Citrus Leaf Volatiles as Affected by Developmental Stage and Genetic Type


Abstract
:
1. Introduction
2. Results and Discussion
2.1. Analysis of Citrus Leaf Volatiles by HS-SPME-GC-MS
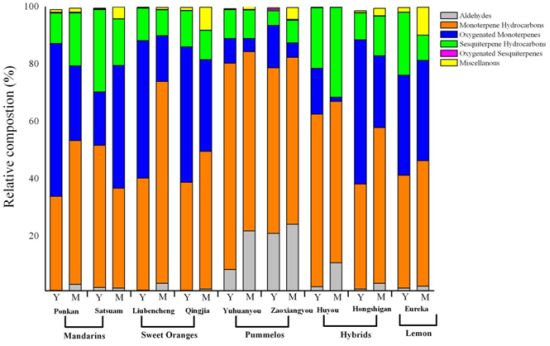
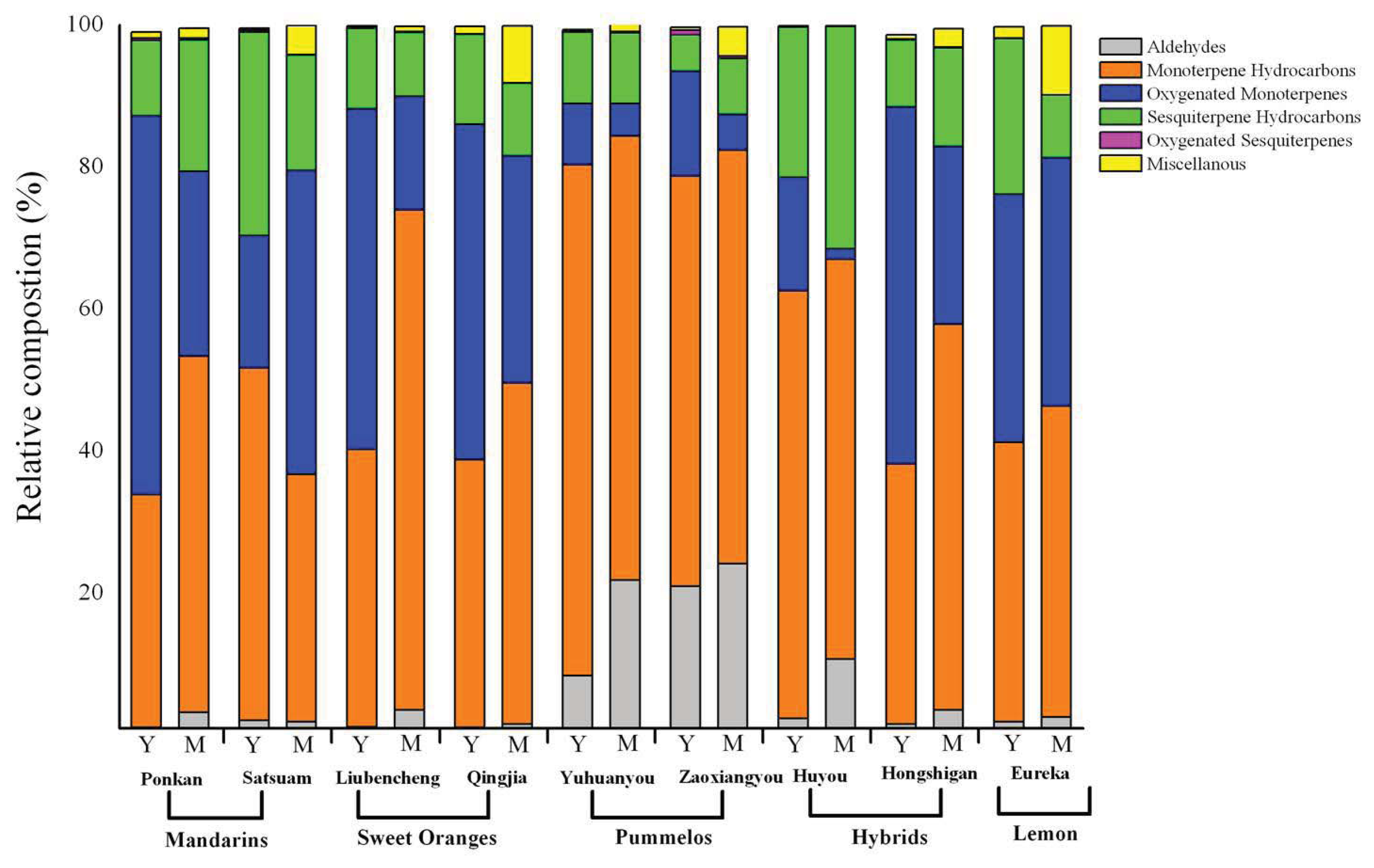
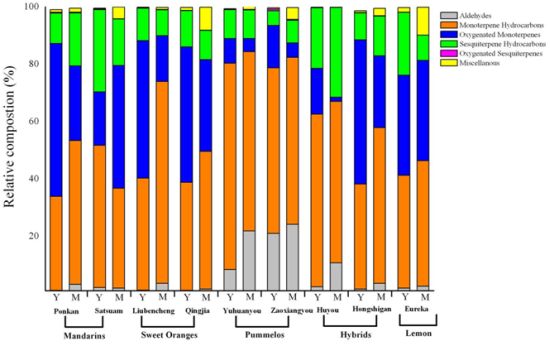
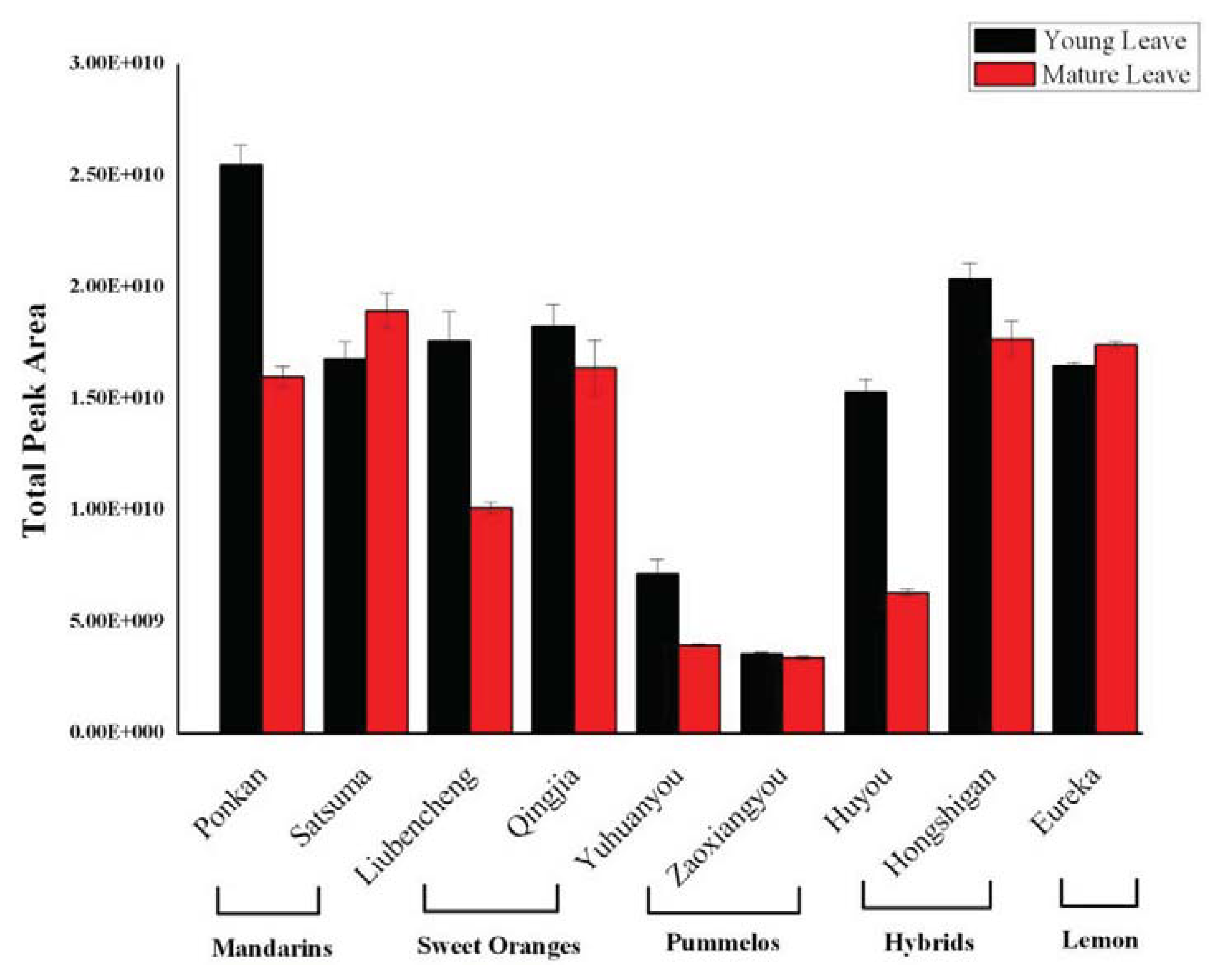
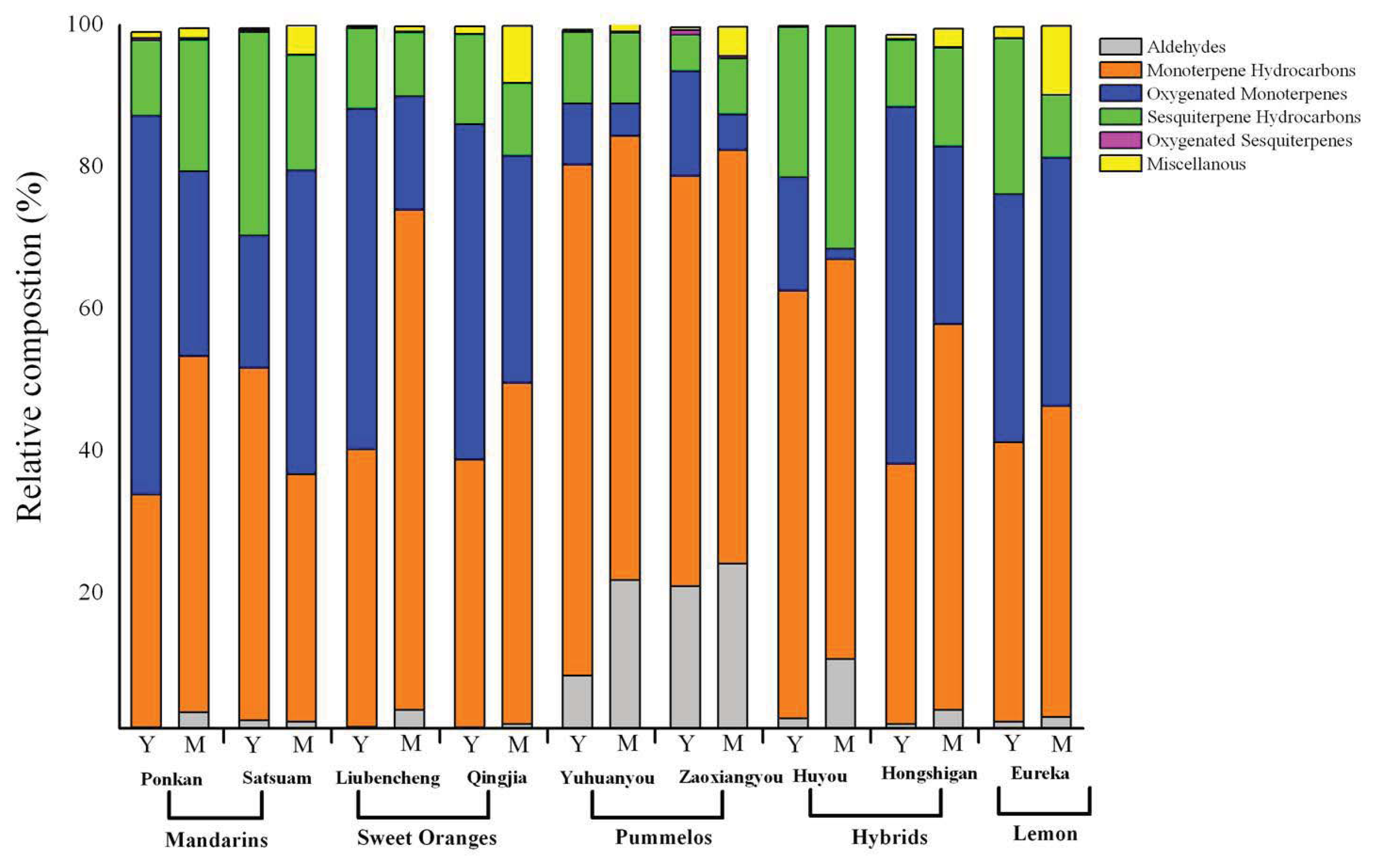
2.2. Variation in Total Volatiles Content and Major Chemical Classes from Young and Mature Leaf
2.3. Variation in Volatile Constituents from Nine Citrus Cultivars
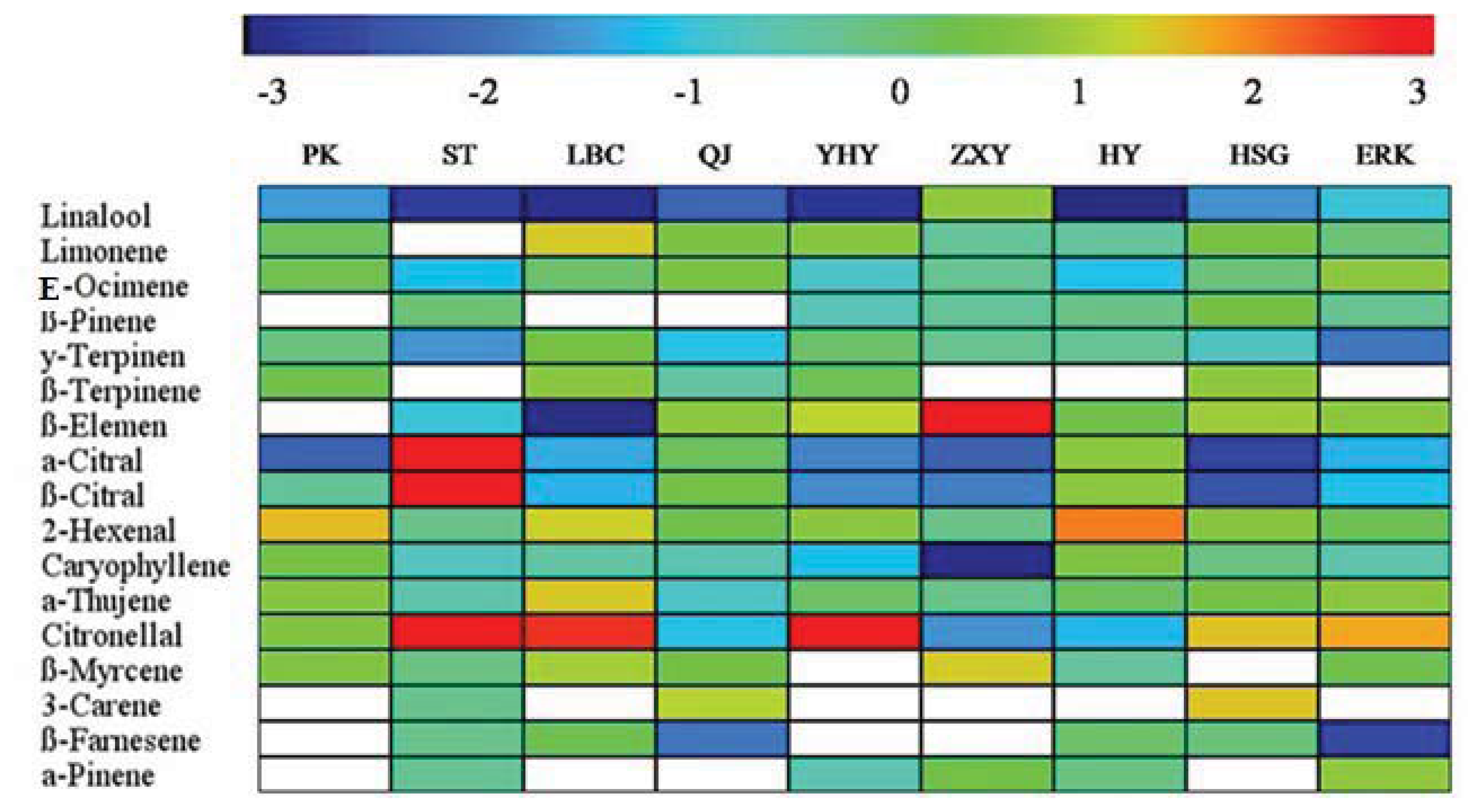
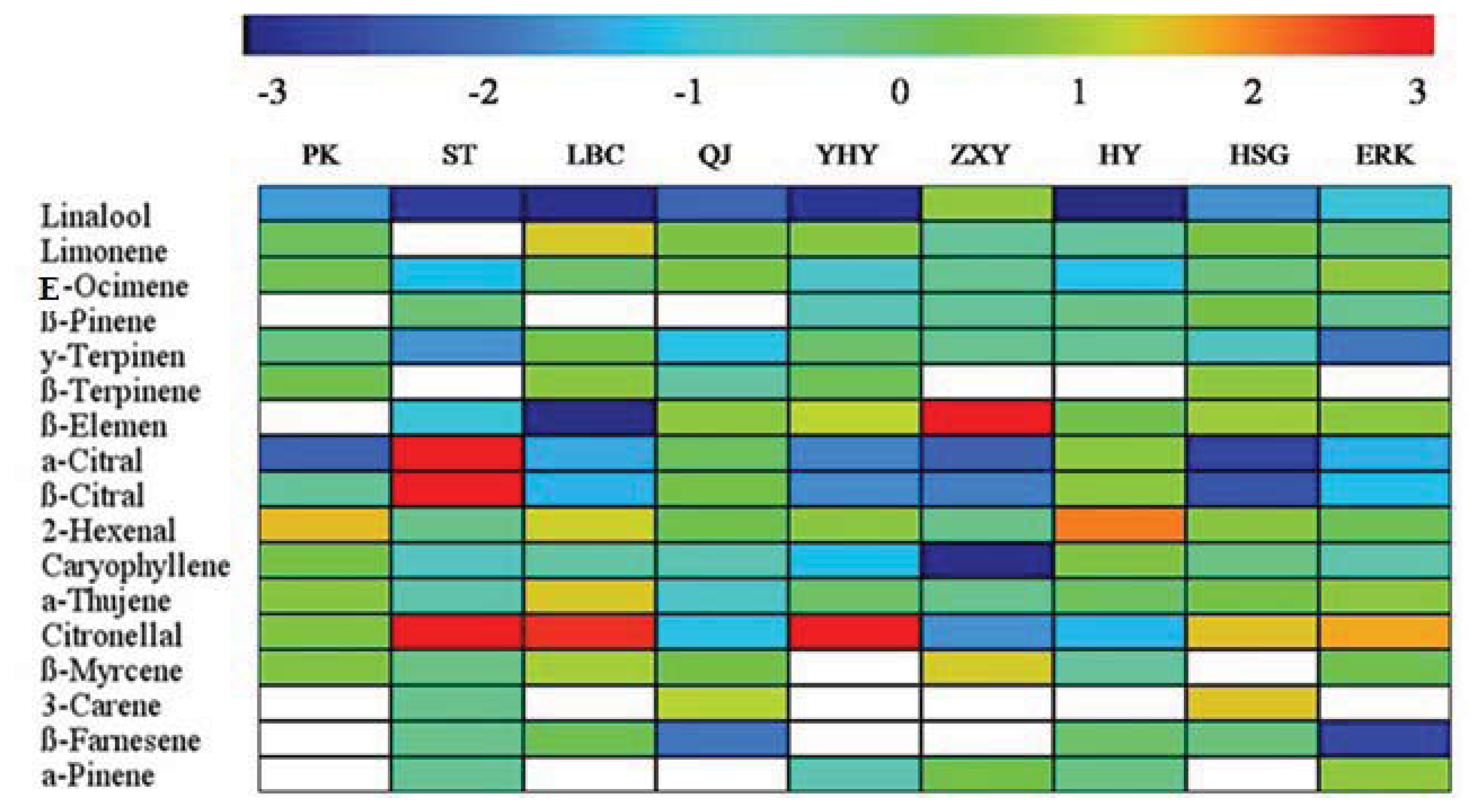
2.4. Changes in Volatile Abundance during Leaf Development
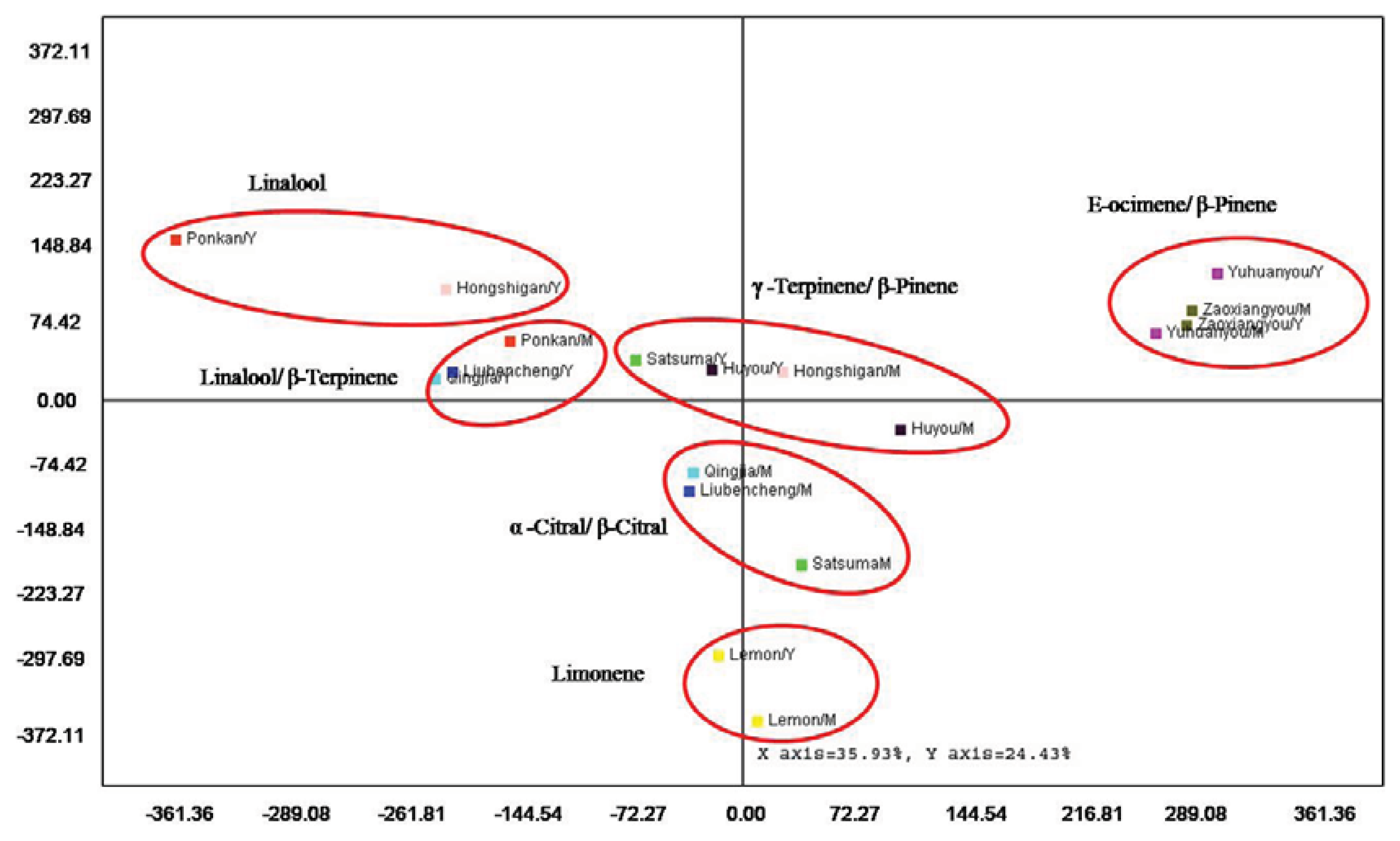
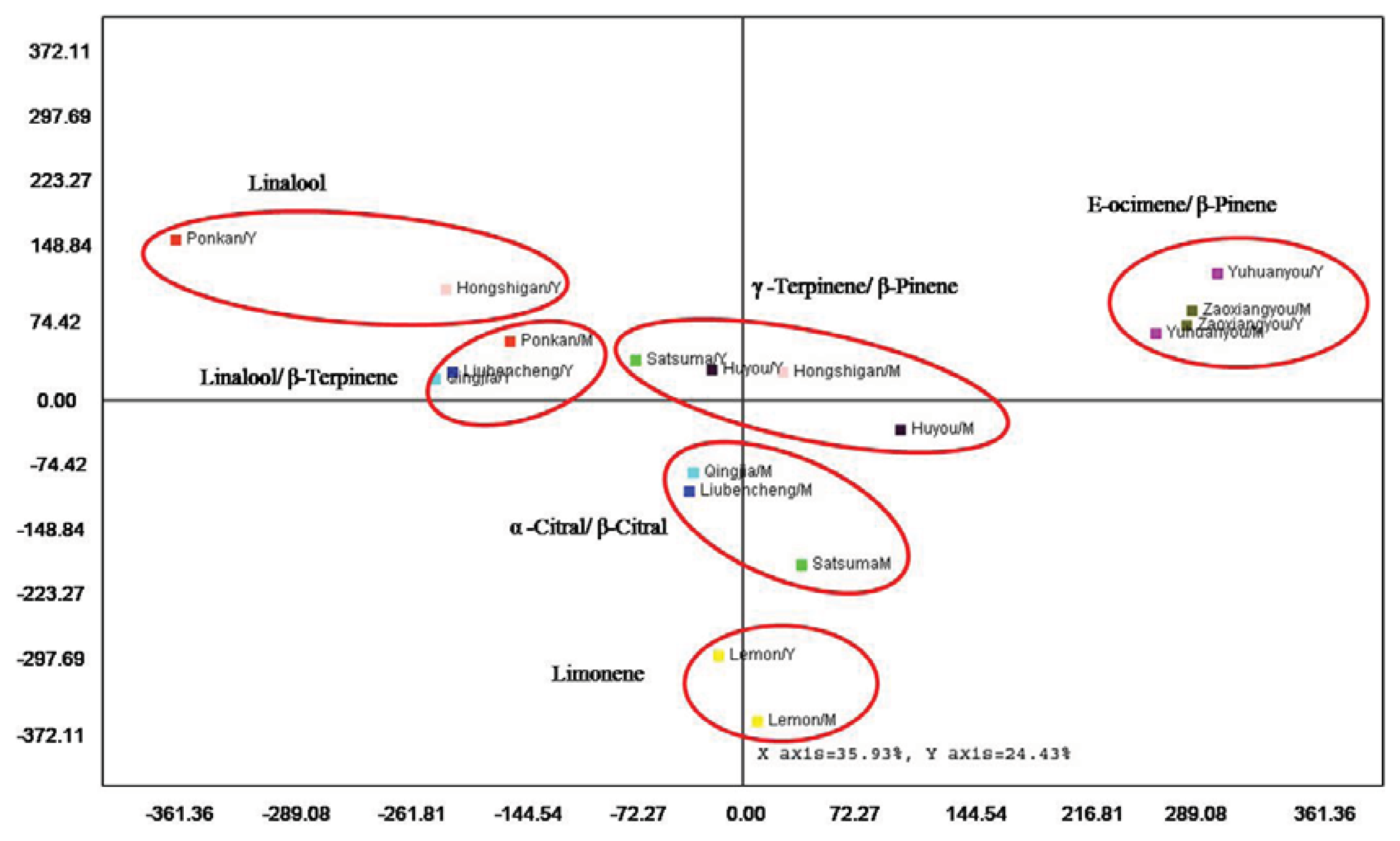
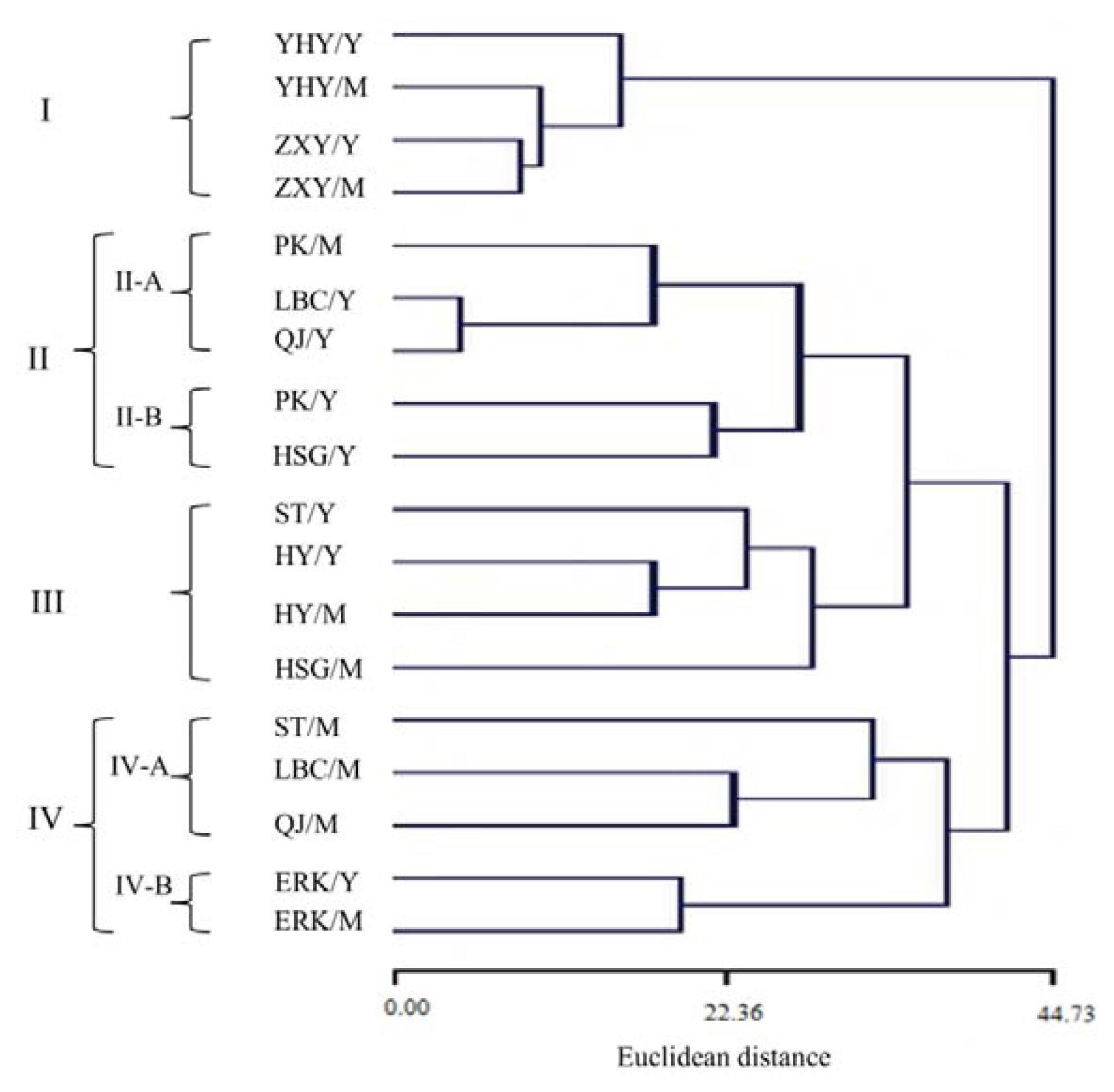
2.5. Multivariate Analysis for Leaf Volatiles among Nine Cultivars
3. Experimental Section
3.1. Materials
3.2. HS-SPME Extraction
3.3. GC-MS Analysis
3.4. Electronic Nose Measurements
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Vickers, C.E.; Gershenzon, J.; Lerdau, M.T.; Loreto, F. A unified mechanism of action for volatile isoprenoids in plant abiotic stress. Nat. Chem. Biol 2009, 5, 283–291. [Google Scholar]
- Tholl, D.; Sohrabi, R.; Huh, J.-H.; Lee, S. The biochemistry of homoterpenes–common constituents of floral and herbivore-induced plant volatile bouquets. Phytochemistry 2011, 72, 1635–1646. [Google Scholar]
- Mahmoud, S.S.; Croteau, R.B. Metabolic engineering of essential oil yield and composition in mint by altering expression of deoxyxylulose phosphate reductoisomerase and menthofuran synthase. Proc. Natl. Acad. Sci. USA 2001, 98, 8915–8920. [Google Scholar]
- Sapir-Mir, M.; Mett, A.; Belausov, E.; Tal-Meshulam, S.; Frydman, A.; Gidoni, D.; Eyal, Y. Peroxisomal localization of arabidopsis isopentenyl diphosphate isomerases suggests that part of the plant isoprenoid mevalonic acid pathway is compartmentalized to peroxisomes. Plant Physiol 2008, 148, 1219–1228. [Google Scholar]
- Takahashi, S.; Koyama, T. Structure and function of cis-prenyl chain elongating enzymes. Chem. Rec 2006, 6, 194–205. [Google Scholar]
- Steffen, A.; Pawliszyn, J. Analysis of flavor volatiles using headspace solid-phase microextraction. J. Agric. Food Chem 1996, 44, 2187–2193. [Google Scholar]
- Goff, S.A.; Klee, H.J. Plant volatile compounds: sensory cues for health and nutritional value? Science 2006, 311, 815–819. [Google Scholar]
- Gang, D.R. Evolution of flavors and scents. Annu. Rev. Plant Biol 2005, 56, 301–325. [Google Scholar]
- Babazadeh Darjazi, B.; Rustaiyan, A.; Talaei, A.; Khalighi, A.; Larijani, K.; Golein, B.; Hayatbakhsh, E.; Taghizad, R. The effects of rootstock on the volatile flavour components of page mandarin [(C. reticulata var Dancy× C. paradisi var Dancan)× C. clemantina] juice and peel. Iran. J. Chem. Chem. Eng 2009, 28, 99–111. [Google Scholar]
- Fleisher, Z. Citrus petitgrain oils of Israel. Perfum. Flavor 1991, 16, 43–47. [Google Scholar]
- Crowell, P.L. Prevention and therapy of cancer by dietary monoterpenes. J. Nutr 1999, 129, 775. [Google Scholar]
- Fisher, K.; Phillips, C. Potential antimicrobial uses of essential oils in food: is citrus the answer? Trends Food Sci. Technol 2008, 19, 156–164. [Google Scholar]
- Lota, M.-L.; de Rocca Serra, D.; Tomi, F.; Jacquemond, C.; Casanova, J. Volatile components of peel and leaf oils of lemon and lime species. J. Agric. Food Chem 2002, 50, 796–805. [Google Scholar]
- Tomi, F.; Barzalona, M.; Casanova, J.; Luro, F. Chemical variability of the leaf oil of 113 hybrids from C. clementina (Commun)×C. deliciosa (Willow Leaf). Flavour Frag. J 2008, 23, 152–163. [Google Scholar]
- Hosni, K.; Zahed, N.; Chrif, R.; Abid, I.; Medfei, W.; Kallel, M.; Brahim, N.B.; Sebei, H. Composition of peel essential oils from four selected Tunisian Citrus species: evidence for the genotypic influence. Food Chem 2010, 123, 1098–1104. [Google Scholar]
- Lan-Phi, N.T.; Shimamura, T.; Ukeda, H.; Sawamura, M. Chemical and aroma profiles of yuzu (C. junos) peel oils of different cultivars. Food Chem 2009, 115, 1042–1047. [Google Scholar]
- Luro, F.; Venturini, N.; Costantino, G.; Paolini, J.; Ollitrault, P.; Costa, J. Genetic and chemical diversity of citron (Citrus. medica L.) based on nuclear and cytoplasmic markers and leaf essential oil composition. Phytochemistry 2012, 77, 186–196. [Google Scholar]
- Flamini, G.; Tebano, M.; Cioni, P.L. Volatiles emission patterns of different plant organs and pollen of Citrus limon. Anal. Chim. Acta 2007, 589, 120–124. [Google Scholar]
- Flamini, G.; Cioni, P.L. Odour gradients and patterns in volatile emission of different plant parts and developing fruits of grapefruit (Citrus paradisi L.). Food Chem 2010, 120, 984–992. [Google Scholar]
- Kasali, A.A.; Lawal, O.A.; Abanikannda, O.T.; Olaniyan, A.A.; Setzer, W.N. Citrus essential oils of nigeria part IV: volatile constituents of leaf oils of mandarins (Citrus. reticulata Blanco) from Nigeria. Record. Nat. Prod 2010, 4, 156–162. [Google Scholar]
- Fanciullino, A.-L.; Tomi, F.; Luro, F.; Desjobert, J.M.; Casanova, J. Chemical variability of peel and leaf oils of mandarins. Flavour Frag. J 2006, 21, 359–367. [Google Scholar]
- Lota, M.-L.; de Rocca Serra, D.; Tomi, F.; Joseph, C. Chemical variability of peel and leaf essential oils of mandarins from Citrus reticulata Blanco. Biochem. Syst. Ecol 2000, 28, 61–78. [Google Scholar]
- Lota, M.-L.; de Rocca Serra, D.; Tomi, F.; Bessiere, J.-M.; Casanova, J. Chemical composition of peel and leaf essential oils of Citrus medica L. and Citrus limonimedica Lush. Flavour Frag. J 1999, 14, 161–166. [Google Scholar]
- Gancel, A.-L.; Ollitrault, P.; Froelicher, Y.; Tomi, F.; Jacquemond, C.; Luro, F.; Brillouet, J.-M. Leaf Volatile Compounds of six citrus somatic allotetraploid hybrids originating from various combinations of lime, lemon, citron, sweet orange, and grapefruit. J. Agric. Food Chem 2005, 53, 2224–2230. [Google Scholar]
- Verdonk, J.C.; Ric de Vos, C.H.; Verhoeven, H.A.; Haring, M.A.; van Tunen, A.J.; Schuurink, R.C. Regulation of floral scent production in petunia revealed by targeted metabolomics. Phytochemistry 2003, 62, 997–1008. [Google Scholar]
- Augusto, F.; Valente, A.L.P.; dos Santos Tada, E.; Rivellino, S.R. Screening of Brazilian fruit aromas using solid-phase microextraction–gas chromatography–mass spectrometry. J. Chromatogr 2000, 873, 117–127. [Google Scholar]
- González-Mas, M.C.; Rambla, J.L.; Alamar, M.C.; Gutiérrez, A.; Granell, A. Comparative analysis of the volatile fraction of fruit juice from different Citrus species. PLoS One 2011, 6, e22016. [Google Scholar]
- Qiao, Y.; Xie, B.; Zhang, Y.; Zhang, Y.; Fan, G.; Yao, X.; Pan, S. Characterization of aroma active compounds in fruit juice and peel oil of jinchen sweet orange fruit (Citrus sinensis (L.) Osbeck) by GC-MS and GC-O. Molecules 2008, 13, 1333–1344. [Google Scholar]
- Barboni, T.; Luro, F.; Chiaramonti, N.; Desjobert, J.-M.; Muselli, A.; Costa, J. Volatile composition of hybrids Citrus juices by headspace solid-phase micro extraction/gas chromatography/mass spectrometry. Food Chem 2009, 116, 382–390. [Google Scholar]
- Lin, S.Y.; Roan, S.F.; Lee, C.L.; Chen, I.Z. Volatile organic components of fresh leaves as indicators of indigenous and cultivated citrus species in Taiwan. Biosci. Biotechnol. Biochem 2010, 74, 806–811. [Google Scholar]
- Tietel, Z.; Bar, E.; Lewinsohn, E.; Feldmesser, E.; Fallik, E.; Porat, R. Effects of wax coatings and postharvest storage on sensory quality and aroma volatile composition of ‘Mor’ mandarins. J. Sci. Food Agric 2010, 90, 995–1007. [Google Scholar]
- Miyazaki, T.; Plotto, A.; Goodner, K.; Gmitter, F.G. Distribution of aroma volatile compounds in tangerine hybrids and proposed inheritance. J. Sci. Food Agric 2011, 91, 449–460. [Google Scholar]
- Fanciullino, A.-L.; Gancel, A.-L.; Froelicher, Y.; Luro, F.; Ollitrault, P.; Brillouet, J.-M. Effects of nucleo-cytoplasmic interactions on leaf volatile compounds from citrus somatic diploid hybrids. J. Agric. Food Chem 2005, 53, 4517–4523. [Google Scholar]
- Perez-Cacho, P.R.; Rouseff, R.L. Fresh squeezed orange juice odor: A review. Crit. Rev. Food Sci. Nutr 2008, 48, 681–695. [Google Scholar]
- Zhang, B.; Xi, W.P.; Wei, W.W.; Shen, J.Y.; Ferguson, I.; Chen, K.S. Changes in aroma-related volatiles and gene expression during low temperature storage and subsequent shelf-life of peach fruit. Postharvest Biol. Technol 2011, 60, 7–16. [Google Scholar]
- Jabalpurwala, F.A.; Smoot, J.M.; Rouseff, R.L. A comparison of citrus blossom volatiles. Phytochemistry 2009, 70, 1428–1434. [Google Scholar]
- Pereira, P.; Cebola, M.-J.; Bernardo-Gil, M. Evolution of the yields and composition of essential oil from portuguese myrtle (Myrtus comunis L.) through the vegetative cycle. Molecules 2009, 14, 3094–3105. [Google Scholar]
- Liu, Z.-M.; Wang, H.-Y.; Liu, S.-S.; Jiang, N.-X. Volatile components of essential oil from mulberry variety “Longsang 1” leaves. Nat. Prod. Res. Dev 2012, 23, 1069–1072. [Google Scholar]
- Ortet, R.; Thomas, O.P.; Regalado, E.L.; Pino, J.A.; Filippi, J.-J.; FernáNDez, M.D. Composition and biological properties of the volatile oil of Artemisia gorgonum Webb. Chem. Biodivers 2010, 7, 1325–1332. [Google Scholar]
- Foudil-Cherif, Y.; Yassaa, N. Chiral characterization of monoterpenes present in the volatile fraction of Myrtus communis L. growing in Algeria. Int. Proc. Chem. Biol. Environ. Eng 2011, 24, 362–366. [Google Scholar]
- Casado, D.; Gemeno, C.; Avilla, J.; Riba, M. Diurnal variation of walnut tree volatiles and electrophysiological responses in Cydia pomonella (Lepidoptera:Tortricidae). Pest Manag. Sci 2008, 64, 736–747. [Google Scholar]
- Lota, M.-L.; de Rocca Serra, D.; Jacquemond, C.; Tomi, F.; Casanova, J. Chemical variability of peel and leaf essential oils of sour orange. Flavour Fragr. J 2001, 16, 89–96. [Google Scholar]
- Müller-Schwarze, D.; Thoss, V. Defense on the rocks: low monoterpenoid levels in plants on pillars without mammalian herbivores. J. Chem. Ecol 2008, 34, 1377–1381. [Google Scholar]
- Raffa, K.F.; Smalley, E.B. Interaction of pre-attack and induced monoterpene concentrations in host conifer defense against bark beetle-fungal complexes. Oecologia 1995, 102, 285–295. [Google Scholar]
- Bryant, J.P.; Reichardt, P.B.; Clausen, T. Chemically mediated interactions between woody plants and browsing mammals. J. Range Manage 1992, 45, 18–24. [Google Scholar]
- Fadel, H. Comparison studies on leaf oils of Egyptian Citrus varieties. J. Islam. Acad. Sci 1991, 4, 196–199. [Google Scholar]
- Alonzo, G.; Del Bosco, S.F.; Palazzolo, E.; Saiano, F.; Tusa, N. Citrus cybrid leaf essential oil. Flavour Frag. J 2000, 15, 91–95. [Google Scholar]
- Gancel, A.-L.; Ollitrault, P.; Froelicher, Y.; Tomi, F.; Jacquemond, C.; Luro, F.; Brillouet, J.-M. Leaf volatile compounds of seven citrus somatic tetraploid hybrids sharing willow leaf mandarin (C. deliciosa Ten.) as their common parent. J. Agric. Food Chem 2003, 51, 6006–6013. [Google Scholar]
- Gershenzon, J.; McConkey, M.E.; Croteau, R.B. Regulation of monoterpene accumulation in leaves of peppermint. Plant Physiol 2000, 122, 205–214. [Google Scholar]
- Aharoni, A.; Giri, A.P.; Deuerlein, S.; Griepink, F.; de Kogel, W.-J.; Verstappen, F.W.A.; Verhoeven, H.A.; Jongsma, M.A.; Schwab, W.; Bouwmeester, H.J. Terpenoid metabolism in wild-type and transgenic Arabidopsis plants. Plant Cell 2003, 15, 2866–2884. [Google Scholar]
- Chen, F.; Tholl, D.; D’Auria, J. C.; Farooq, A.; Pichersky, E.; Gershenzon, J. Biosynthesis and emission of terpenoid volatiles from Arabidopsis flowers. Plant Cell 2003, 15, 481–494. [Google Scholar]
- Tanaka, T. Misunderstanding with regards citrus classification and nomenclature. Bull. Univ. Osaka Pref. Ser. B 1969, 21, 13. [Google Scholar]
- Moore, G.A. Oranges and lemons: Clues to the taxonomy of Citrus from molecular markers. Trends Genet 2001, 17, 536–540. [Google Scholar]
- Lota, M.-L.; de Rocca Serra, D.; Tomi, F.; Casanova, J. Chemical variability of peel and leaf essential oils of 15 species of mandarins. Biochem. Syst. Ecol 2001, 29, 77–104. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Citrus types | Common name | Cultivars | Code | |
|---|---|---|---|---|
| 1 | C. reticulata Blanco | Mandarin | Ponkan | PK |
| 2 | C. unshiu Marc. | Mandarin | Satsuma | ST |
| 3 | C. sinensis (L.) Osbeck | Sweet orange | Liubencheng | LBC |
| 4 | C. sinensis (L.) Osbeck | Sweet orange | Qingjia | QJ |
| 5 | C. grandis (L.) Osbeck | Pomelo | Yuhuanyou | YHY |
| 6 | C. grandis (L.) Osbeck | Pomelo | Zaoxiangyou | ZXY |
| 7 | C. changshanensis Chen et. Fu (C. aurantium × C. grandis) | Citrus Hybrid | Huyou | HY |
| 8 | C. reticulate × C. sinensis | Citrus Hybrid | Hongshigan | HSG |
| 9 | C. limon (L.) Burm. | Lemon | Eureka | ERK |
| RI a | Compound name | Young leaves | Mature leaves | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FC c | PK d | ST | LBC | QJ | YHY | ZXY | HY | HSG | ERK | PK | ST | LBC | QJ | YHY | ZXY | HY | HSG | ERK | ||
| Aldehydes | ||||||||||||||||||||
| 809 | Hexanal | A1 | M | M | M | M | 1.20 | 4.30 | M | M | M | M | M | M | M | 6.86 | 7.05 | 2.67 | M | M |
| 815 | 2-hexenal | A2 | M | 1.31 | M | M | 7.15 | 13.64 | 1.74 | 1.17 | 1.20 | 2.21 | 1.37 | 2.63 | 1.25 | 14.29 | 14.60 | 7.83 | 2.23 | 1.72 |
| 911 | Heptanal | A3 | /e | / | / | / | / | / | / | / | / | / | / | / | / | / | / | T | / | / |
| 921 | 2,4-hexadienal, (E,E) *,b | A4 | T | T | T | T | / | / | T | T | T | T | T | T | / | M | / | M | T | T |
| 978 | Benzaldehyde * | A5 | / | / | / | T | / | / | / | T | T | / | / | / | / | / | / | / | / | / |
| 1058 | Melonal * | A6 | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / | T |
| 1109 | Nonanal | A7 | M | / | / | / | T | 3.00 | / | / | / | / | / | / | / | M | 2.51 | T | / | / |
| 1210 | Decanal | A8 | T | T | T | T | T | T | T | T | M | M | M | T | T | M | T | / | M | M |
| 1312 | Undecanal | A9 | / | / | / | / | / | / | / | / | T | / | T | / | / | / | / | / | T | M |
| Monoterpene hydrocarbons | ||||||||||||||||||||
| 922 | α-thujene | MH1 | 2.70 | M | 2.58 | 3.27 | M | M | M | 2.24 | T | 5.27 | M | 7.95 | 2.26 | M | M | 1.03 | 3.70 | / |
| 945 | α-pinene | MH2 | / | 1.32 | / | / | 2.56 | 1.96 | 2.24 | / | M | / | 1.33 | / | / | 2.10 | 3.11 | 2.56 | / | M |
| 962 | camphene | MH3 | T | T | T | T | M | M | T | / | / | T | T | T | T | M | M | T | T | / |
| 992 | β-pinene | MH4 | / | 5.97 | / | / | 27.91 | 21.32 | 10.18 | 11.23 | 1.04 | / | 7.29 | / | / | 21.88 | 20.65 | 11.11 | 18.05 | 1.06 |
| 993 | β-terpinene | MH5 | 14.38 | / | 15.18 | 15.36 | 4.04 | / | / | 1.65 | / | 21.83 | / | 30.55 | 13.64 | 5.70 | / | M | 3.38 | / |
| 997 | β-myrcene | MH6 | 1.88 | M | 2.01 | 2.17 | / | M | 1.01 | / | 1.22 | 3.50 | M | 4.74 | 3.46 | / | 1.41 | M | / | 1.80 |
| 1011 | α-phellandrene | MH7 | / | / | / | / | / | / | / | M | M | / | / | / | / | / | / | / | / | / |
| 1013 | 3-carene | MH8 | / | / | 4.23 | 3.31 | / | / | / | 1.88 | / | / | M | / | 8.51 | / | / | / | 5.87 | / |
| 1019 | α-terpinene | MH9 | M | M | M | M | M | M | M | M | / | 1.07 | M | 1.66 | M | M | M | M | M | / |
| 1029 | p-cymene | MH10 | / | 14.35 | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| 1030 | Limonene | MH11 | 2.99 | / | 4.34 | 4.10 | 3.64 | 4.60 | 7.91 | 4.63 | 31.71 | 4.11 | 13.71 | 13.06 | 7.29 | 6.93 | 4.47 | 7.40 | 7.78 | 38.18 |
| 1041 | Z-ocimene | MH12 | M | M | M | M | M | 0.65 | M | M | M | M | M | M | M | M | M | M | M | / |
| 1056 | E-ocimene | MH13 | 6.10 | 3.71 | 6.55 | 4.92 | 31.41 | 25.20 | 7.71 | 6.35 | 3.37 | 8.97 | 1.95 | 8.33 | 8.51 | 22.09 | 25.44 | 4.22 | 7.30 | 1.76 |
| 1068 | γ-terpinene | MH14 | 2.69 | 19.12 | 1.59 | 1.70 | M | M | 27.07 | 5.88 | M | 3.09 | 7.38 | 2.63 | M | M | M | 26.66 | 4.25 | M |
| 1084 | β-cymene | MH15 | / | T | / | / | / | / | T | / | T | / | / | / | / | M | M | / | / | T |
| 1093 | Terpinolene | MH16 | 1.17 | 3.04 | 1.59 | 1.57 | M | M | 2.12 | 1.48 | M | 1.80 | / | M | 2.16 | M | M | 1.48 | 2.53 | M |
| 1113 | 1,3,8-p-menthatriene | MH17 | / | / | / | / | T | T | / | / | T | / | T | T | T | T | M | / | / | T |
| 1131 | α-pyronene * | MH18 | / | M | / | M | / | / | / | M | M | / | M | M | / | 1.14 | / | / | M | M |
| 1143 | Allo-ocimene | MH19 | / | / | T | T | / | 1.12 | T | T | / | T | / | / | / | / | T | M | / | / |
| Oxygenated monoterpenes | ||||||||||||||||||||
| 1069 | Terpineol, cis- β * | OM1 | 1.79 | M | 2.51 | 1.91 | M | M | / | 1.22 | T | M | T | M | M | / | / | T | M | T |
| 1112 | Linalool | OM2 | 47.22 | 15.20 | 24.08 | 24.37 | 1.87 | M | 14.66 | 36.31 | 7.04 | 19.49 | 2.44 | 1.90 | 6.31 | M | M | M | 13.77 | 4.21 |
| 1117 | Thujone * | OM3 | / | / | / | / | / | / | / | T | T | / | T | M | M | / | / | / | T | T |
| 1136 | Limonene oxide, cis * | OM4 | / | / | T | T | / | / | / | T | T | / | / | / | / | 0.11 | / | / | / | / |
| 1139 | Limonene oxide, trans * | OM5 | / | / | T | T | / | / | T | / | M | / | / | M | M | / | / | / | M | M |
| 1145 | Citronellal | OM6 | T | T | M | 1.48 | M | M | T | 1.39 | 5.02 | M | 1.98 | 5.45 | M | 2.09 | M | T | 4.39 | 18.42 |
| 1148 | Isopulegol | OM7 | / | / | / | / | / | T | / | / | / | T | / | / | / | / | 1.24 | / | / | M |
| 1156 | Pinocarvone * | OM8 | / | / | / | / | / | T | T | / | / | T | / | / | T | T | M | 1.31 | / | / |
| 1161 | Terpinen-4-ol | OM9 | M | M | M | / | M | M | M | M | / | M | / | M | / | / | / | / | M | / |
| 1177 | Umbellulone * | OM10 | T | / | / | M | / | / | / | T | / | / | / | / | M | / | M | / | / | / |
| 1188 | p-cymen-8-ol * | OM11 | T | M | / | / | / | / | / | / | / | T | 1.41 | / | / | T | / | / | / | / |
| 1192 | α-terpineol | OM12 | 1.65 | 2.23 | 3.62 | 2.68 | M | 3.34 | M | 1.98 | 1.36 | T | M | T | M | / | / | / | T | M |
| 1198 | p-menth-8-en-2-one * | OM13 | / | / | T | T | / | / | M | / | T | / | / | T | / | / | / | / | T | T |
| 1207 | Carveol * | OM14 | / | / | / | T | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| 1216 | p-menth-1-en-9-al * | OM15 | / | / | / | T | / | / | / | T | M | / | / | / | T | / | / | / | T | M |
| 1220 | cis-carveol * | OM16 | T | T | / | / | / | / | T | / | T | / | / | / | / | / | / | / | T | / |
| 1231 | cis-geraniol | OM17 | T | T | M | M | / | M | T | M | M | T | 1.22 | M | M | / | T | / | 4.60 | M |
| 1237 | β-citronellol | OM18 | / | / | T | M | / | / | / | M | / | / | / | M | M | / | / | / | / | M |
| 1239 | methyl thymyl ether | OM19 | 2.01 | / | / | / | / | / | / | / | / | 5.51 | / | / | / | / | / | / | / | / |
| 1243 | β-citral | OM20 | T | T | 6.52 | 5.99 | 2.14 | 4.10 | T | 3.28 | 7.61 | / | 15.10 | 3.15 | 9.57 | M | 1.37 | / | M | 4.03 |
| 1252 | p-menth-1-en-3-one | OM21 | / | T | M | M | / | / | T | M | T | / | T | T | T | / | / | / | T | T |
| 1258 | trans-geraniol | OM22 | T | T | M | M | T | T | T | M | M | T | M | T | M | T | T | / | T | M |
| 1268 | α-citral | OM23 | T | T | 8.93 | 8.92 | 2.93 | 5.42 | M | 4.81 | 11.23 | T | 19.65 | 4.06 | 12.47 | 1.02 | 1.32 | / | M | 5.32 |
| 1285 | α-thujenal * | OM24 | T | / | / | T | / | / | / | T | / | T | / | / | / | / | / | / | T | / |
| 1300 | p-Mentha-1(7),8(10)-dien-9-ol | OM25 | M | / | T | T | / | / | T | 1.37 | M | T | T | / | T | / | / | T | M | T |
| 1305 | Carvacrol | OM26 | T | / | / | / | / | / | T | T | / | / | / | / | / | / | / | T | / | / |
| 1755 | E,E-farnesal | OM27 | / | / | / | T | / | / | T | / | T | T | / | T | T | / | / | / | / | T |
| Sesquiterpene hydrocarbons | ||||||||||||||||||||
| 1335 | Ylangene * | SH1 | / | / | T | / | / | / | / | / | M | / | / | M | / | 1.92 | / | / | / | M |
| 1336 | δ-elemene | SH2 | 1.64 | M | / | / | / | / | 1.34 | / | / | 3.79 | / | / | T | / | / | 2.37 | / | / |
| 1347 | α-cubebene | SH3 | T | M | T | T | M | / | M | T | T | T | T | T | / | / | / | M | T | T |
| 1367 | Copaene | SH4 | T | / | / | / | / | / | T | T | / | M | / | / | / | / | / | T | / | / |
| 1379 | β-bourbonene | SH5 | / | T | / | / | T | / | / | / | / | T | / | / | / | T | / | T | / | / |
| 1393 | β-elemene | SH6 | / | 12.93 | 4.88 | 6.42 | M | M | 7.04 | 2.79 | 8.76 | / | 7.80 | M | 6.41 | M | 1.37 | 10.63 | 6.31 | / |
| 1409 | Zingiberene * | SH7 | M | / | M | M | / | / | T | T | T | M | / | M | T | / | / | M | M | T |
| 1415 | Caryophyllene | SH8 | 1.07 | 6.50 | 1.65 | 2.27 | 5.26 | 2.54 | M | 1.90 | 5.23 | 1.79 | 4.76 | 1.48 | 1.83 | 2.74 | T | M | 2.13 | 4.43 |
| 1419 | (Z)-α-bergamotene * | SH9 | / | / | T | T | / | / | 4.13 | T | T | / | T | / | T | M | / | 5.72 | / | T |
| 1423 | β-cubebene | SH10 | M | T | T | T | T | T | T | T | M | T | M | M | T | M | M | T | ||
| 1426 | α-elemene * | SH11 | T | / | / | / | / | / | / | T | / | / | / | T | / | / | / | / | T | / |
| 1431 | Aromadendrene | SH12 | / | M | T | T | / | M | M | T | / | / | T | M | T | M | M | M | T | / |
| 1433 | γ-elemene* | SH13 | 2.66 | T | / | / | 1.93 | 1.06 | 2.39 | / | T | 5.28 | / | / | / | 2.05 | M | 3.20 | M | T |
| 1435 | α-guaiene * | SH14 | T | T | / | / | / | / | T | / | 1.43 | T | M | T | / | / | / | T | / | M |
| 1441 | α-himachalene * | SH15 | / | / | T | T | M | / | / | / | / | / | / | / | / | / | / | T | / | / |
| 1450 | α-caryophyllene | SH16 | M | 1.67 | M | M | M | M | 1.14 | M | 1.19 | M | M | M | M | M | M | 1.64 | M | M |
| 1452 | Allo-aromadendrene | SH17 | T | / | / | / | / | / | / | / | / | T | T | T | / | T | T | / | / | / |
| 1456 | Bicyclosesquiphellandrene * | SH18 | / | T | M | / | T | / | M | T | / | M | / | M | T | T | / | M | M | / |
| 1457 | β-santalene * | SH19 | / | / | / | / | / | / | / | / | M | / | T | / | / | / | / | / | / | T |
| 1462 | β-farnesene | SH20 | / | / | 2.50 | 1.41 | / | / | 2.01 | 2.21 | 1.29 | / | M | 3.71 | M | / | / | 2.62 | 2.54 | M |
| 1470 | γ-selinene * | SH21 | 2.15 | M | / | / | / | / | / | / | T | 2.90 | T | T | / | / | / | / | / | T |
| 1475 | Germacrene D | SH22 | M | / | T | T | M | M | / | T | M | M | T | / | T | M | T | 1.75 | T | M |
| 1478 | β-selinene | SH23 | T | M | M | M | T | / | M | M | M | T | M | T | T | / | / | M | M | T |
| 1481 | γ-himachalene | SH24 | / | T | T | T | / | / | T | T | T | / | T | / | T | / | / | T | T | T |
| 1488 | α-selinene | SH25 | / | M | M | M | / | / | M | M | M | / | M | 1.11 | M | / | / | M | M | M |
| 1493 | δ-guaiene * | SH26 | / | / | / | / | T | / | / | / | / | / | / | / | / | T | T | / | / | / |
| 1496 | α-muurolene | SH27 | T | T | T | / | T | T | T | T | / | M | T | T | / | M | M | M | T | / |
| 1497 | β-gurjunene | SH28 | T | / | / | T | / | / | / | / | T | T | / | / | T | / | / | / | / | T |
| 1500 | α-bulnesene * | SH29 | T | T | T | T | / | / | T | / | / | T | / | / | / | / | / | T | / | T |
| 1501 | β-cadinene | SH30 | / | / | T | / | / | / | M | / | / | / | T | / | / | / | / | T | / | / |
| 1502 | cis-α-bisabolene * | SH31 | / | M | / | / | / | / | / | / | M | / | / | / | / | / | / | / | / | T |
| 1503 | α-amorphene | SH32 | / | / | / | / | M | M | / | / | / | / | / | / | / | M | 3.98 | / | / | / |
| 1504 | γ-muurolene | SH33 | M | ND | T | T | / | / | T | T | / | M | M | T | T | / | / | M | T | / |
| 1505 | β-bisabolene | SH34 | / | / | / | / | / | / | / | / | 2.21 | / | / | / | / | / | / | / | / | 1.21 |
| 1506 | α-farnesene* | SH35 | M | 4.46 | M | M | M | / | / | 1.26 | / | M | M | M | M | T | T | / | M | / |
| 1509 | δ-cadinene | SH36 | M | M | T | T | M | / | / | T | M | M | M | M | T | / | M | M | M | T |
| 1510 | β-sesquiphellandrene | SH37 | M | / | M | M | / | / | M | M | T | M | / | M | T | / | / | M | M | / |
| 1512 | 8-isopropenyl-1,5-dimethyl-cyclodeca-1,5-diene * | SH38 | / | / | / | / | T | T | M | / | / | / | / | / | / | T | T | T | / | / |
| 1514 | Valencene | SH39 | / | / | / | / | / | / | T | / | T | / | / | / | / | / | / | T | / | T |
| 1515 | Eudesma-3,7(11)-diene * | SH40 | T | T | / | T | T | / | T | / | / | M | / | T | T | T | / | M | / | / |
| 1526 | α-panasinsene * | SH41 | / | / | / | / | / | / | / | / | / | T | / | / | / | / | / | / | / | / |
| 1531 | (E,Z)-α-farnesene | SH42 | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| 1643 | Cadalene * | SH43 | / | / | / | T | / | / | T | / | T | / | / | / | T | / | / | / | / | / |
| Oxygenated sesquiterpenes | ||||||||||||||||||||
| 1530 | Nerolidol 2 | OS1 | T | / | / | / | / | M | T | T | T | T | / | / | / | / | T | / | T | T |
| 1531 | Germacrene D-4-ol | OS2 | T | T | / | / | M | M | / | / | / | T | / | T | / | / | M | / | / | / |
| 1587 | Caryophyllene oxide | OS3 | T | M | / | T | T | T | T | T | T | T | T | T | T | T | / | / | T | T |
| 1592 | α-humulene oxide * | OS4 | / | / | / | / | / | / | / | / | T | / | / | / | / | / | / | / | / | / |
| 1614 | Cubenol * | OS5 | T | T | / | / | T | / | T | / | T | T | / | / | / | / | / | / | / | / |
| 1626 | Spathulenol | OS6 | T | / | / | / | / | / | T | / | / | T | / | / | / | / | / | T | / | / |
| 1657 | β-eudesmol | OS7 | T | / | / | / | / | M | T | / | / | T | / | / | / | / | T | / | T | / |
| 1765 | α-sinensal | OS8 | M | / | T | T | / | / | / | T | / | T | / | / | T | / | / | / | / | / |
| Ketones | ||||||||||||||||||||
| 1399 | cis-jasmone * | K1 | / | T | / | / | / | / | / | T | T | / | / | / | / | / | / | / | T | / |
| 1484 | β-ionone * | K2 | / | / | T | T | T | T | / | T | / | / | / | / | / | / | / | / | T | T |
| Esters | ||||||||||||||||||||
| 1219 | Acetic acid, octyl ester * | E1 | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / | / | T | / |
| 1328 | Methyl geranate | E2 | / | / | T | T | / | / | / | T | / | / | T | T | T | / | / | / | M | / |
| 1330 | Myrtenyl acetate * | E3 | T | / | / | / | / | / | / | / | / | T | / | / | T | / | / | / | / | / |
| 1360 | Citronellol acetate | E4 | T | / | / | T | / | / | T | / | T | T | M | M | 2.66 | M | / | T | M | 1.24 |
| 1370 | Nerol acetate | E5 | / | / | T | M | / | M | / | / | M | / | 3.93 | M | 4.49 | M | 2.39 | / | 1.63 | 2.21 |
| 1398 | Geranyl acetate | E6 | M | / | T | T | / | / | / | / | / | 1.34 | / | / | / | / | / | / | / | 5.25 |
| Miscellaneous | ||||||||||||||||||||
| 877 | Styrene * | Ah1 | / | T | T | T | / | / | T | / | T | / | / | / | / | M | T | / | T | T |
| 1176 | Naphthalene * | Ah2 | / | / | / | / | / | / | / | / | M | / | / | / | / | T | T | T | / | M |
| 966 | Phenol * | P1 | / | / | / | / | / | / | / | / | / | / | / | / | / | / | 1.56 | / | / | / |
| 1298 | p-thymol * | P2 | / | M | / | / | / | / | T | / | / | / | T | / | / | / | / | / | / | / |
| 1076 | 1-octanol | Ac1 | / | / | / | / | / | M | / | / | / | / | / | / | / | T | / | T | / | / |
| 1292 | Phenyl-but-3-en-1-ol * | Ac2 | T | / | T | T | / | / | / | T | / | T | / | T | / | / | / | / | / | / |
| 1172 | Vinylcyclohexane * | U1 | / | / | / | M | / | / | / | M | M | / | / | / | M | / | / | / | T | / |
| 1387 | 1-decen-3-yne * | U2 | / | / | / | / | / | / | / | T | / | / | / | / | T | / | / | / | T | / |
| 1286 | Indole | ND | / | T | / | / | / | / | T | / | M | / | / | / | / | / | / | / | / | / |
| Compounds | FC a | PK b | ST | LBC | QJ | YHY | ZXY | HY | HSG | ERK | Mean rank |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Linalool | OM2 | 47.22 1st | 15.20 2nd | 24.08 1st | 24.37 1st | 1.87 11th | 0.15 33th | 14.66 2nd | 36.31 1st | 7.04 5th | 18.99 1st |
| E-ocimene | MH13 | 6.10 3rd | 3.71 8th | 6.5 4th | 4.92 6th | 31.41 1st | 25.20 1st | 7.71 5th | 6.35 3th | 3.37 8th | 10.59 2nd |
| β-pinene | MH4 | /c | 5.97 6th | / | / | 27.91 2nd | 21.32 2nd | 10.18 3rd | 11.23 2nd | 1.04 16th | 8.63 3rd |
| Limonene | MH11 | 2.99 4th | / | 4.34 7th | 4.10 7th | 3.64 6th | 4.60 5th | 7.91 4th | 4.63 6th | 31.71 1st | 7.10 4th |
| γ-terpinene | MH14 | 2.69 6th | 19.12 1st | 1.59 15th | 1.70 14th | 0.37 20th | 0.55 18th | 27.07 1st | 5.88 4th | 0.73 18th | 6.63 5th |
| β-terpinene | MH5 | 14.38 2nd | / | 15.18 2nd | 15.36 2nd | 4.04 5th | / | / | 1.65 14th | / | 5.62 6th |
| β-elemene | SH6 | / | 12.93 4th | 4.88 6th | 6.42 4th | 0.32 22th | 0.15 5th | 7.04 6th | 2.79 8th | 8.76 3rd | 4.81 7th |
| α-Citral | OM23 | 0.04 45th | 0.02 50th | 8.93 3rd | 8.92 3rd | 2.93 7th | 5.42 4th | 0.11 32th | 4.81 5th | 11.23 2nd | 4.71 8th |
| β-Citral | OM20 | 0.03 53th | 0.01 55th | 6.52 5th | 5.99 5th | 2.14 9th | 4.10 7th | 0.08 38th | 3.28 7th | 7.61 4th | 3.31 9th |
| 2-hexenal | A2 | 0.67 19th | 1.31 13th | 0.92 17th | 0.83 19th | 7.15 3rd | 13.64 3rd | 1.74 12th | 1.17 20th | 1.20 14th | 3.18 10th |
| Caryophyllene | SH8 | 1.07 15th | 6.50 5th | 1.65 14th | 2.27 11th | 5.26 4th | 2.54 10th | 0.12 31th | 1.90 12th | 5.23 6th | 2.95 11th |
| α-terpineol | OM12 | 1.65 12th | 2.23 10th | 3.62 9th | 2.68 10th | 0.83 13th | 3.34 8th | 0.26 23th | 1.98 11th | 1.36 9th | 2.00 12th |
| p-cymene | MH10 | / | 14.35 3rd | / | / | / | / | / | / | / | 1.59 13th |
| α-thujene | MH1 | 2.70 5th | 0.44 20th | 2.58 10th | 3.27 9th | 0.48 18th | 0.73 15th | 0.76 17th | 2.24 9th | 0.05 48th | 1.47 14th |
| Terpinolene | MH16 | 1.17 14th | 3.04 9th | 1.59 16th | 1.57 15th | 0.27 25th | 0.39 22th | 2.12 10th | 1.48 15th | 0.21 16th | 1.31 15th |
| Citronellal | OM6 | 0.07 14th | 0.03 14th | 0.85 17th | 1.48 15th | 0.21 16th | 0.47 12th | 0.04 17th | 1.39 16th | 5.02 7th | 1.06 16th |
| β-myrcene | MH6 | / | / | 2.01 13th | 2.17 12th | / | 0.47 20th | 1.01 15th | / | 1.22 13th | 1.07 17th |
| 3-carene | MH8 | / | / | 4.23 8th | 3.31 8th | / | / | / | / | / | 1.05 18th |
| β-farnesene | SH19 | / | / | 2.50 12th | 1.41 17th | / | / | 2.01 11th | / | 1.29 12th | 1.05 19th |
| α-pinene | MH2 | / | 1.32 12th | / | / | 2.56 8th | 1.96 11th | 2.24 9th | / | 0.19 33th | 0.92 20th |
| Compounds | FC a | PK b | ST | LBC | QJ | YHY | ZXY | HY | HSG | ERK | Mean rank |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Limonene | MH11 | 4.11 7th | 13.71 3rd | 13.06 2th | 7.29 6th | 6.93 4th | 4.47 5th | 7.40 5th | 7.78 3rd | 38.18 1st | 11.44 1st |
| E-ocimene | MH13 | 8.97 3rd | 1.95 11th | 8.33 3th | 8.51 5th | 22.09 1st | 25.44 1st | 4.22 7th | 7.30 4th | 1.76 10th | 9.84 2nd |
| β-pinene | MH4 | /c | 7.29 6th | / | / | 21.88 2nd | 20.65 2nd | 11.11 2nd | 18.05 1st | 1.06 14th | 8.89 3rd |
| β-terpinene | MH5 | 21.83 1st | / | 30.55 1st | 13.64 1st | 5.70 6th | / | / | 3.38 11th | / | 8.34 4th |
| Linalool | OM2 | 19.49 2nd | 2.44 9th | 1.90 12th | 6.31 8th | 0.1531th | 0.31 24th | 0.41 21th | 13.77 2nd | 4.21 6th | 5.44 5th |
| 2-hexenal | A2 | 2.21 12th | 1.37 13th | 2.63 10th | 1.25 15th | 14.29 3rd | 14.60 3rd | 7.83 4th | 2.23 14th | 1.72 11th | 5.35 6th |
| γ-terpinene | MH14 | 3.09 8th | 7.38 5th | 2.63 11th | 0.96 16th | 0.48 19th | 0.56 19th | 26.66 1st | 4.25 9th | 0.23 27th | 5.14 7th |
| α-citral | OM23 | 0.01 13th | 19.65 1st | 4.06 7th | 12.47 2nd | 1.02 13th | 1.32 14th | / | 0.86 19th | 5.32 3rd | 4.97 8th |
| β-citral | OM20 | / | 15.10 2nd | 3.15 9th | 9.57 3rd | 0.78 16th | 1.37 13th | / | 0.67 22th | / | 3.85 9th |
| β-elemene | SH6 | / | 7.80 4th | 0.23 27th | 6.41 7th | 0.85 14th | 1.37 12th | 10.63 3rd | 6.31 5th | / | 3.73 10th |
| Citronellal | OM6 | 0.13 12th | 1.98 10th | 5.45 5th | 0.82 18th | 2.09 9th | 0.18 27th | 0.02 15th | 4.39 8th | 18.42 2nd | 3.72 11th |
| α-thujene | MH1 | 5.27 6th | 0.37 24th | 7.95 4th | 2.26 12th | 0.62 18th | 0.75 18th | 1.03 17th | 3.70 10th | / | 2.44 12th |
| Hexanal | A1 | 0.84 17th | 0.29 26th | 0.82 17th | 0.25 28th | 6.86 5th | 7.05 4th | 2.67 9th | 0.87 18th | 0.23 26th | 2.21 13th |
| Caryophyllene | SH8 | 1.79 14th | 4.76 7th | 1.48 14th | 1.83 14th | 2.74 7th | 0.07 17th | 0.22 24th | 2.13 15th | 4.43 5th | 2.16 14th |
| β-myrcene | MH6 | 3.50 9th | 0.96 17th | 4.74 6th | 3.46 10th | / | 1.41 11th | 0.95 19th | / | 1.80 9th | 1.87 15th |
| 3-carene | MH8 | / | 0.96 16th | / | 8.51 4th | / | / | / | 5.876th | / | 1.7016th |
| Nerol acetate | E5 | / | 3.93 8th | 0.23 25th | 4.49 9th | 0.42 20th | 2.399th | / | 1.6316th | 2.218th | 1.7017th |
| γ-elemene | SH13 | 5.28 6th | / | / | / | 2.0510th | 0.7916th | 3.208th | 0.1730th | 0.0314th | 1.2818th |
| β-farnesene | SH19 | / | 0.12 17th | / | / | / | / | 2.6210th | 2.5412th | 0.2525th | 1.0819th |
| α-pinene | MH2 | / | 1.33 14th | / | / | 2.108th | 3.117th | 2.5611th | / | 0.3923th | 1.0620th |
Supplementary Files
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Azam, M.; Jiang, Q.; Zhang, B.; Xu, C.; Chen, K. Citrus Leaf Volatiles as Affected by Developmental Stage and Genetic Type. Int. J. Mol. Sci. 2013, 14, 17744-17766. https://doi.org/10.3390/ijms140917744
Azam M, Jiang Q, Zhang B, Xu C, Chen K. Citrus Leaf Volatiles as Affected by Developmental Stage and Genetic Type. International Journal of Molecular Sciences. 2013; 14(9):17744-17766. https://doi.org/10.3390/ijms140917744
Chicago/Turabian StyleAzam, Muhammad, Qian Jiang, Bo Zhang, Changjie Xu, and Kunsong Chen. 2013. "Citrus Leaf Volatiles as Affected by Developmental Stage and Genetic Type" International Journal of Molecular Sciences 14, no. 9: 17744-17766. https://doi.org/10.3390/ijms140917744