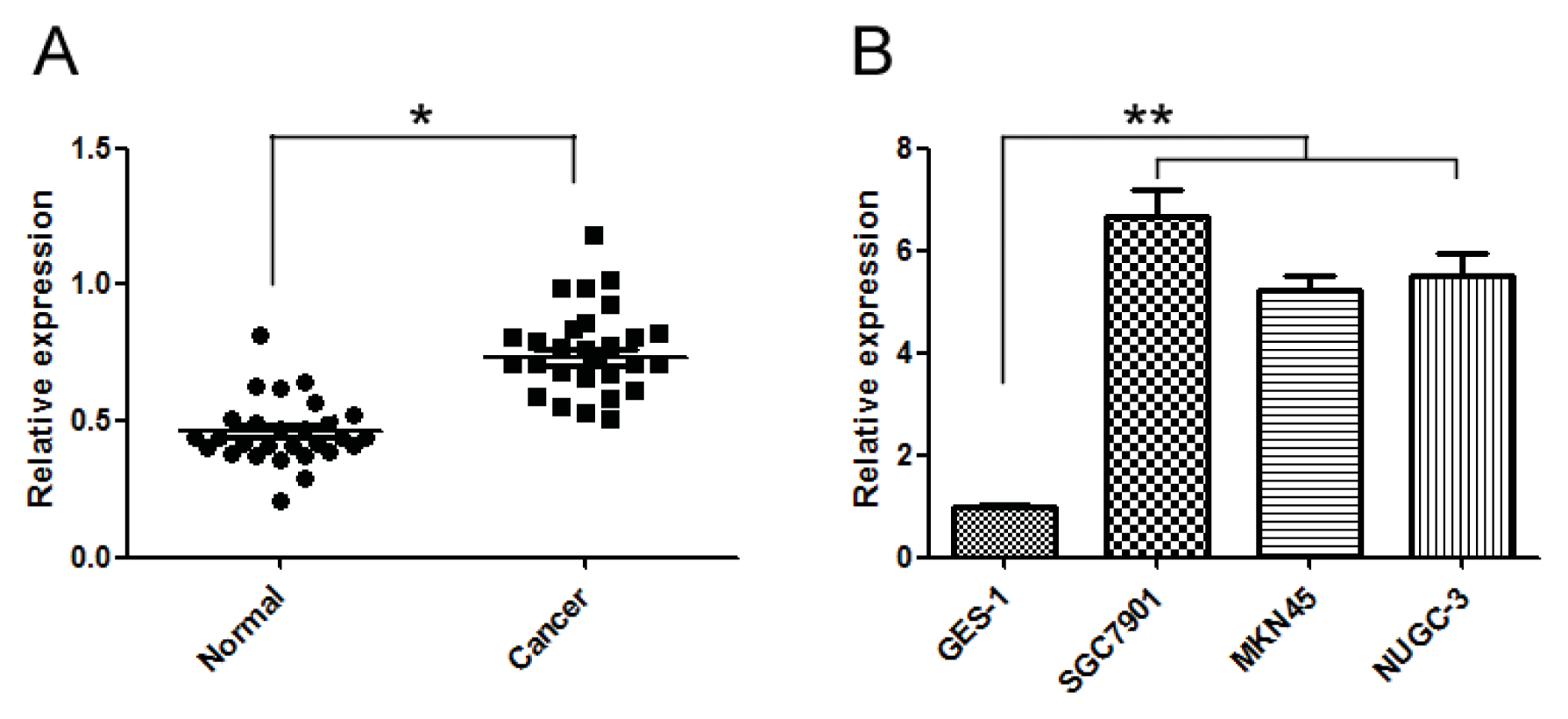
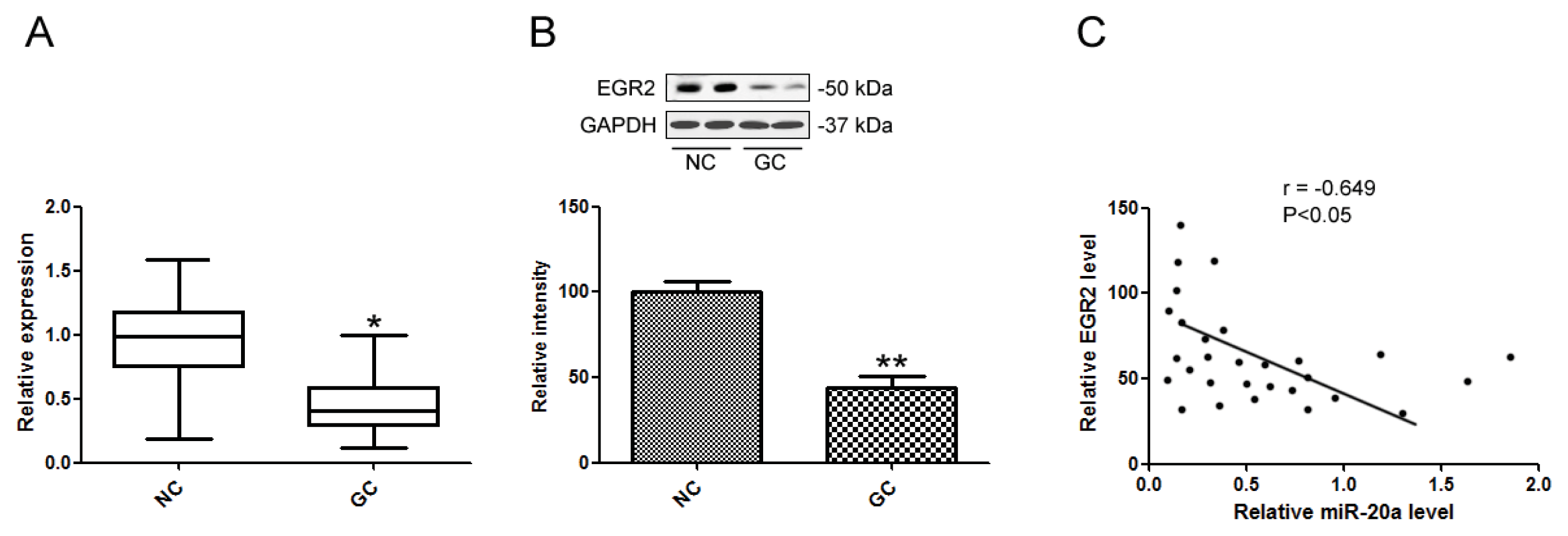
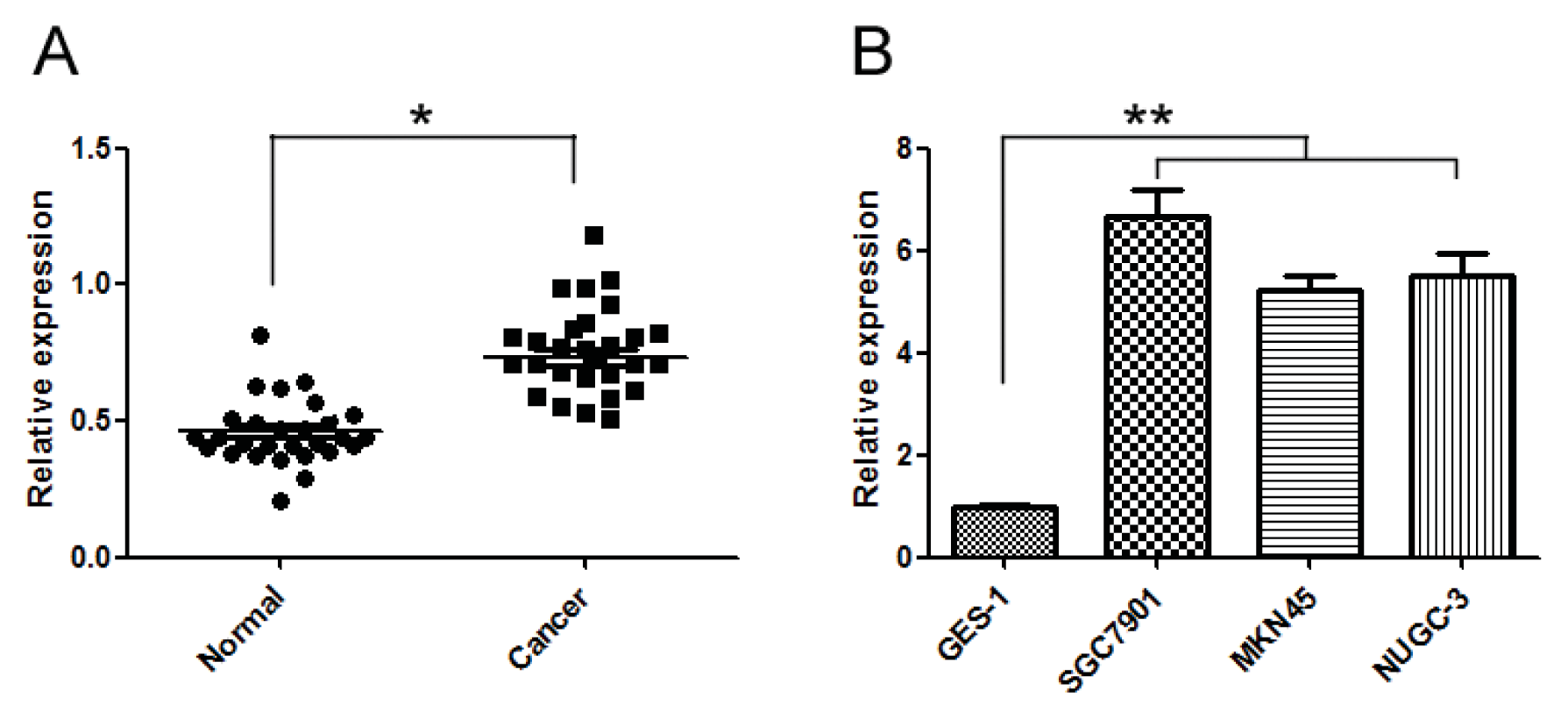
2.1. miR-20a Was Increased in GC Tissue Samples and Cell Lines
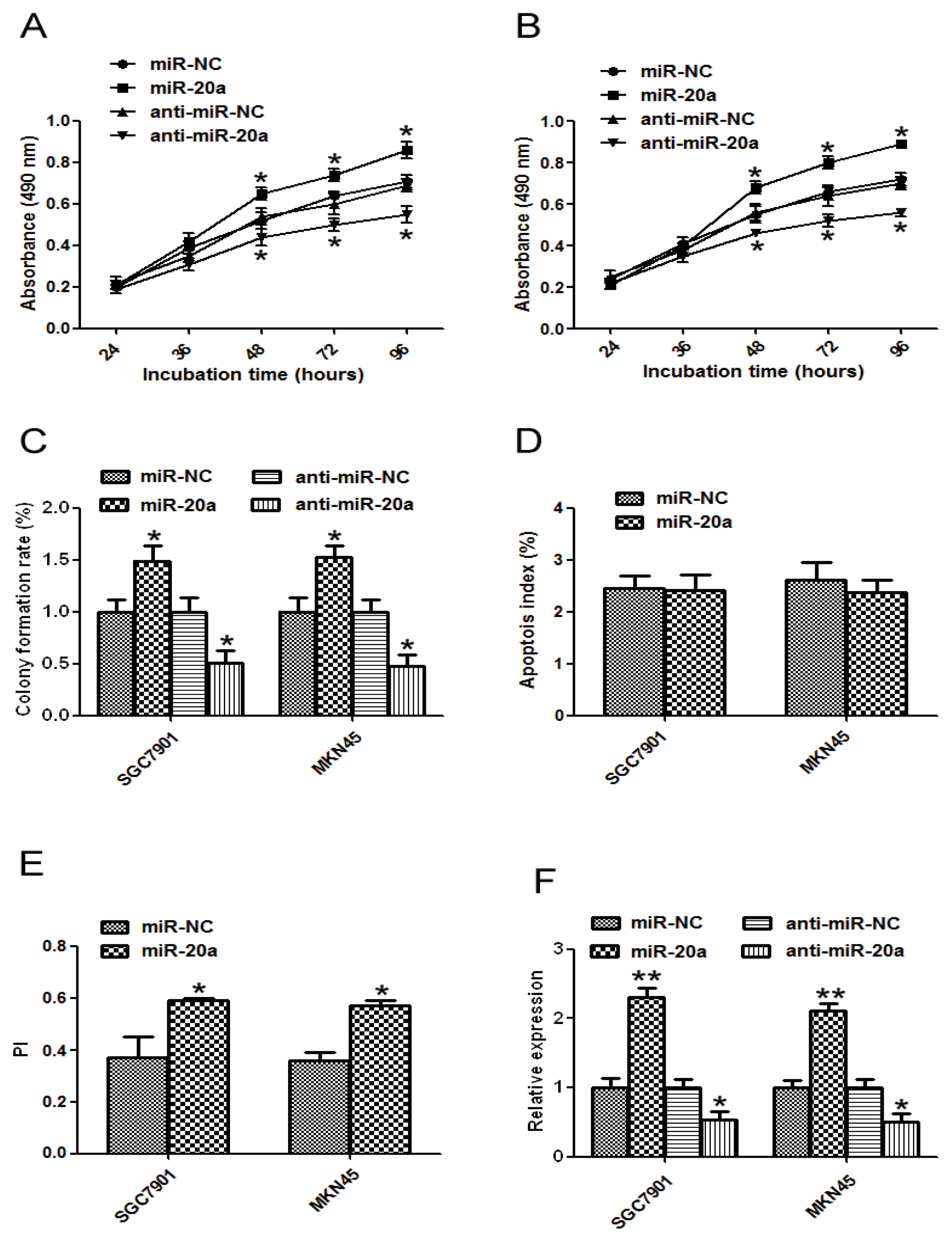
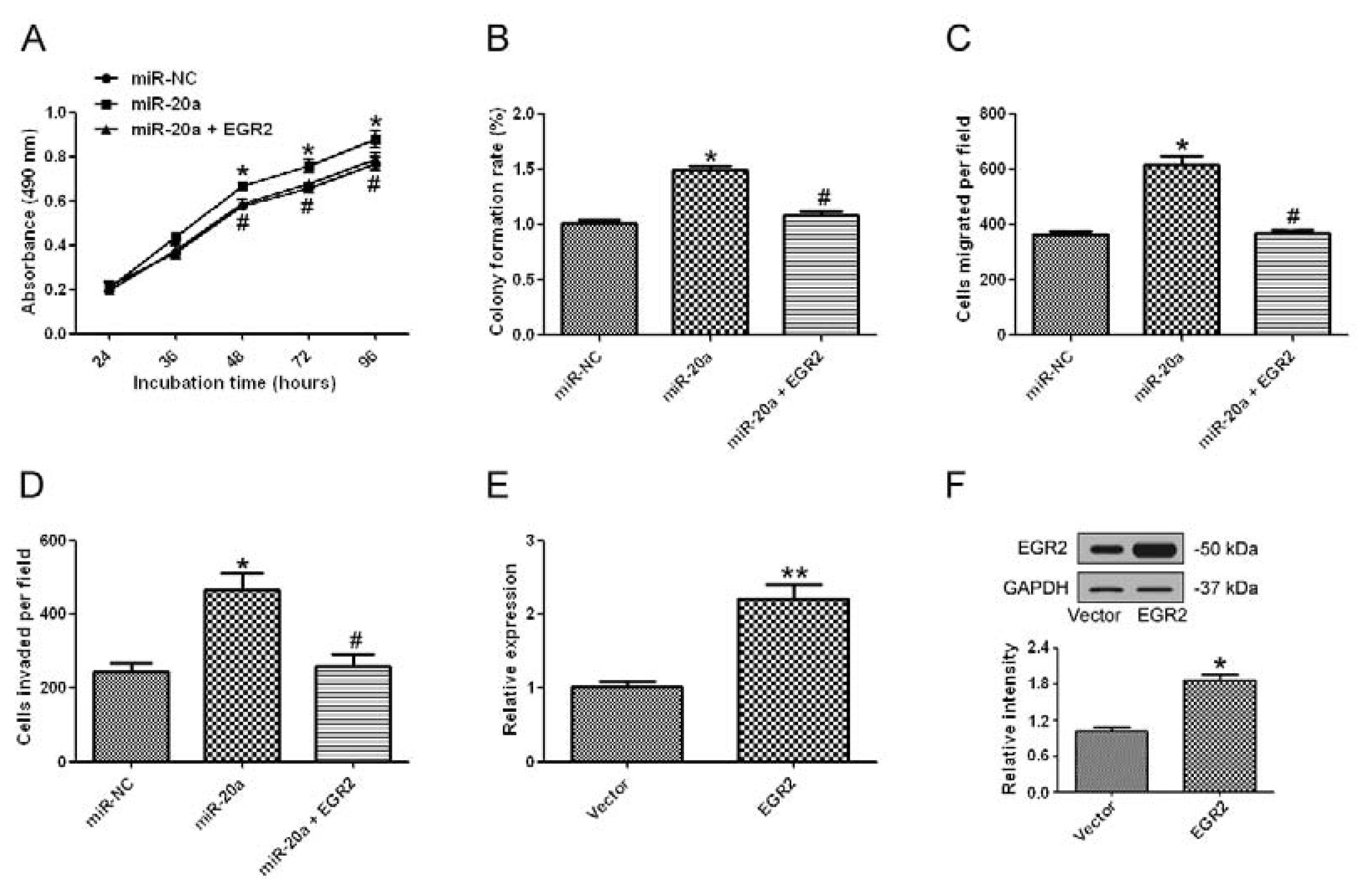
2.2. miR-20a Promoted Growth of GC Cells
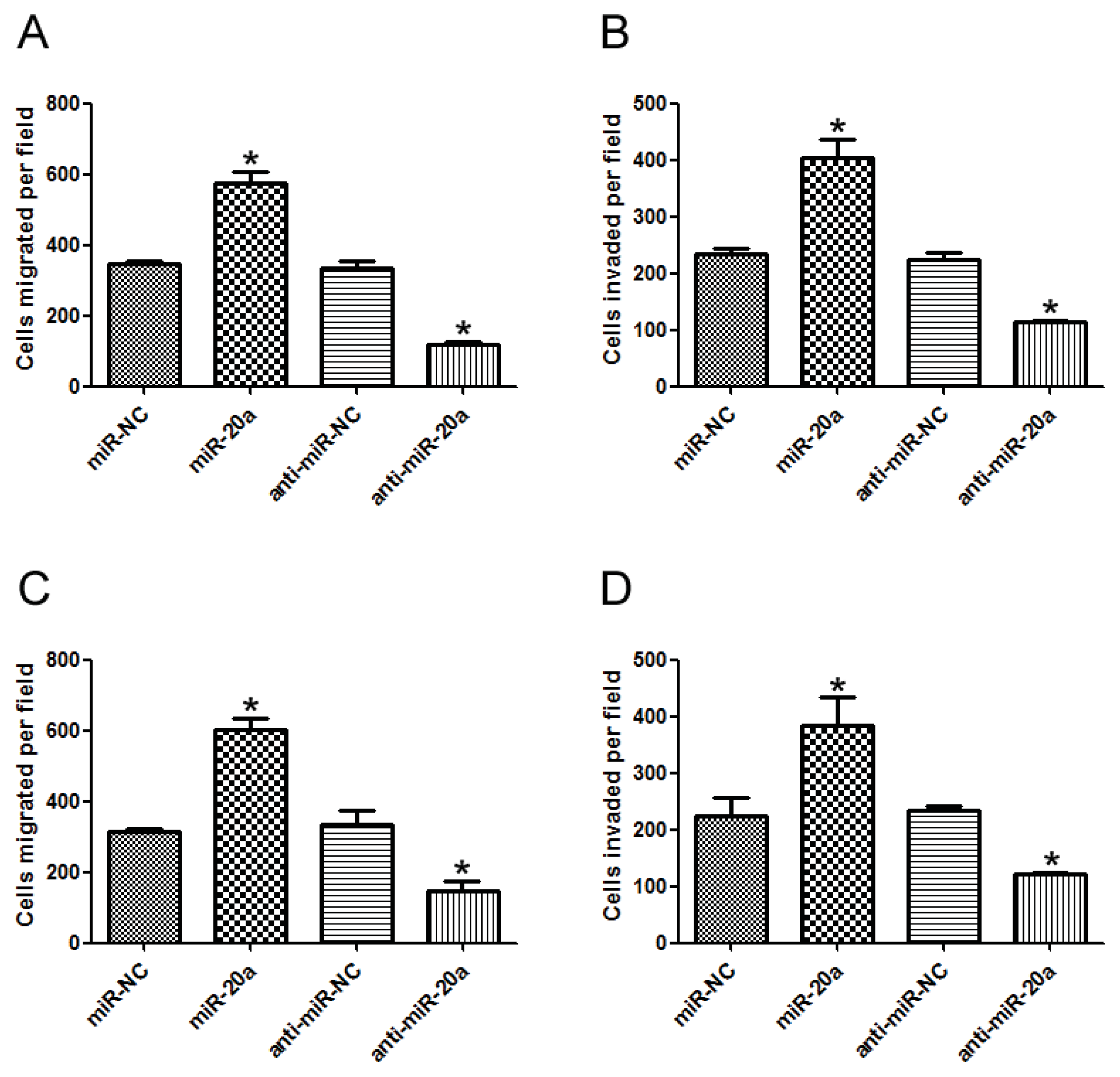
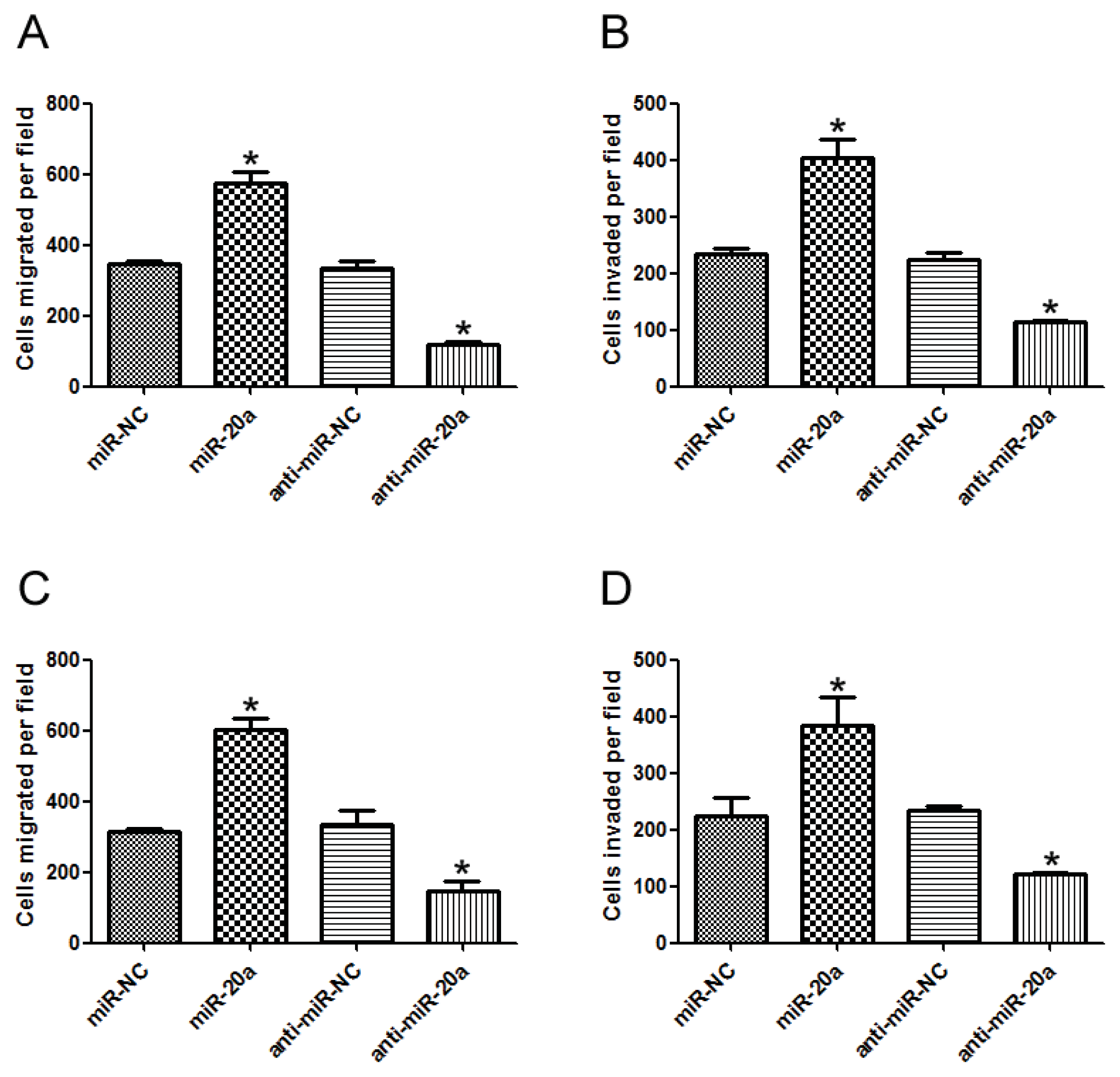
2.3. miR-20a Promoted Migration and Invasion of GC Cells
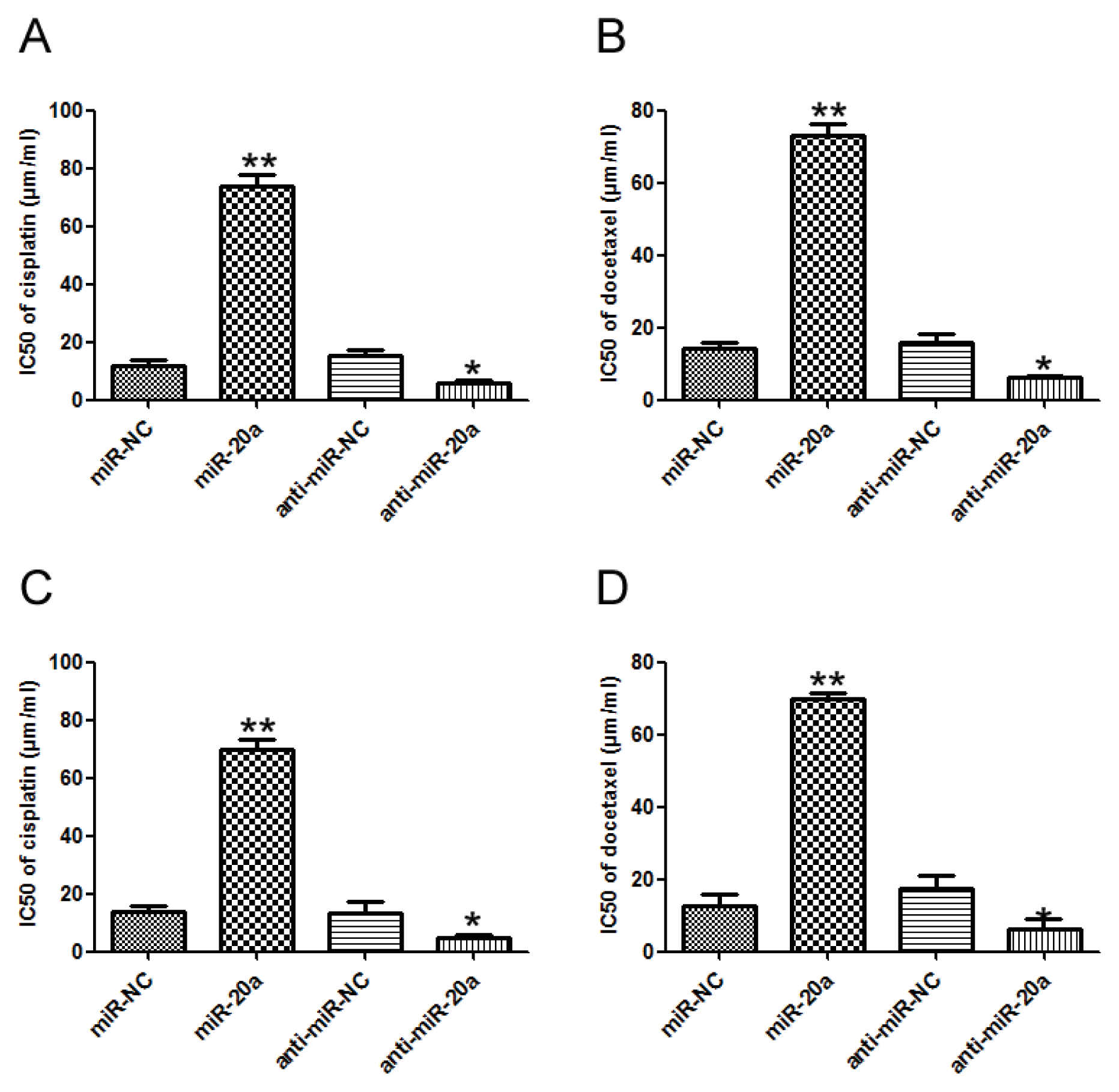
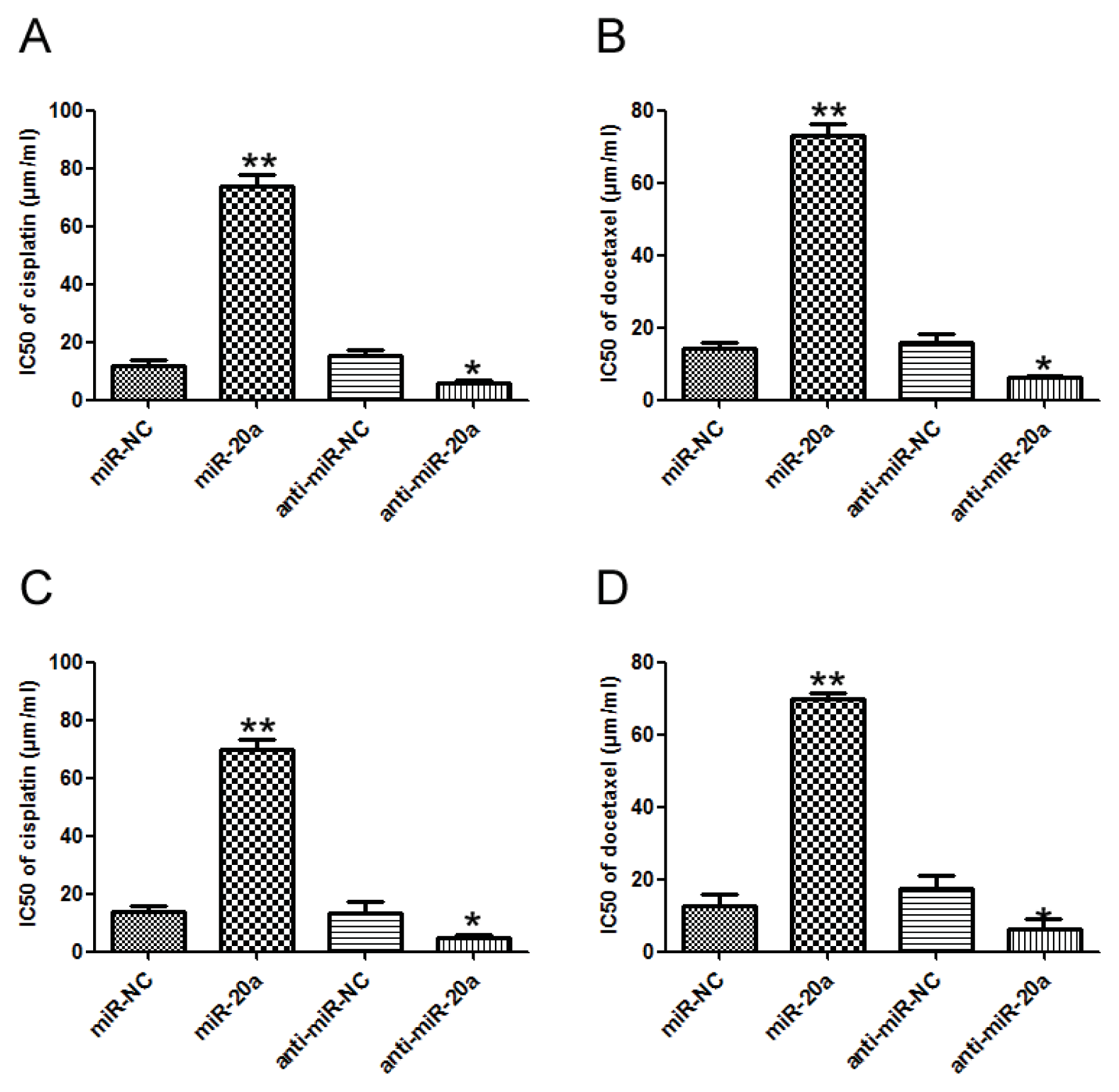
2.4. miR-20a Promoted Chemoresistance of GC Cells
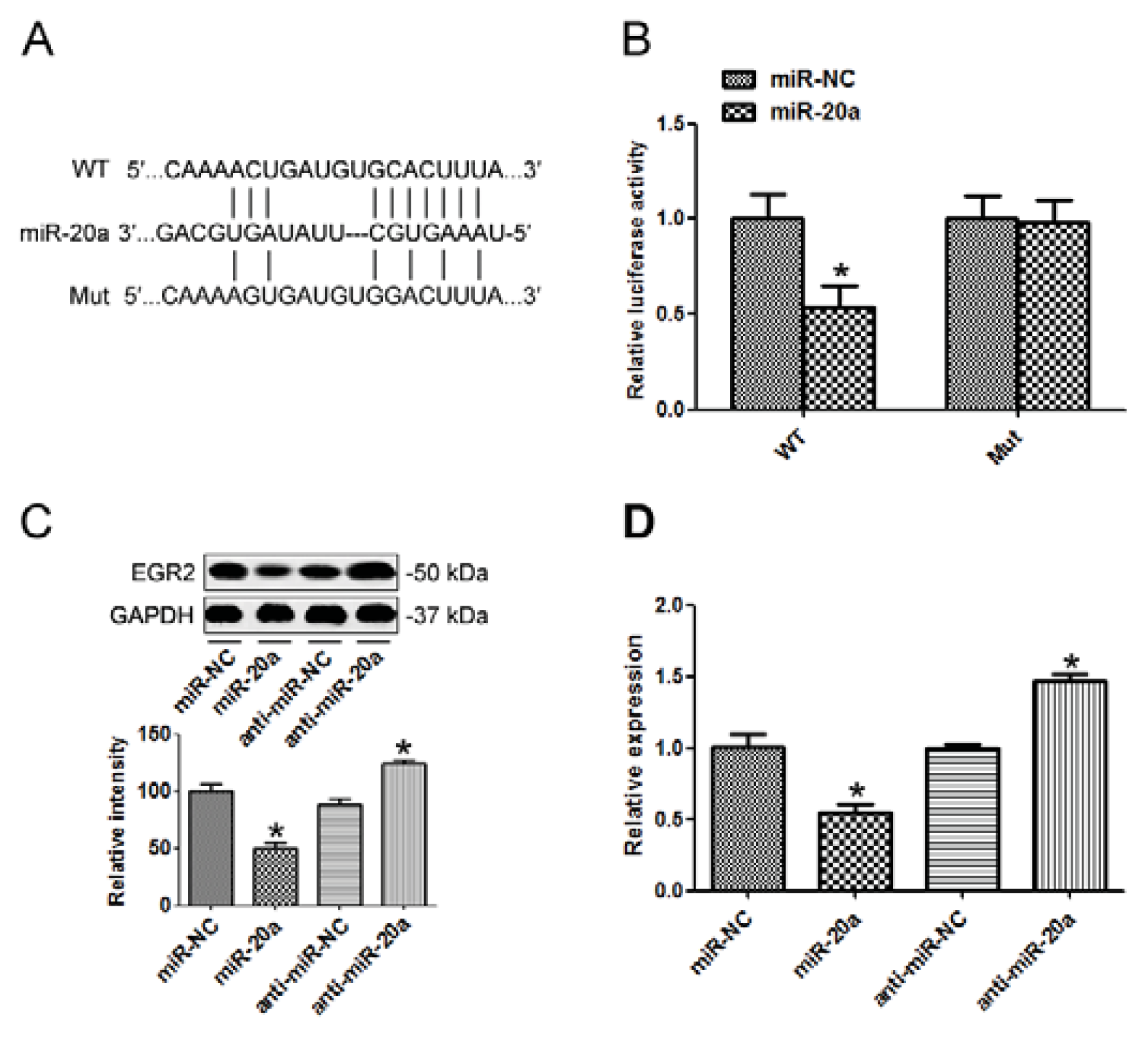
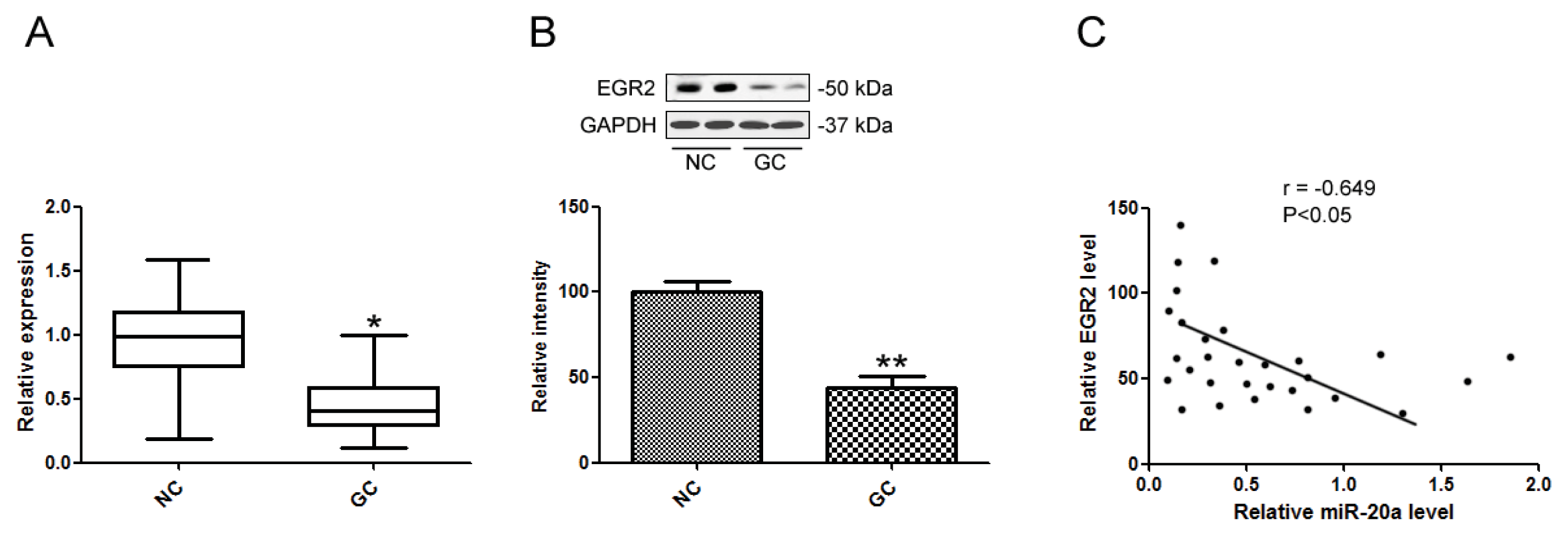
2.5. EGR2 Was a Direct Target of miR-20a
2.7. miR-20a Promoted GC Progression by Targeting EGR2
3. Discussion
4. Materials and Methods
4.1. Cell lines and Tissue Samples
4.2. Plasmids and Transfection
4.3. Quantitative Real Time PCR
4.4. In Vitro Cell Proliferation Assay
4.5. Colony Formation Assay
4.6. In Vitro Migration and Invasion Assay
4.7. Luciferase Reporter Assay
4.8. Western Blot
4.9. Flow Cytometric Analysis of Apoptosis
4.10. Cell Cycle Analysis by Flow Cytometry
4.11. Statistical Analysis
5. Conclusions
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin 2011, 61, 69–90. [Google Scholar]
- Ali, Z.; Deng, Y.; Ma, C. Progress of research in gastric cancer. J. Nanosci. Nanotechnol 2012, 12, 8241–8248. [Google Scholar]
- Link, A.; Kupcinskas, J.; Wex, T.; Malfertheiner, P. Macro-role of microRNA in gastric cancer. Dig. Dis 2012, 30, 255–267. [Google Scholar]
- Song, J.H.; Meltzer, S.J. microRNAs in pathogenesis, diagnosis, and treatment of gastroesophageal cancers. Gastroenterology 2012, 143, 35–47. [Google Scholar]
- Liu, H. microRNAs in breast cancer initiation and progression. Cell. Mol. Life Sci 2012, 69, 3587–3599. [Google Scholar]
- Rossi, S.; di Narzo, A.F.; Mestdagh, P.; Jacobs, B.; Bosman, F.T.; Gustavsson, B.; Majoie, B.; Roth, A.; Vandesompele, J.; Rigoutsos, I.; et al. microRNAs in colon cancer: A roadmap for discovery. FEBS Lett 2012, 586, 3000–3007. [Google Scholar] [Green Version]
- Du, L.; Pertsemlidis, A. microRNA regulation of cell viability and drug sensitivity in lung cancer. Expert Opin. Biol. Ther 2012, 12, 1221–1239. [Google Scholar]
- O’Kelly, F.; Marignol, L.; Meunier, A.; Lynch, T.H.; Perry, A.S.; Hollywood, D. microRNAs as putative mediators of treatment response in prostate cancer. Nat. Rev. Urol 2012, 9, 397–407. [Google Scholar]
- Guo, M.M.; Hu, L.H.; Wang, Y.Q.; Chen, P.; Huang, J.G.; Lu, N.; He, J.H.; Liao, C.G. miR-22 is down-regulated in gastric cancer, and its overexpression inhibits cell migration and invasion via targeting transcription factor Sp1. Med. Oncol 2013, 30, 542. [Google Scholar]
- Yao, Q.; Cao, Z.; Tu, C.; Zhao, Y.; Liu, H.; Zhang, S. microRNA-146a acts as a metastasis suppressor in gastric cancer by targeting WASF2. Cancer Lett 2013, 335, 219–224. [Google Scholar]
- Zheng, L.; Qi, T.; Yang, D.; Qi, M.; Li, D.; Xiang, X.; Huang, K.; Tong, Q. microRNA-9 suppresses the proliferation, invasion and metastasis of gastric cancer cells through targeting cyclin D1 and Ets1. PLoS One 2013, 8, e55719. [Google Scholar]
- Wang, F.; Li, T.; Zhang, B.; Li, H.; Wu, Q.; Yang, L.; Nie, Y.; Wu, K.; Shi, Y.; Fan, D. microRNA-19a/b regulates multidrug resistance in human gastric cancer cells by targeting PTEN. Biochem. Biophys. Res. Commun 2013, 434, 688–694. [Google Scholar]
- Wang, M.; Li, C.; Yu, B.; Su, L.; Li, J.; Ju, J.; Yu, Y.; Gu, Q.; Zhu, Z.; Liu, B. Overexpressed miR-301a promotes cell proliferation and invasion by targeting RUNX3 in gastric cancer. J. Gastroenterol. 2013. [Epub ahead of print].. [Google Scholar]
- An, J.; Pan, Y.; Yan, Z.; Li, W.; Cui, J.; Jiao, Y.; Tian, L.; Xing, R.; Lu, Y. miR-23a in Amplified 19p13.13 loci targets metallothionein 2A and promotes growth in gastric cancer cells. J. Cell. Biochem. 2013, 114, 2160–2169. [Google Scholar]
- Yan, H.J.; Liu, W.S.; Sun, W.H.; Wu, J.; Ji, M.; Wang, Q.; Zheng, X.; Jiang, J.T.; Wu, C.P. miR-17-5p inhibitor enhances chemosensitivity to gemcitabine via upregulating Bim expression in pancreatic cancer cells. Dig. Dis. Sci 2012, 57, 3160–3167. [Google Scholar]
- Kang, H.W.; Wang, F.; Wei, Q.; Zhao, Y.F.; Liu, M.; Li, X.; Tang, H. miR-20a promotes migration and invasion by regulating TNKS2 in human cervical cancer cells. FEBS Lett 2012, 586, 897–904. [Google Scholar]
- Huang, G.; Nishimoto, K.; Zhou, Z.; Hughes, D.; Kleinerman, E.S. miR-20a encoded by the miR-17-92 cluster increases the metastatic potential of osteosarcoma cells by regulating Fas expression. Cancer Res 2012, 72, 908–916. [Google Scholar]
- Gigek, C.O.; Chen, E.S.; Calcagno, D.Q.; Wisnieski, F.; Burbano, R.R.; Smith, M.A. Epigenetic mechanisms in gastric cancer. Epigenomics 2012, 4, 279–294. [Google Scholar]
- Li, X.; Pan, J.H.; Song, B.; Xiong, E.Q.; Chen, Z.W.; Zhou, Z.S.; Su, Y.P. Suppression of CX43 expression by miR-20a in the progression of human prostate cancer. Cancer Biol. Ther 2012, 13, 890–898. [Google Scholar]
- Wang, M.; Gu, H.; Wang, S.; Qian, H.; Zhu, W.; Zhang, L.; Zhao, C.; Tao, Y.; Xu, W. Circulating miR-17-5p and miR-20a: Molecular markers for gastric cancer. Mol. Med. Rep 2012, 5, 1514–1520. [Google Scholar]
- Chai, H.; Liu, M.; Tian, R.; Li, X.; Tang, H. miR-20a targets BNIP2 and contributes chemotherapeutic resistance in colorectal adenocarcinoma SW480 and SW620 cell lines. Acta Biochim. Biophys. Sin 2011, 43, 217–225. [Google Scholar]
- Philippe, L.; Alsaleh, G.; Pichot, A.; Ostermann, E.; Zuber, G.; Frisch, B.; Sibilia, J.; Pfeffer, S.; Bahram, S.; Wachsmann, D.; et al. miR-20a regulates ASK1 expression and TLR4-dependent cytokine release in rheumatoid fibroblast-like synoviocytes. Annu. Rheum. Dis 2013, 72, 1071–1079. [Google Scholar]
- Chang, Y.; Liu, C.; Yang, J.; Liu, G.; Feng, F.; Tang, J.; Hu, L.; Li, L.; Jiang, F.; Chen, C.; et al. miR-20a triggers metastasis of gallbladder carcinoma. J. Hepatol. 2013. [Google Scholar] [CrossRef]
- Fan, M.Q.; Huang, C.B.; Gu, Y.; Xiao, Y.; Sheng, J.X.; Zhong, L. Decrease expression of microRNA-20a promotes cancer cell proliferation and predicts poor survival of hepatocellular carcinoma. J. Exp. Clin. Cancer Res 2013, 32, 21. [Google Scholar]
- Wang, M.; Gu, H.; Qian, H.; Zhu, W.; Zhao, C.; Zhang, X.; Tao, Y.; Zhang, L.; Xu, W. miR-17-5p/20a are important markers for gastric cancer and murine double minute 2 participates in their functional regulation. Eur. J. Cancer 2013, 49, 2010–2021. [Google Scholar]
- Wang, F.; Song, X.; Li, X.; Xin, J.; Wang, S.; Yang, W.; Wang, J.; Wu, K.; Chen, X.; Liang, J.; et al. Noninvasive visualization of microRNA-16 in the chemoresistance of gastric cancer using a dual reporter gene imaging system. PLoS One 2013, 8, e61792. [Google Scholar]
- Tekiner, T.A.; Basaga, H. Role of microRNA deregulation in breast cancer cell chemoresistance and stemness. Curr. Med. Chem. 2013. [Epub ahead of print]. [Google Scholar]
- Wang, Y.Q.; Guo, R.D.; Guo, R.M.; Sheng, W.; Yin, L.R. microRNA-182 promotes cell growth, invasion, and chemoresistance by targeting programmed cell death 4 (PDCD4) in human ovarian carcinomas. J. Cell. Biochem 2013, 114, 1464–1473. [Google Scholar]
- Joseph, L.J.; le Beau, M.M.; Jamieson, G.A., Jr; Acharya, S.; Shows, T.B.; Rowley, J.D.; Sukhatme, V.P. Molecular cloning, sequencing, and mapping of EGR2, a human early growth response gene encoding a protein with “zinc-binding finger” structure. Proc. Natl. Acad. Sci. USA 1988, 85, 7164–7168. [Google Scholar]
- Dzialo-Hatton, R.; Milbrandt, J.; Hockett, R.D., Jr; Weaver, C.T. Differential expression of Fas ligand in Th1 and Th2 cells is regulated by early growth response gene and NF-AT family members. J. Immunol. 2001, 166, 4534–4542. [Google Scholar]
- Yokota, I.; Sasaki, Y.; Kashima, L.; Idogawa, M.; Tokino, T. Identification and characterization of early growth response 2, a zinc-finger transcription factor, as a p53-regulated proapoptotic gene. Int. J. Oncol 2010, 37, 1407–1416. [Google Scholar]
- Wu, Q.; Jin, H.; Yang, Z.; Luo, G.; Lu, Y.; Li, K.; Ren, G.; Su, T.; Pan, Y.; Feng, B.; et al. miR-150 promotes gastric cancer proliferation by negatively regulating the pro-apoptotic gene EGR2. Biochem. Biophys. Res. Commun 2010, 392, 340–345. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathologic features | No. | Relative expression of miR-20a a | p-value b |
|---|---|---|---|
| Gender | 0.727 | ||
| Male | 18 | 0.72 (0.51–1.33) | |
| Female | 10 | 0.70 (0.52–1.02) | |
| Site of tumor | 0.810 | ||
| Upper stomach | 8 | 0.68 (0.52–1.00) | |
| Middle stomach | 6 | 0.70 (0.51–1.22) | |
| Lower stomach | 14 | 0.71 (0.51–1.32) | |
| Differentiation | 0.655 | ||
| Poor | 12 | 0.73 (0.59–1.33) | |
| Moderate | 16 | 0.72 (0.52–1.12) | |
| Metastasis | 0.003 | ||
| N0 | 4 | 0.55 (0.51–0.61) | |
| N1 | 5 | 0.62 (0.51–0.64) | |
| N2 | 9 | 0.74 (0.52–1.09) | |
| N3 | 9 | 0.92 (0.51–1.33) |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, X.; Zhang, Z.; Yu, M.; Li, L.; Du, G.; Xiao, W.; Yang, H. Involvement of miR-20a in Promoting Gastric Cancer Progression by Targeting Early Growth Response 2 (EGR2). Int. J. Mol. Sci. 2013, 14, 16226-16239. https://doi.org/10.3390/ijms140816226
Li X, Zhang Z, Yu M, Li L, Du G, Xiao W, Yang H. Involvement of miR-20a in Promoting Gastric Cancer Progression by Targeting Early Growth Response 2 (EGR2). International Journal of Molecular Sciences. 2013; 14(8):16226-16239. https://doi.org/10.3390/ijms140816226
Chicago/Turabian StyleLi, Xiangsheng, Zhichao Zhang, Ming Yu, Liqi Li, Guangsheng Du, Weidong Xiao, and Hua Yang. 2013. "Involvement of miR-20a in Promoting Gastric Cancer Progression by Targeting Early Growth Response 2 (EGR2)" International Journal of Molecular Sciences 14, no. 8: 16226-16239. https://doi.org/10.3390/ijms140816226