Ziyuglycoside II Inhibits the Growth of Human Breast Carcinoma MDA-MB-435 Cells via Cell Cycle Arrest and Induction of Apoptosis through the Mitochondria Dependent Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Ziyuglycoside II on Cell Growth
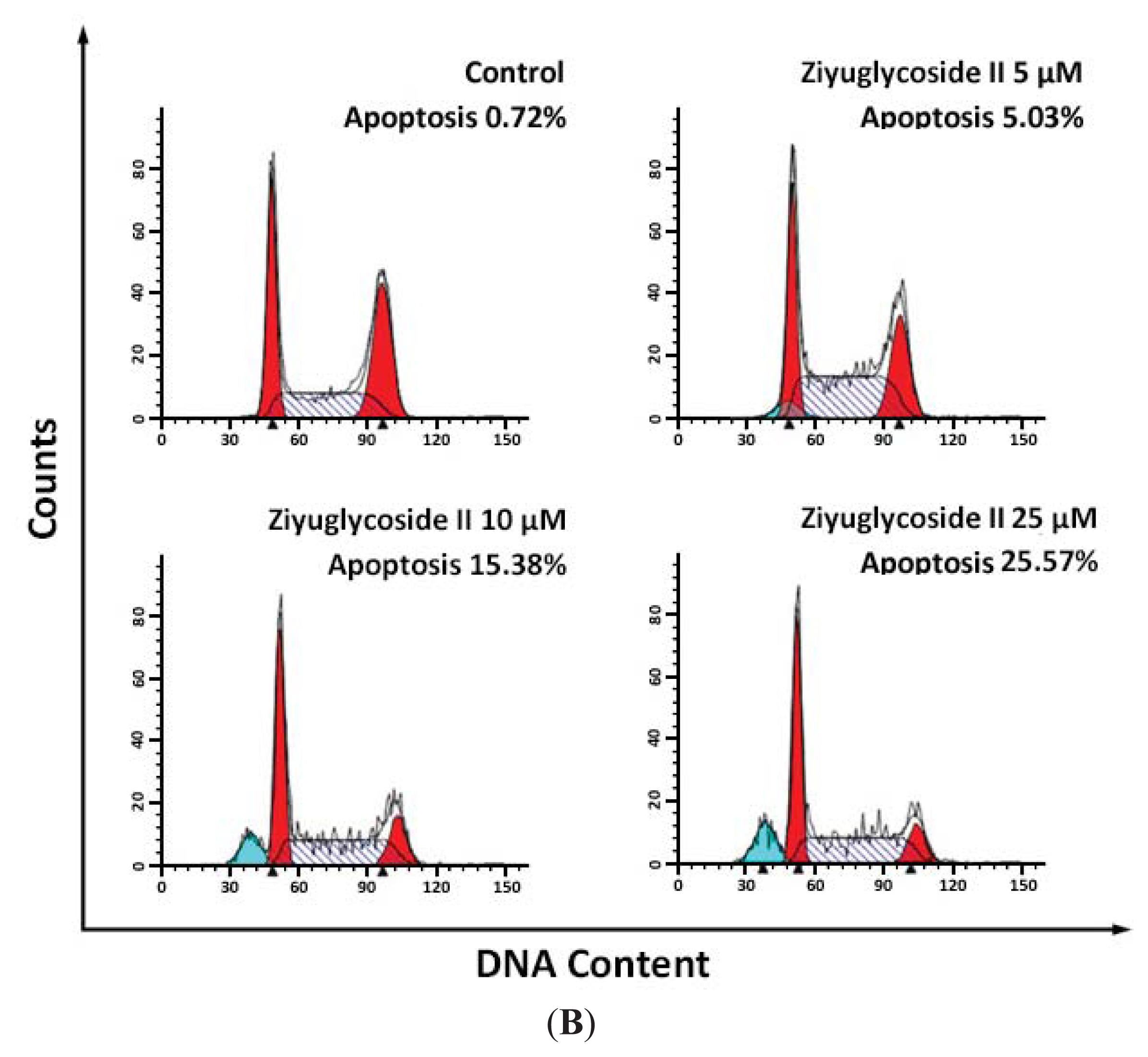
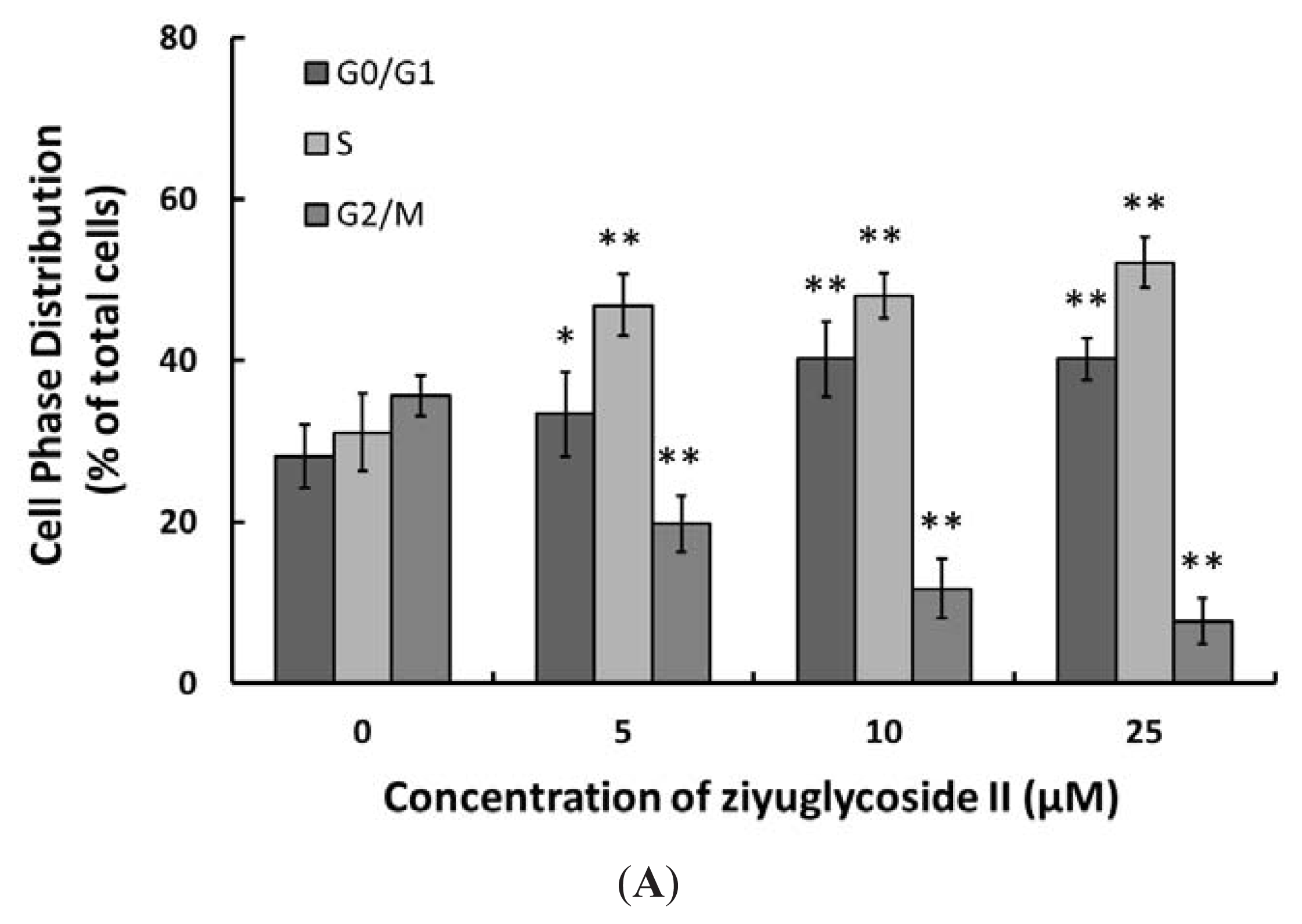
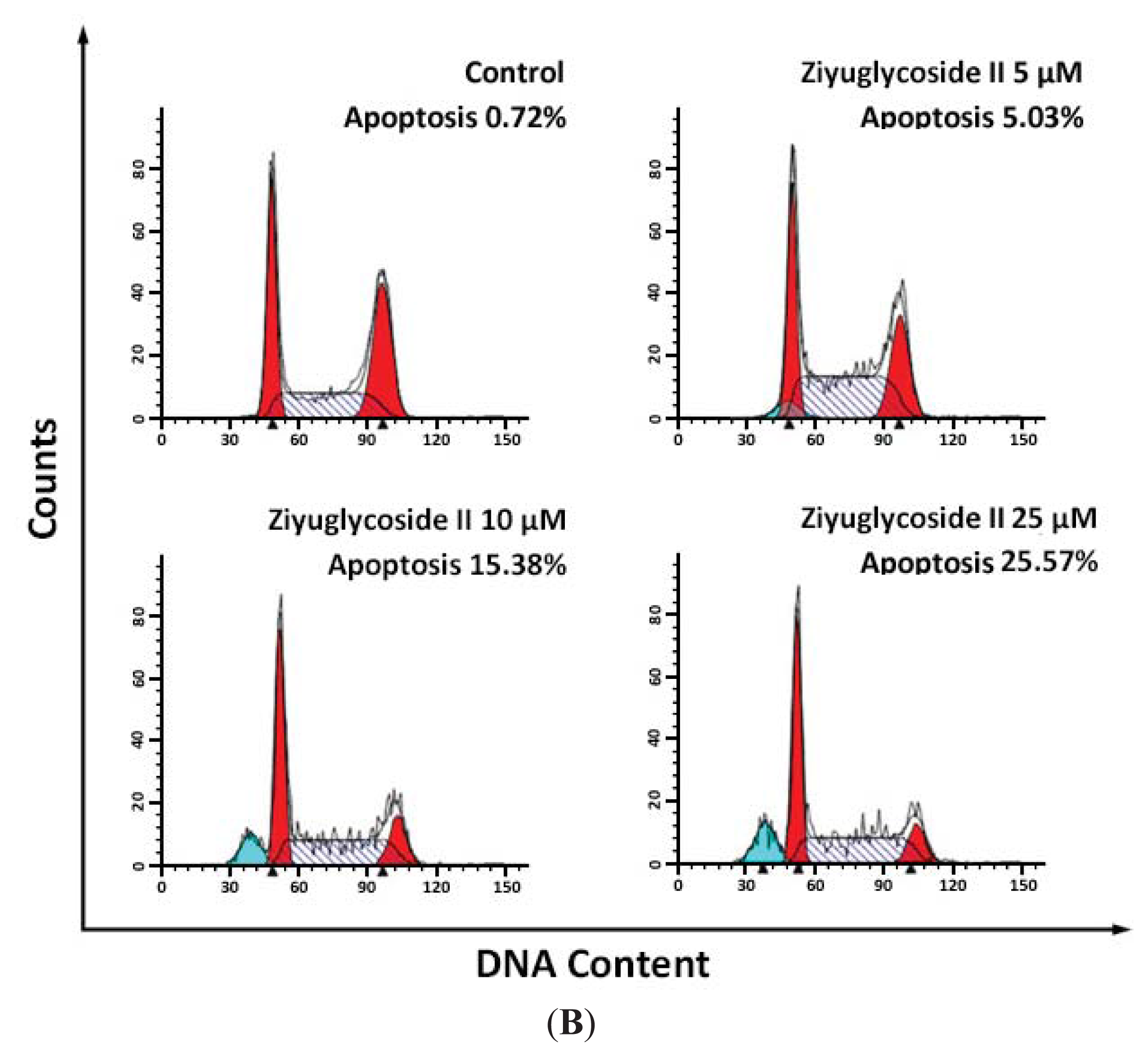
2.2. Effect of Ziyuglycoside II on Cell Cycle and Apoptosis
2.3. Effect of Ziyuglycoside II on the Generation of ROS
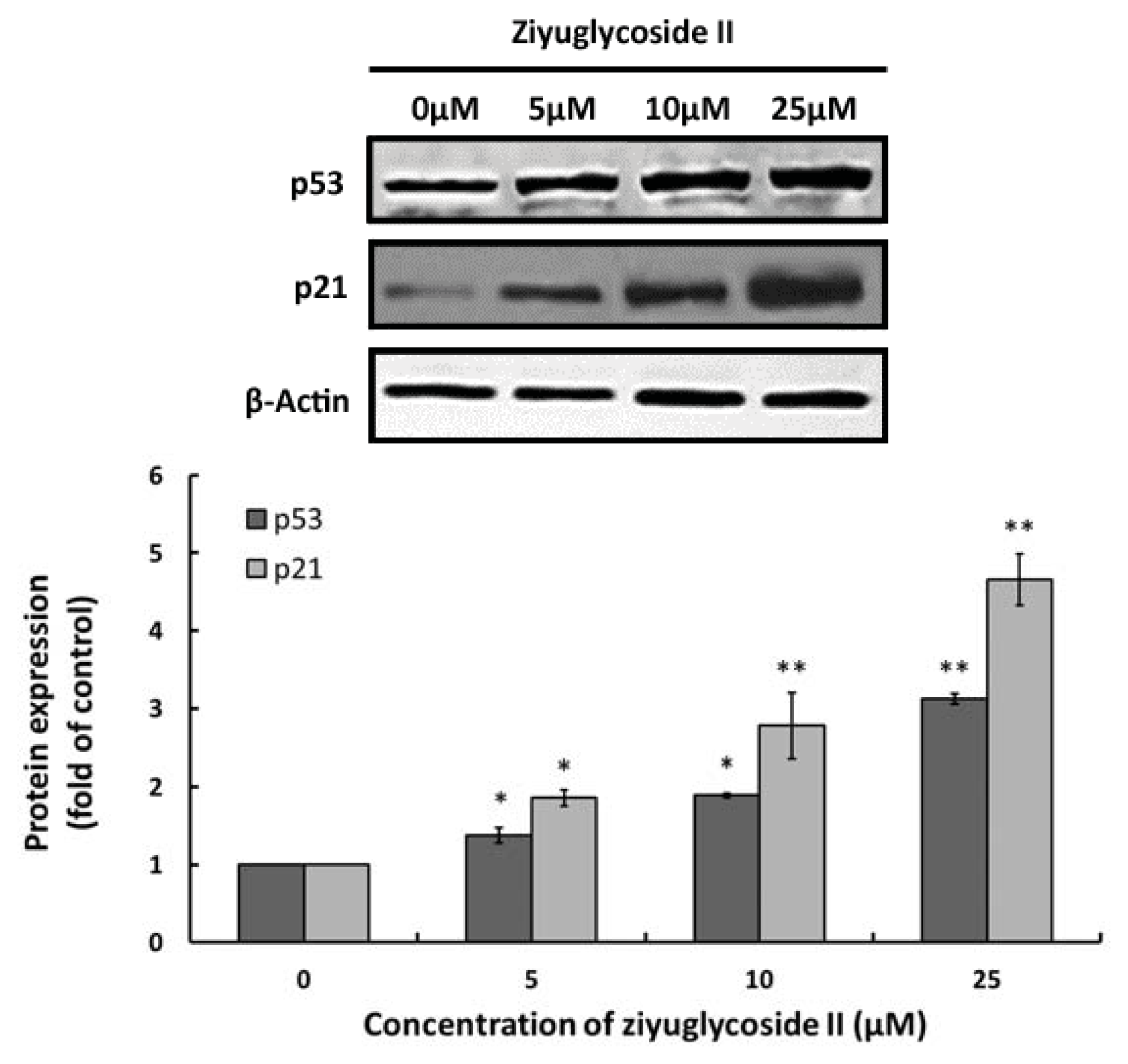
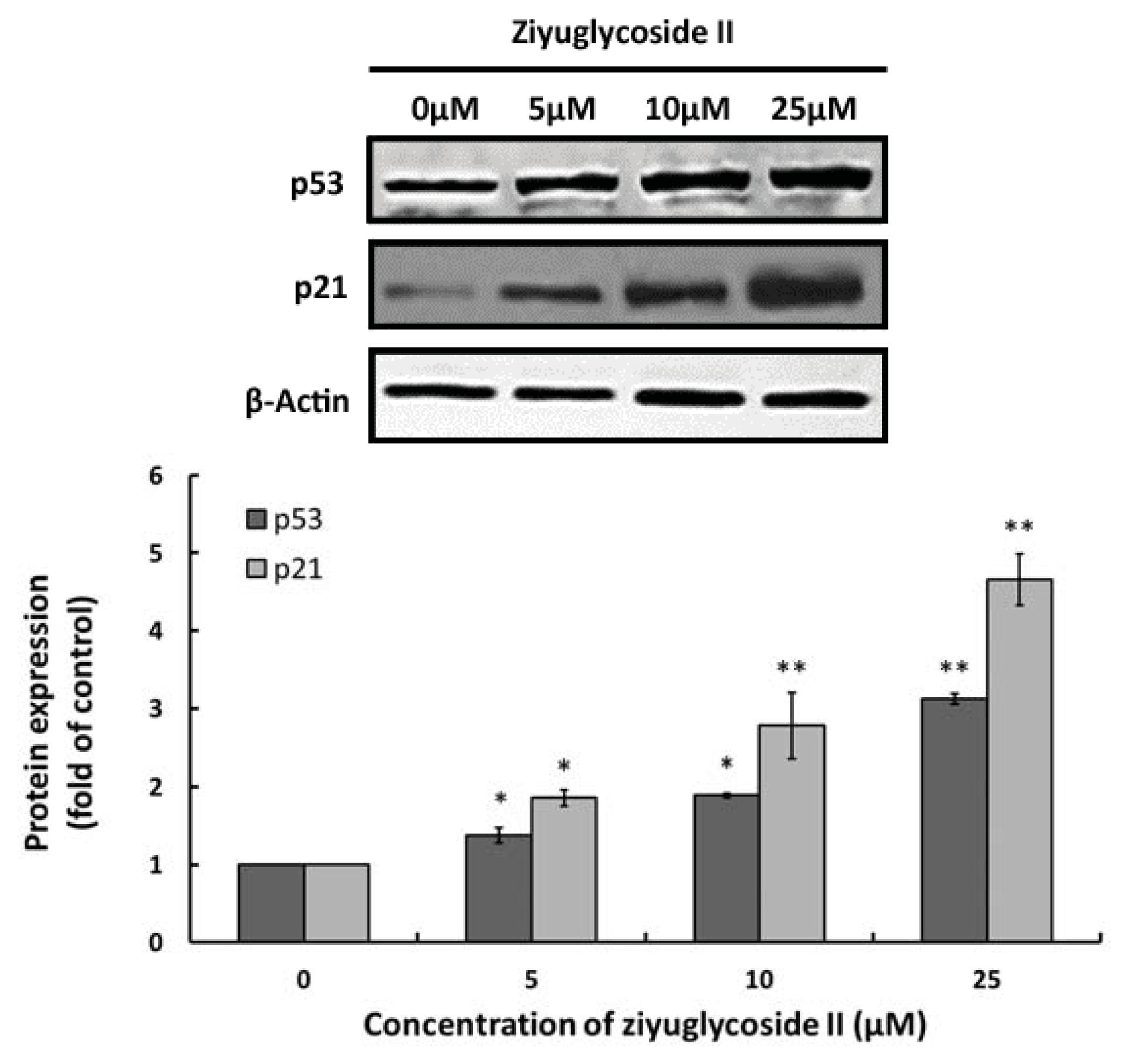
2.4. Effect of Ziyuglycoside II on Expressions of p53 and p21
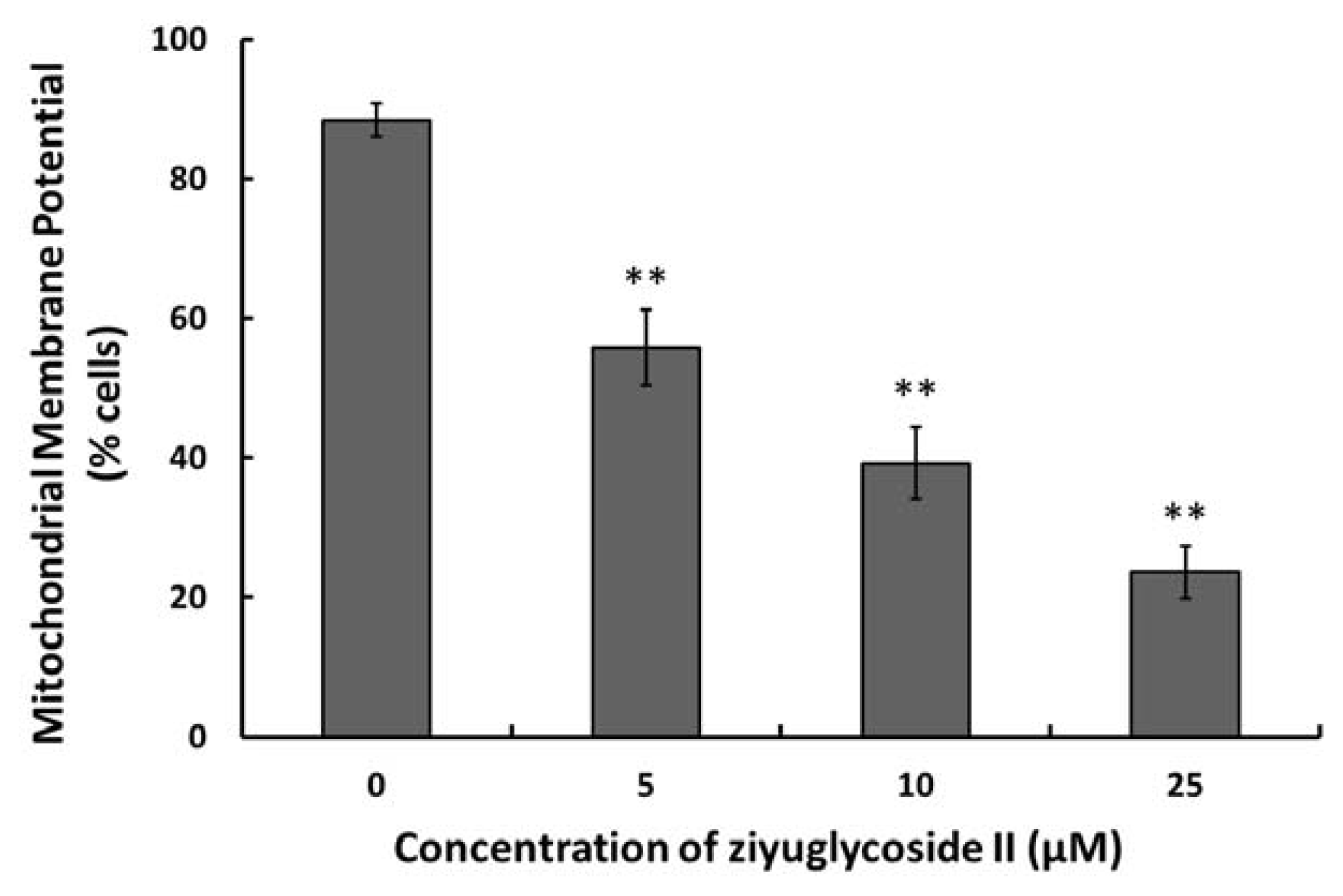
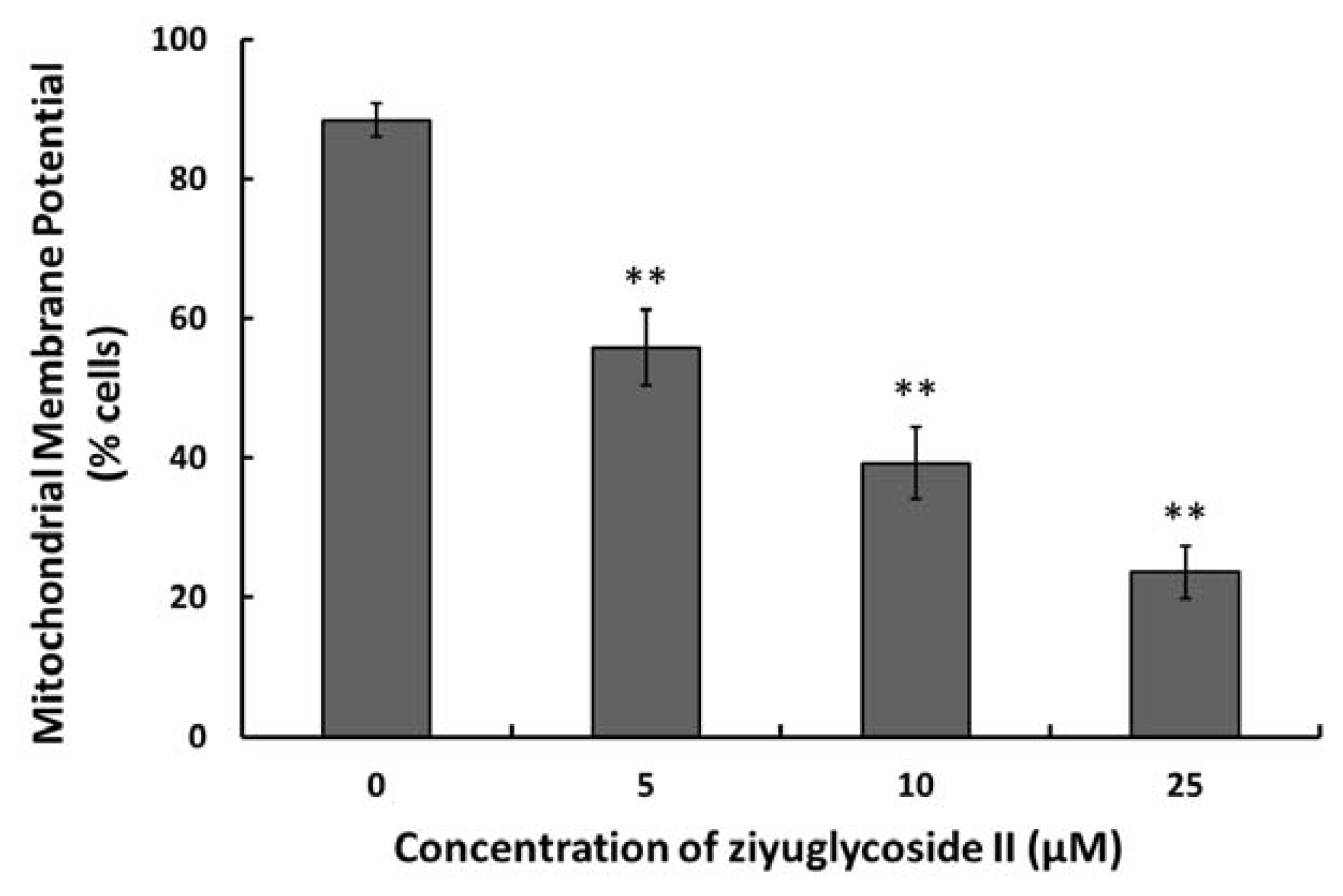
2.5. Effect of Ziyuglycoside II on the MMP
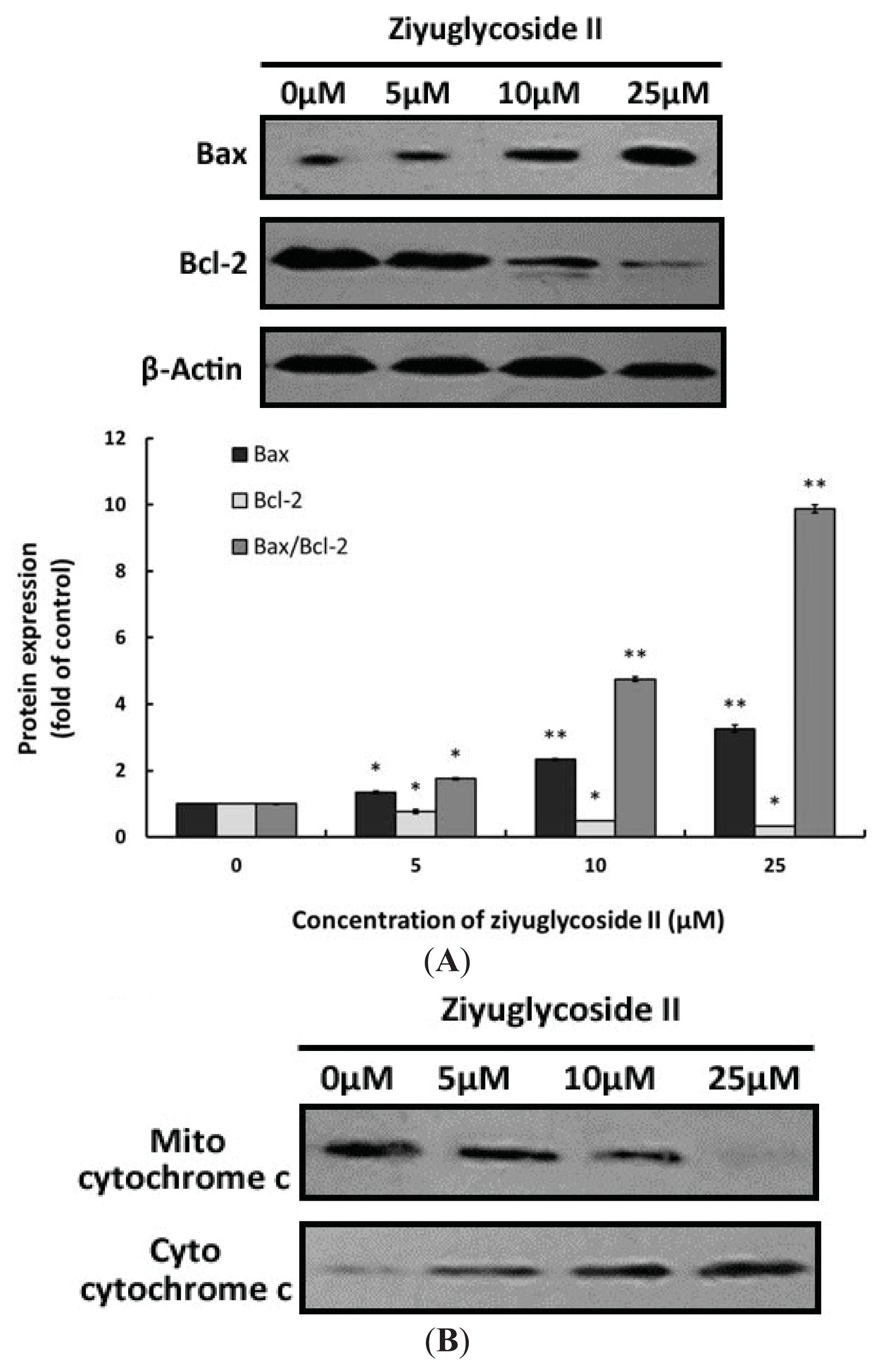
2.6. Effect of Ziyuglycoside II on the Expressions of Bax, Bcl-2 and Cytochrome C
2.7. Effect of Ziyuglycoside II on the Activity of Caspase-3 and Caspase-9
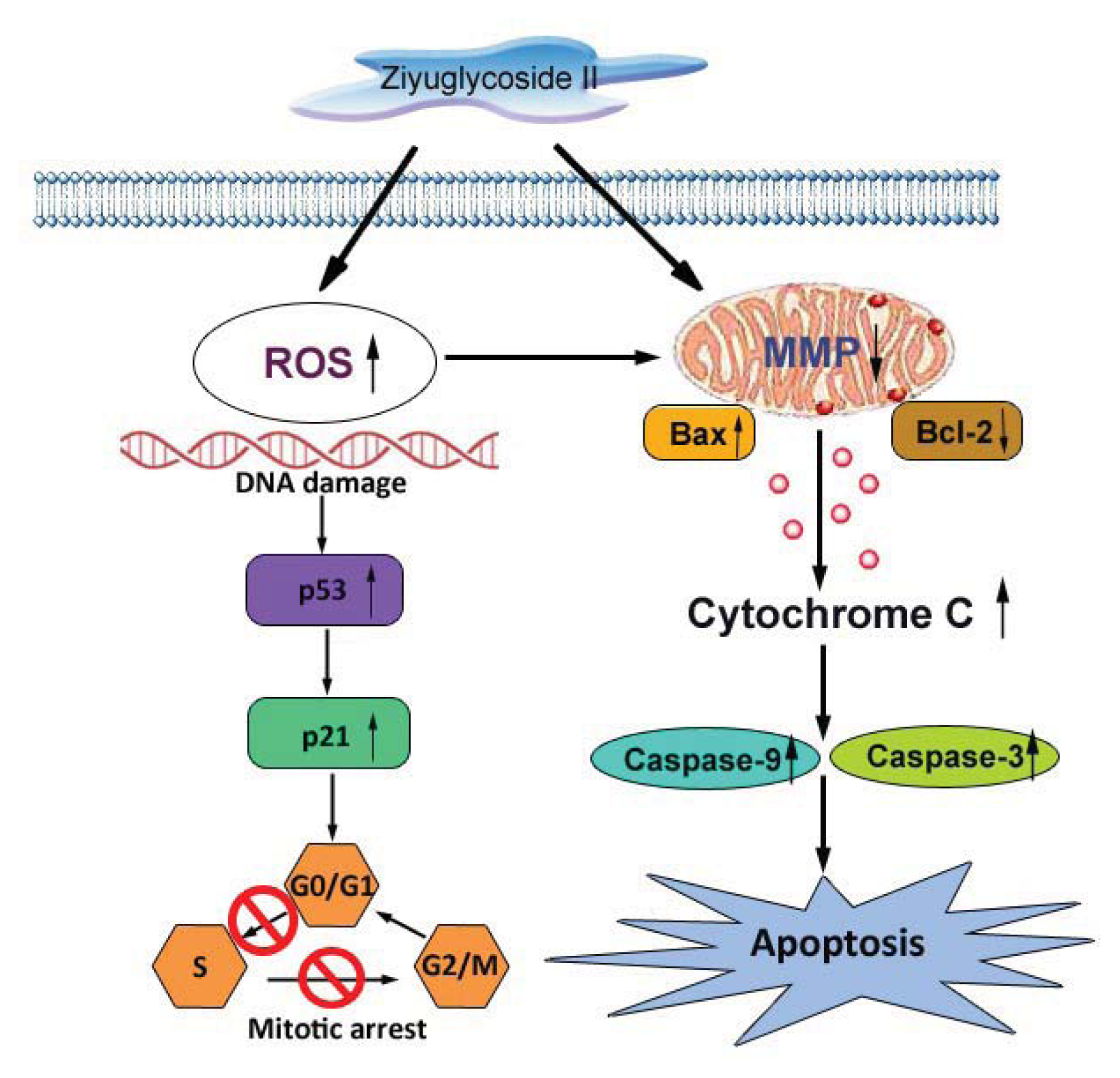
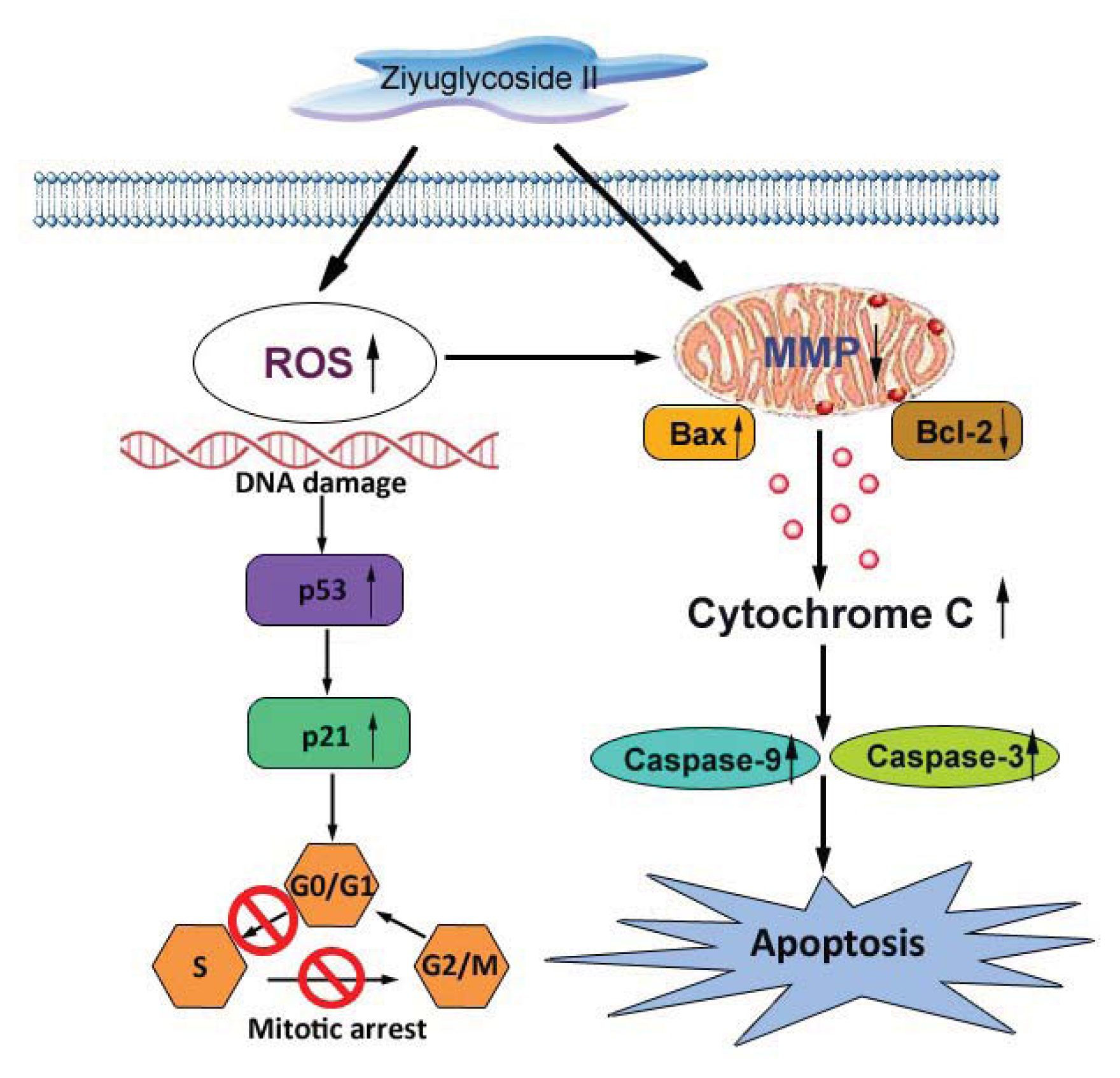
2.8. Discussion
3. Experimental Section
3.1. Materials
3.2. Cell Culture
3.3. Cell Growth Assay
3.4. Cell Cycle Distribution and Apoptosis Analysis
3.5. Intracellular Reactive Oxygen Species (ROS) Detection
3.6. Measurement of Mitochondrial Membrane Potential (MMP)
3.7. Measurement of Cytochrome C Release
3.8. Western Blotting Analysis
3.9. Caspase-3 and -9 Activity Assays
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Adami, H.O.; Signorello, L.B.; Trichopoulos, D. Towards an understanding of breast cancer etiology. Semin. Cancer Biol 1998, 8, 255–262. [Google Scholar]
- Hsu, Y.L.; Kuo, P.L.; Lin, L.T.; Lin, C.C. Asiatic acid, a triterpene, induces apoptosis and cell cycle arrest through activation of extracellular signal-regulated kinase and p38 mitogen-activated protein kinase pathways in human breast cancer cells. J. Pharmacol. Exp. Ther 2005, 313, 333–344. [Google Scholar]
- Lee, S.M.; Li, M.L.; Tse, Y.C.; Leung, S.C.; Lee, M.M.; Tsui, S.K.; Fung, K.P.; Lee, C.Y.; Waye, M.M. Paeoniae Radix, a Chinese herbal extract, inhibit hepatoma cells growth by inducing apoptosis in a p53 independent pathway. Life Sci 2002, 71, 2267–2277. [Google Scholar]
- Kim, S.; Choi, J.H.; Kim, J.B.; Nam, S.J.; Yang, J.H.; Kim, J.H.; Lee, J.E. Berberine suppresses TNF-alpha-induced MMP-9 and cell invasion through inhibition of AP-1 activity in MDA-MB-231 human breast cancer cells. Molecules 2008, 13, 2975–2985. [Google Scholar]
- Cai, Z.; Li, W.; Wang, H.; Yan, W.; Zhou, Y.; Wang, G.; Cui, J.; Wang, F. Anti-tumor and immunomodulating activities of a polysaccharide from the root of Sanguisorba officinalis L. Int. J. Biol. Macromol 2012, 51, 484–488. [Google Scholar]
- Wang, Z.; Loo, W.T.; Wang, N.; Chow, L.W.; Wang, D.; Han, F.; Zheng, X.; Chen, J.P. Effect of Sanguisorba officinalis L. on breast cancer growth and angiogenesis. Expert. Opin. Ther. Targets 2012, 16, S79–S89. [Google Scholar]
- Choi, E.S.; Kim, J.S.; Kwon, K.H.; Kim, H.S.; Cho, N.P.; Cho, S.D. Methanol extract of Sanguisorba officinalis L. with cytotoxic activity against PC3 human prostate cancer cells. Mol. Med. Rep 2012, 6, 670–674. [Google Scholar]
- Kim, Y.H.; Chung, C.B.; Kim, J.G.; Ko, K.I.; Park, S.H.; Kim, J.H.; Eom, S.Y.; Kim, Y.S.; Hwang, Y.I.; Kim, K.H. Anti-wrinkle activity of ziyuglycoside I isolated from a Sanguisorba officinalis root extract and its application as a cosmeceutical ingredient. Biosci. Biotechnol. Biochem 2008, 72, 303–311. [Google Scholar]
- Nerlich, A.G.; Bachmeier, B.E. Density-dependent lineage instability of MDA-MB-435 breast cancer cells. Oncol. Lett 2013, 5, 1370–1374. [Google Scholar]
- Chambers, A.F. MDA-MB-435 and M14 cell lines: Identical but not M14 melanoma? Cancer Res 2009, 69, 5292–5293. [Google Scholar]
- Freeman, B.A.; Crapo, J.D. Biology of disease: Free radicals and tissue injury. Lab. Investig 1982, 47, 412–426. [Google Scholar]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The mitochondrial membrane potential (Δψm) in apoptosis; An update. Apoptosis 2003, 8, 115–128. [Google Scholar]
- Scorrano, L.; Ashiya, M.; Buttle, K.; Weiler, S.; Oakes, S.A.; Mannella, C.A.; Korsmeyer, S.J. A distinct pathway remodels mitochondrial cristae and mobilizes cytochrome c during apoptosis. Dev. Cell 2002, 2, 55–67. [Google Scholar]
- Ott, M.; Robertson, J.D.; Gogvadze, V.; Zhivotovsky, B.; Orrenius, S. Cytochrome c release from mitochondria proceeds by a two-step process. Proc. Natl. Acad. Sci. USA 2002, 99, 1259–1263. [Google Scholar]
- Fan, T.-J.; Han, L.-H.; Cong, R.-S.; Liang, J. Caspase family proteases and apoptosis. Acta Biochim. Biophys. Sin 2005, 37, 719–727. [Google Scholar]
- Gottlieb, R.A. Mitochondria: Execution central. FEBS Lett 2000, 482, 6–12. [Google Scholar]
- Sun, X.M.; MacFarlane, M.; Zhuang, J.; Wolf, B.B.; Green, D.R.; Cohen, G.M. Distinct caspase cascades are initiated in receptor-mediated and chemical-induced apoptosis. J. Biol. Chem 1999, 274, 5053–5060. [Google Scholar]
- Lacroix, M. p53 and breast cancer, an update. Endocr. Relat. Cancer 2006, 13, 293–325. [Google Scholar]
- Waga, S.; Hannon, G.J.; Beach, D.; Stillman, B. The p21 inhibitor of cyclin-dependent kinases controls DNA replication by interaction with PCNA. Nature 1994, 369, 574–578. [Google Scholar]
- Mehmet, H. Caspases find a new place to hide. Nature 2000, 403, 29–30. [Google Scholar]
- Shi, Y. Mechanisms of caspase activation and inhibition during apoptosis. Mol. Cell 2002, 9, 459–470. [Google Scholar]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol 2003, 552, 335–344. [Google Scholar]
- Liu, X.; Kim, C.N.; Yang, J.; Jemmerson, R.; Wang, X. Induction of apoptotic program in cell-free extracts: Requirement for dATP and cytochrome c. Cell 1996, 86, 147–157. [Google Scholar]
- Vander Heiden, M.G.; Chandel, N.S.; Williamson, E.K.; Schumacker, P.T.; Thompson, C.B. Bcl-xL regulates the membrane potential and volume homeostasis of mitochondria. Cell 1997, 91, 627–637. [Google Scholar]
- Vander Heiden, M.G.; Chandel, N.S.; Li, X.X.; Schumacker, P.T.; Colombini, M.; Thompson, C.B. Outer mitochondrial membrane permeability can regulate coupled respiration and cell survival. Proc. Natl. Acad. Sci. USA 2000, 97, 4666–4671. [Google Scholar]
- Heiden, M.G.V.; Chandel, N.S.; Schumacker, P.T.; Thompson, C.B. Bcl-xL prevents cell death following growth factor withdrawal by facilitating mitochondrial ATP/ADP exchange. Mol. Cell 1999, 3, 159–167. [Google Scholar]
- Aoudjit, F.; Vuori, K. Integrin signaling inhibits paclitaxel-induced apoptosis in breast cancer cells. Oncogene 2001, 20, 4995–5004. [Google Scholar]
- Sunters, A.; Fernandez de Mattos, S.; Stahl, M.; Brosens, J.J.; Zoumpoulidou, G.; Saunders, C.A.; Coffer, P.J.; Medema, R.H.; Coombes, R.C.; Lam, E.W. FoxO3a transcriptional regulation of Bim controls apoptosis in paclitaxel-treated breast cancer cell lines. J. Biol. Chem 2003, 278, 49795–49805. [Google Scholar]
- Wahl, A.F.; Donaldson, K.L.; Fairchild, C.; Lee, F.Y.; Foster, S.A.; Demers, G.W.; Galloway, D.A. Loss of normal p53 function confers sensitization to Taxol by increasing G2/M arrest and apoptosis. Nat. Med 1996, 2, 72–79. [Google Scholar]
- Vogel, C.L.; Cobleigh, M.A.; Tripathy, D.; Gutheil, J.C.; Harris, L.N.; Fehrenbacher, L.; Slamon, D.J.; Murphy, M.; Novotny, W.F.; Burchmore, M.; et al. Efficacy and safety of trastuzumab as a single agent in first-line treatment of HER2-overexpressing metastatic breast cancer. J. Clin. Oncol 2002, 20, 719–726. [Google Scholar]
- Bang, Y.-J.; van Cutsem, E.; Feyereislova, A.; Chung, H.C.; Shen, L.; Sawaki, A.; Lordick, F.; Ohtsu, A.; Omuro, Y.; Satoh, T.; et al. Trastuzumab in combination with chemotherapy versus chemotherapy alone for treatment of HER2-positive advanced gastric or gastro-oesophageal junction cancer (ToGA): A phase 3, open-label, randomised controlled trial. Lancet 2010, 376, 687–697. [Google Scholar]
- Fujimoto-Ouchi, K.; Sekiguchi, F.; Yasuno, H.; Moriya, Y.; Mori, K.; Tanaka, Y. Antitumor activity of trastuzumab in combination with chemotherapy in human gastric cancer xenograft models. Cancer Chemother. Pharmacol 2007, 59, 795–805. [Google Scholar]
- Hördegen, P.; Cabaret, J.; Hertzberg, H.; Langhans, W.; Maurer, V. In vitro screening of six anthelmintic plant products against larval Haemonchus contortus with a modified methyl-thiazolyl-tetrazolium reduction assay. J. Ethnopharmacol 2006, 108, 85–89. [Google Scholar]
- Lu, H.F.; Sue, C.C.; Yu, C.S.; Chen, S.C.; Chen, G.W.; Chung, J.G. Diallyl disulfide (DADS) induced apoptosis undergo caspase-3 activity in human bladder cancer T24 cells. Food Chem. Toxicol 2004, 42, 1543–1552. [Google Scholar]
- Lin, J.-G.; Chen, G.-W.; Li, T.-M.; Chouh, S.-T.; Tan, T.-W.; Chung, J.-G. Aloe-emodin induces apoptosis in T24 human bladder cancer cells through the p53 dependent apoptotic pathway. J. Urol 2006, 175, 343–347. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Yang, J.S.; Hour, M.J.; Kuo, S.C.; Huang, L.J.; Lee, M.R. Selective induction of G2/M arrest and apoptosis in HL-60 by a potent anticancer agent, HMJ-38. Anticancer Res 2004, 24, 1769–1778. [Google Scholar]



© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhu, X.; Wang, K.; Zhang, K.; Huang, B.; Zhang, J.; Zhang, Y.; Zhu, L.; Zhou, B.; Zhou, F. Ziyuglycoside II Inhibits the Growth of Human Breast Carcinoma MDA-MB-435 Cells via Cell Cycle Arrest and Induction of Apoptosis through the Mitochondria Dependent Pathway. Int. J. Mol. Sci. 2013, 14, 18041-18055. https://doi.org/10.3390/ijms140918041
Zhu X, Wang K, Zhang K, Huang B, Zhang J, Zhang Y, Zhu L, Zhou B, Zhou F. Ziyuglycoside II Inhibits the Growth of Human Breast Carcinoma MDA-MB-435 Cells via Cell Cycle Arrest and Induction of Apoptosis through the Mitochondria Dependent Pathway. International Journal of Molecular Sciences. 2013; 14(9):18041-18055. https://doi.org/10.3390/ijms140918041
Chicago/Turabian StyleZhu, Xue, Ke Wang, Kai Zhang, Biao Huang, Jue Zhang, Yi Zhang, Lan Zhu, Bin Zhou, and Fanfan Zhou. 2013. "Ziyuglycoside II Inhibits the Growth of Human Breast Carcinoma MDA-MB-435 Cells via Cell Cycle Arrest and Induction of Apoptosis through the Mitochondria Dependent Pathway" International Journal of Molecular Sciences 14, no. 9: 18041-18055. https://doi.org/10.3390/ijms140918041