Molecular Cloning and Functional Analysis of the Duck TLR4 Gene

Abstract
:1. Introduction
2. Results
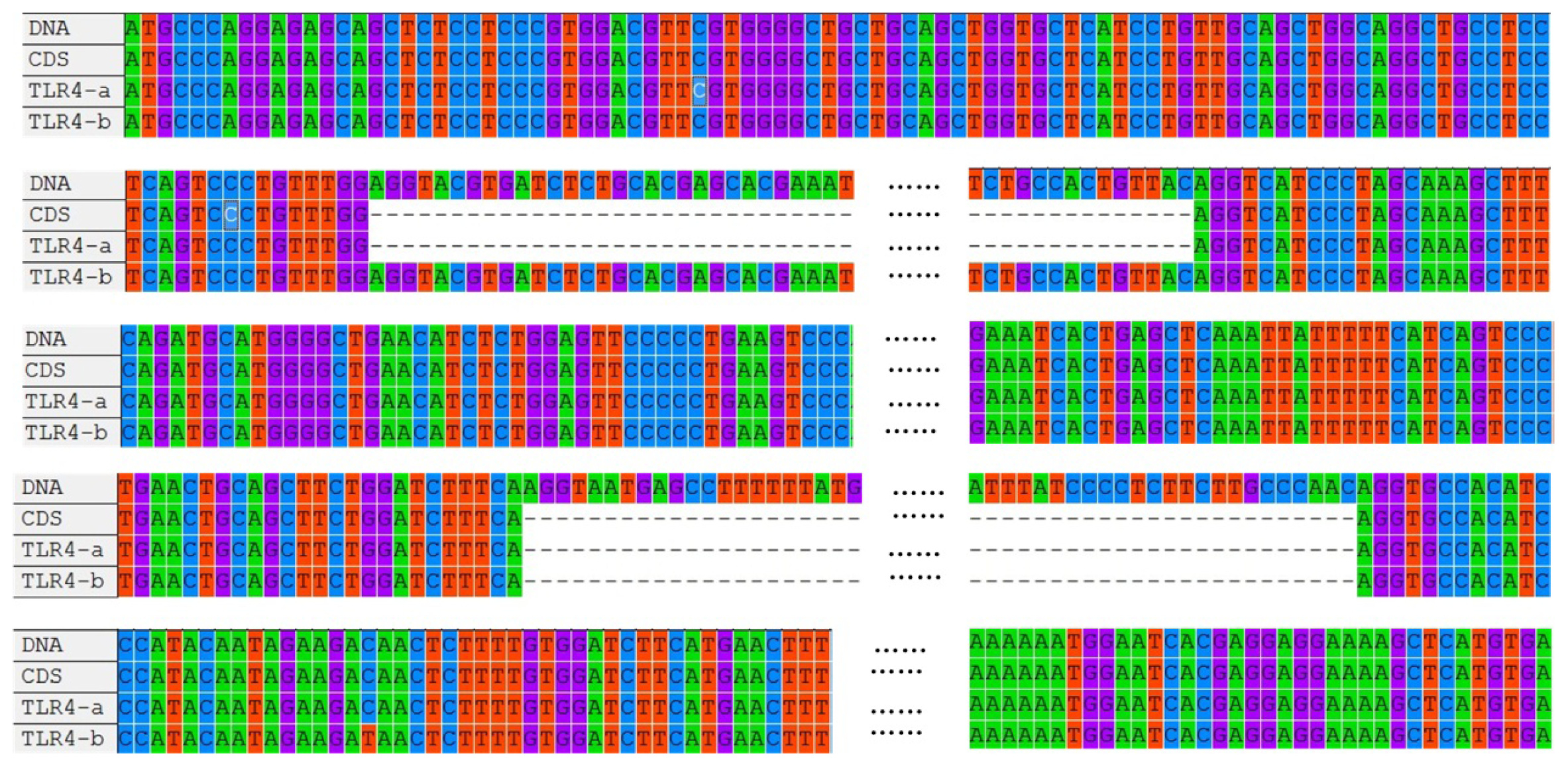
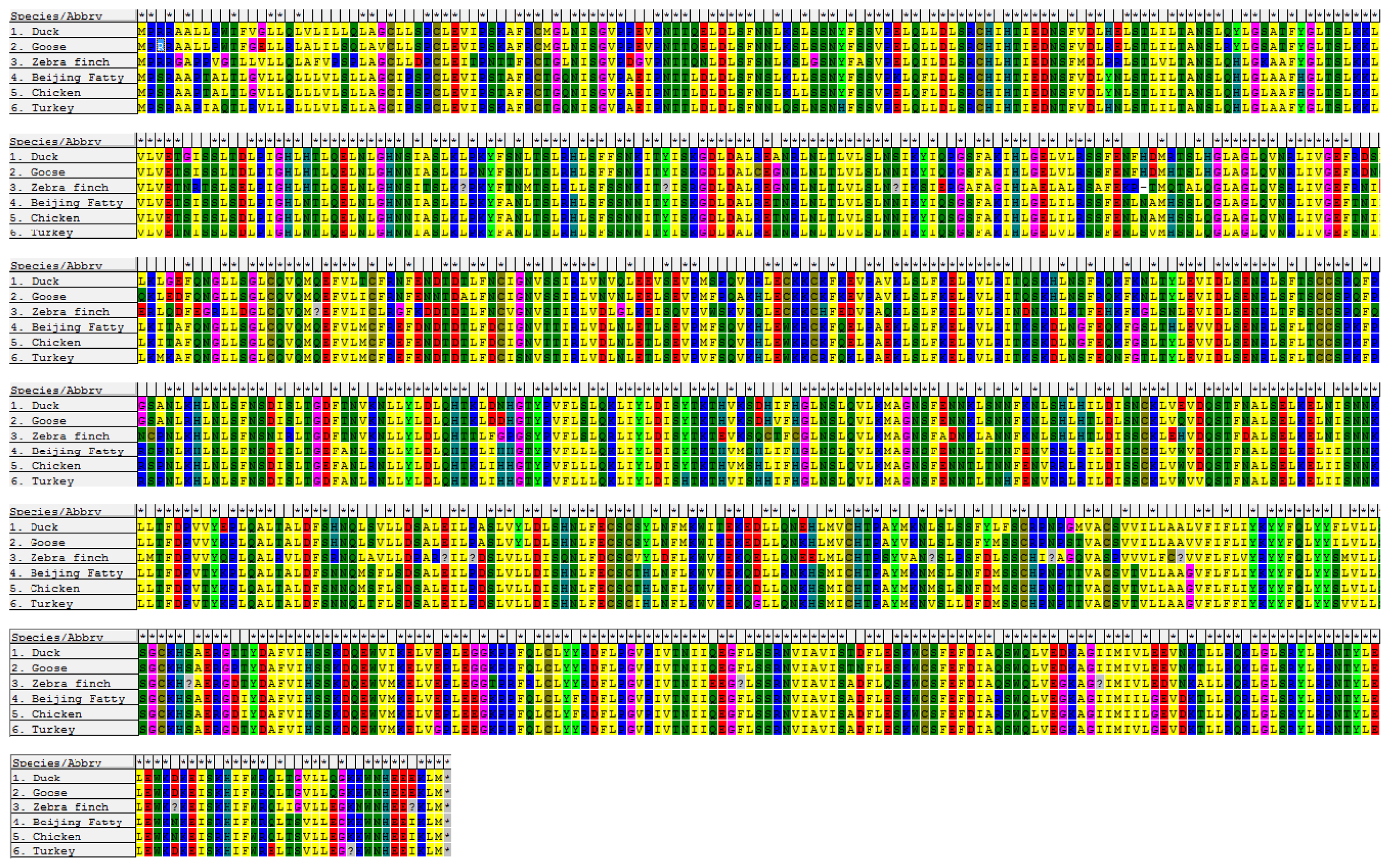
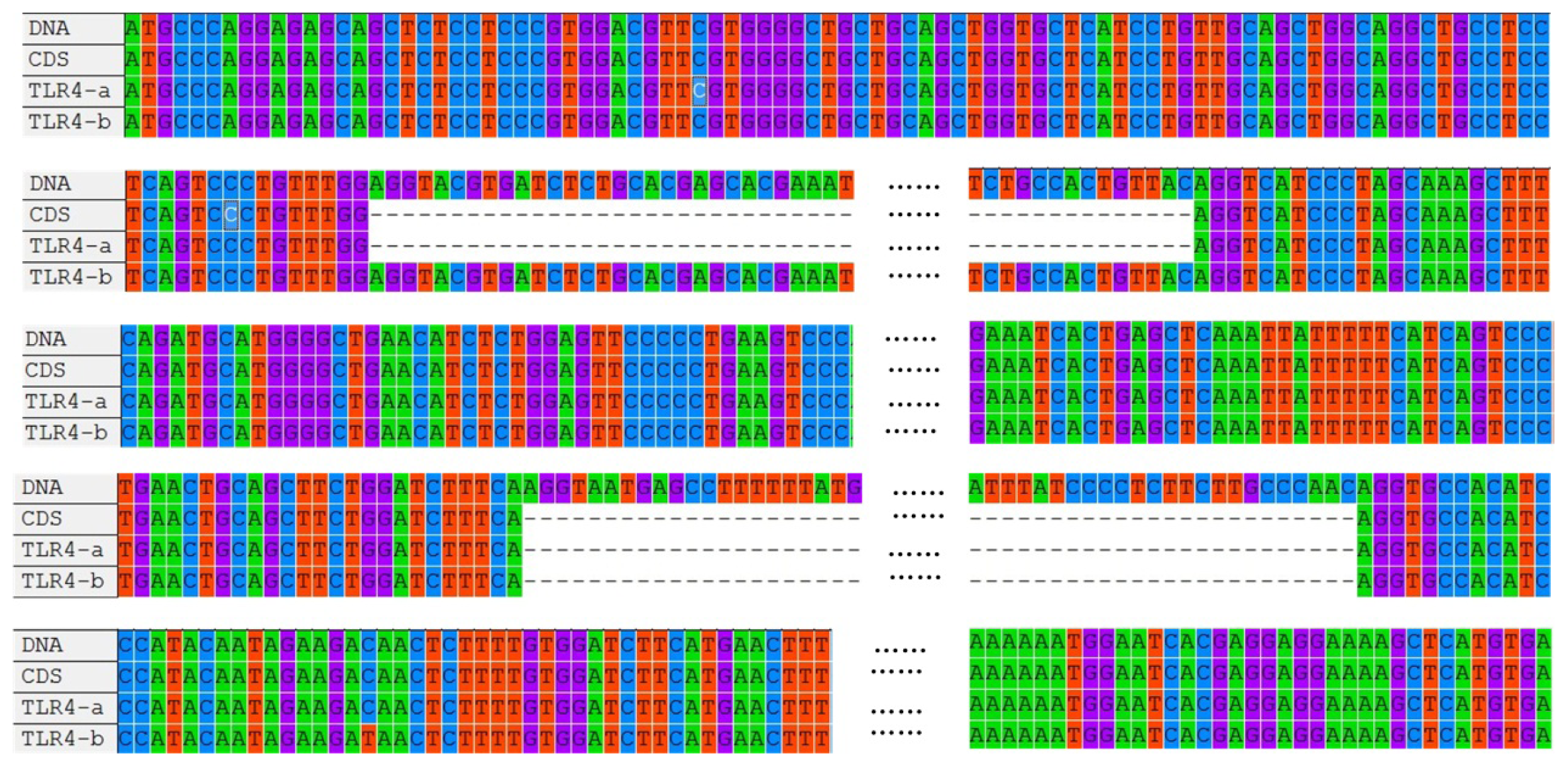
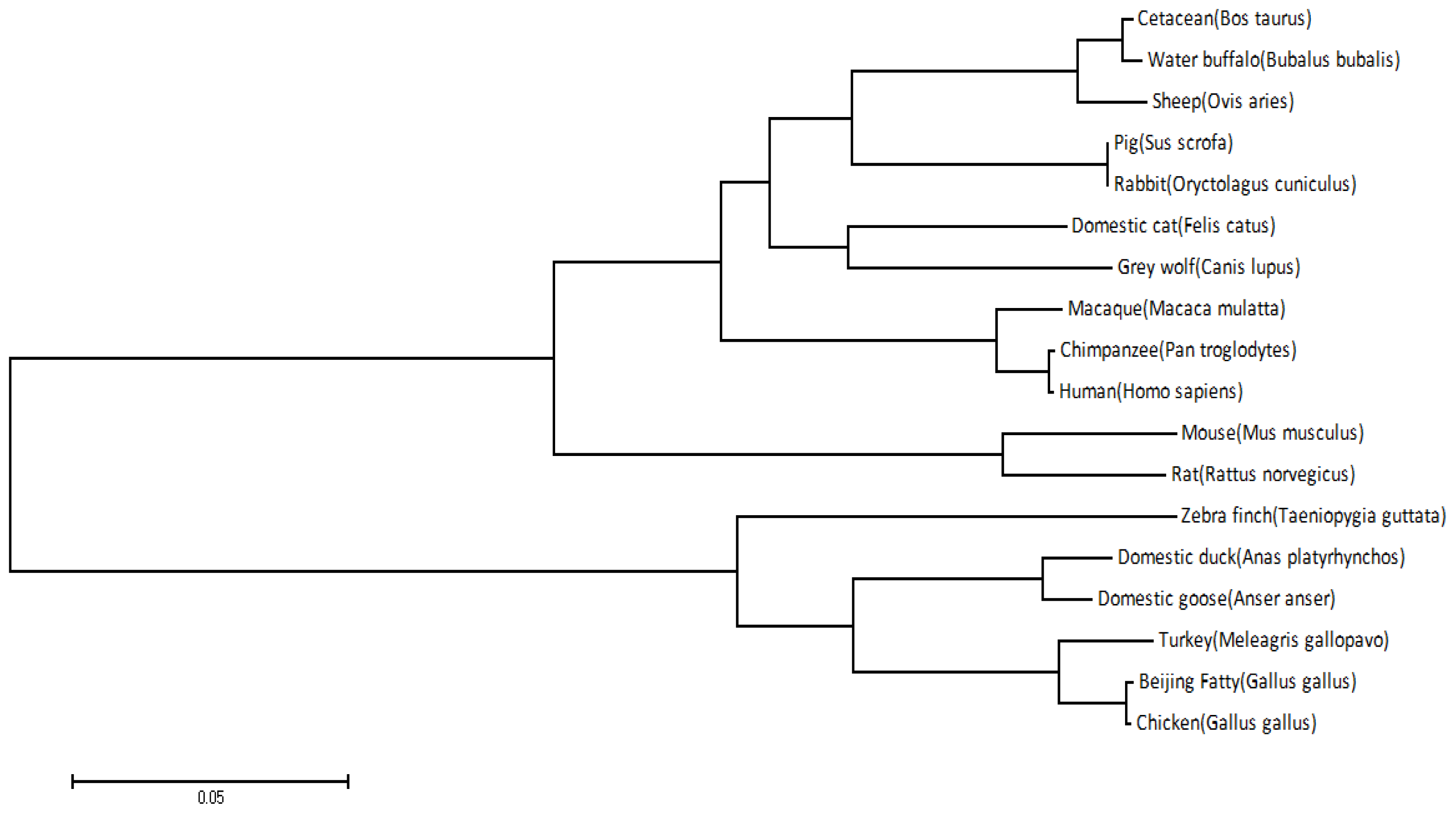
2.1. Cloning and Sequence Analysis of duTLR4
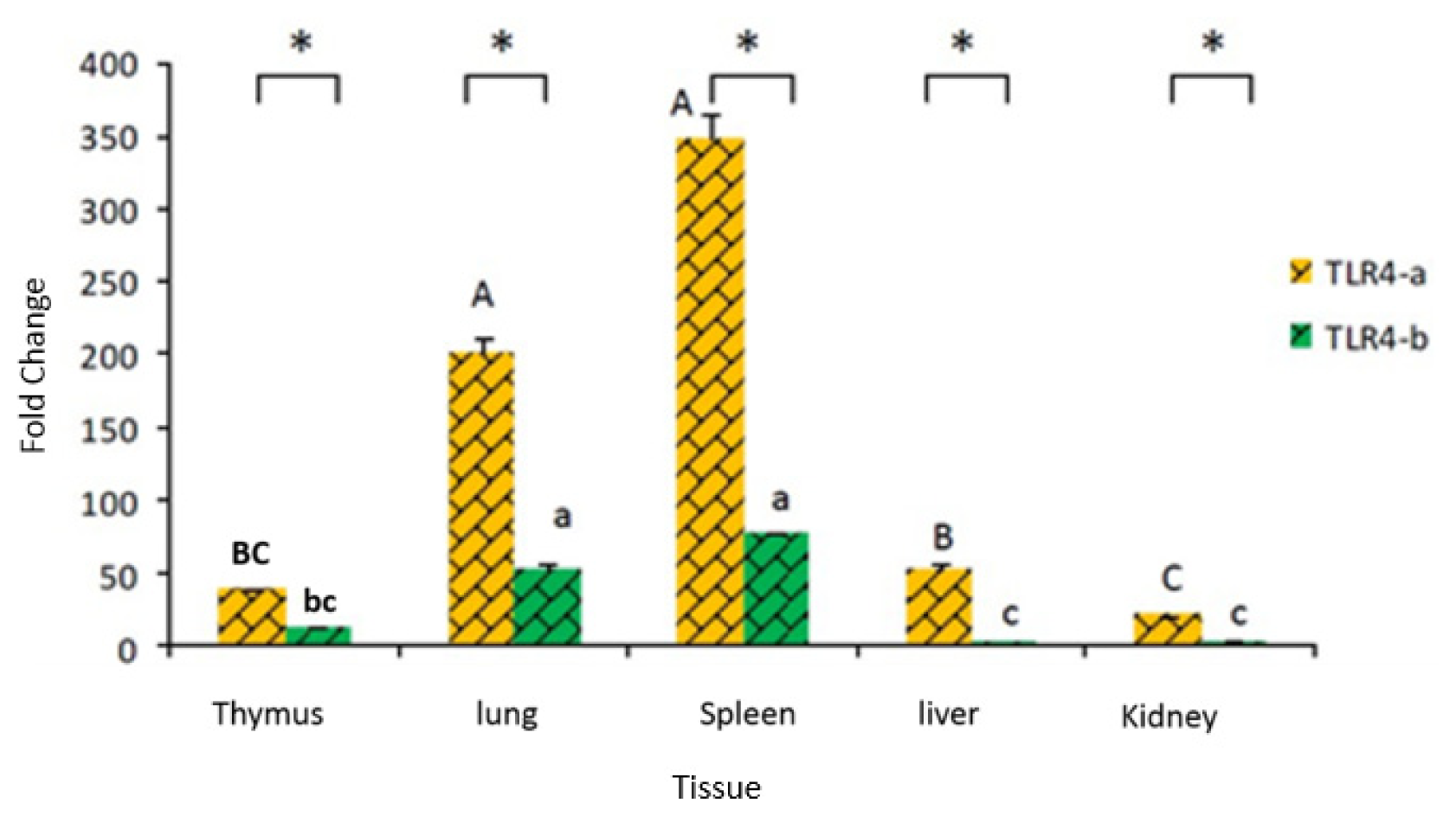
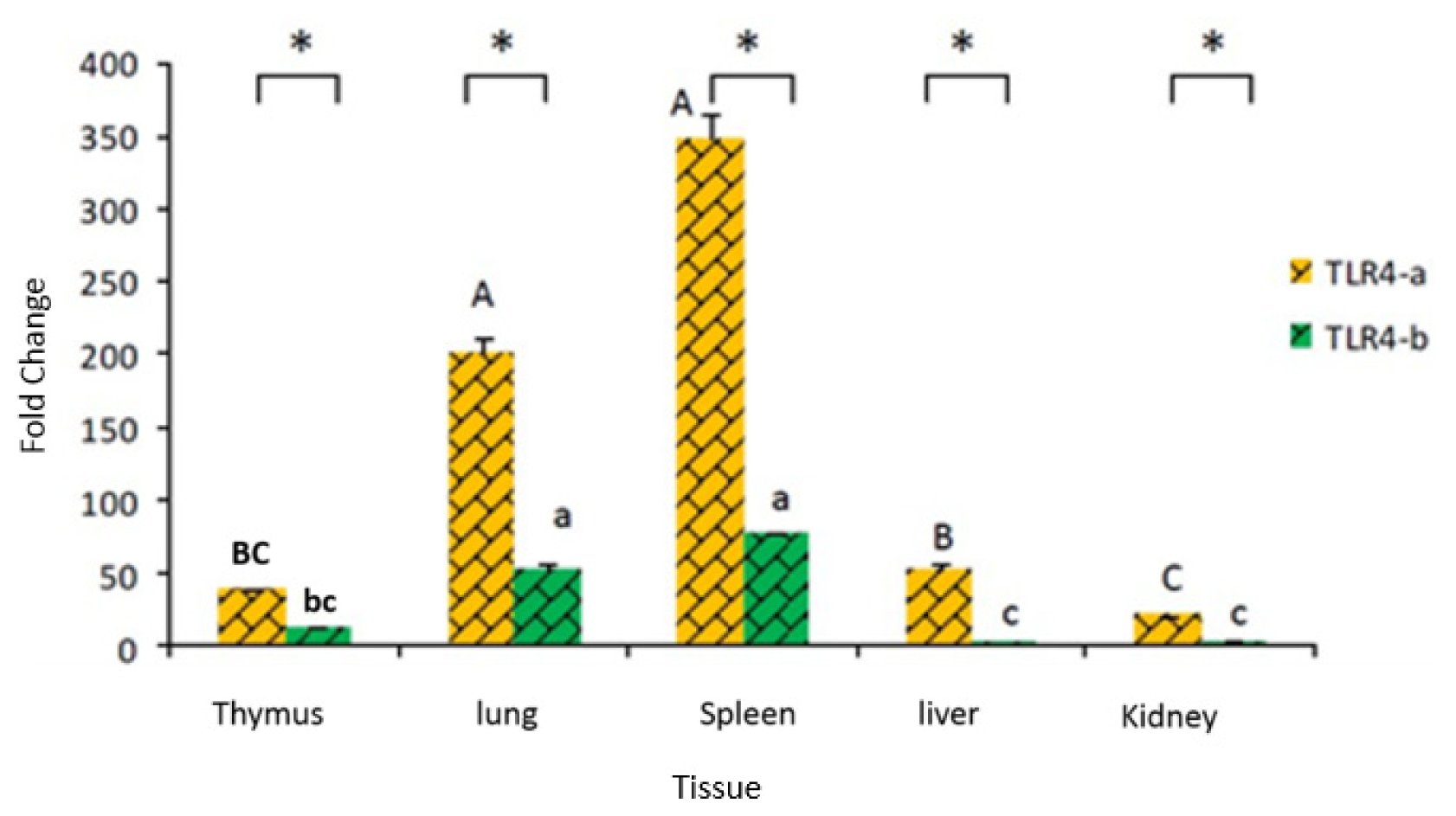
2.2. Tissue Distribution of Duck TLR4 mRNA
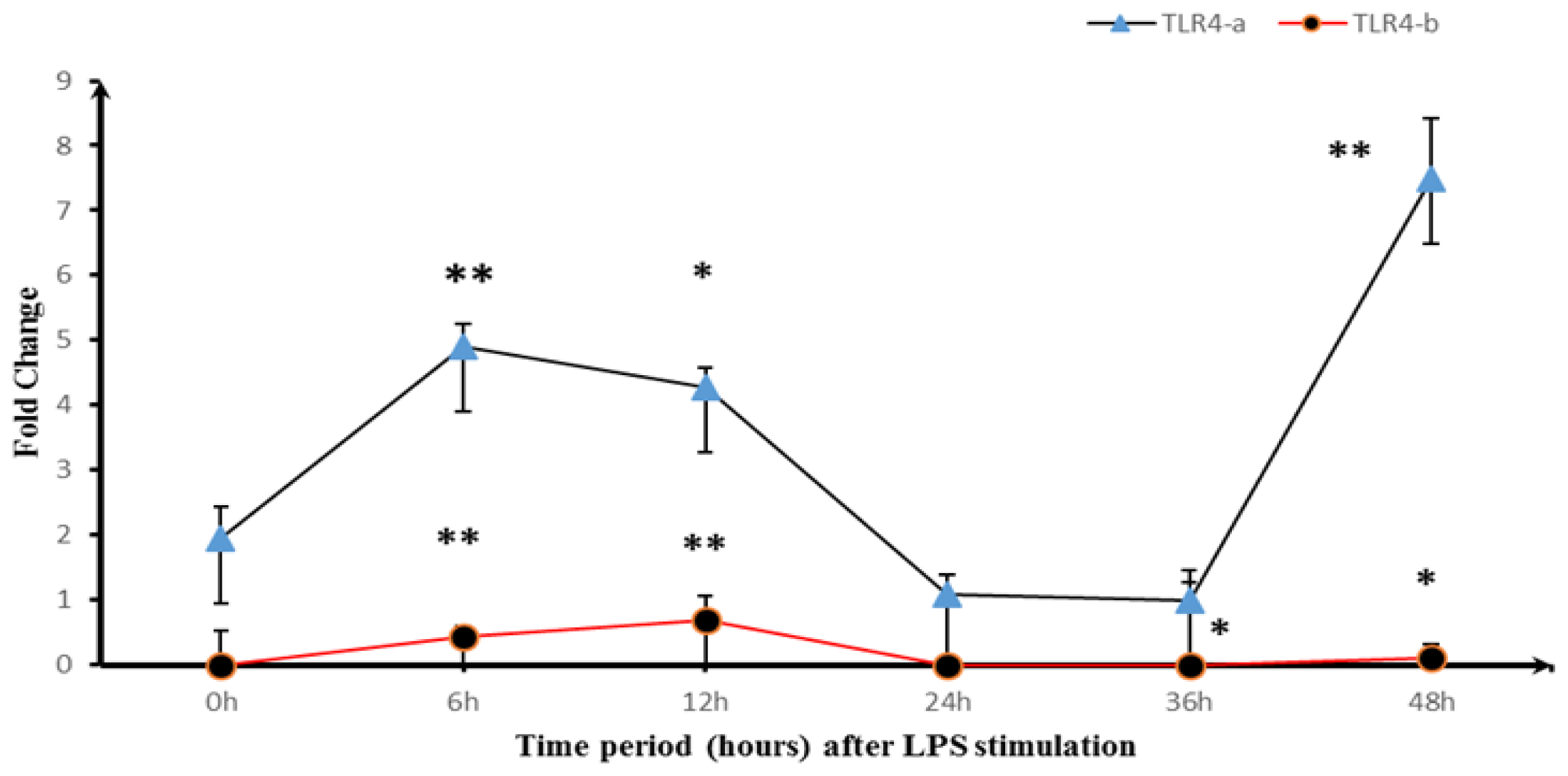
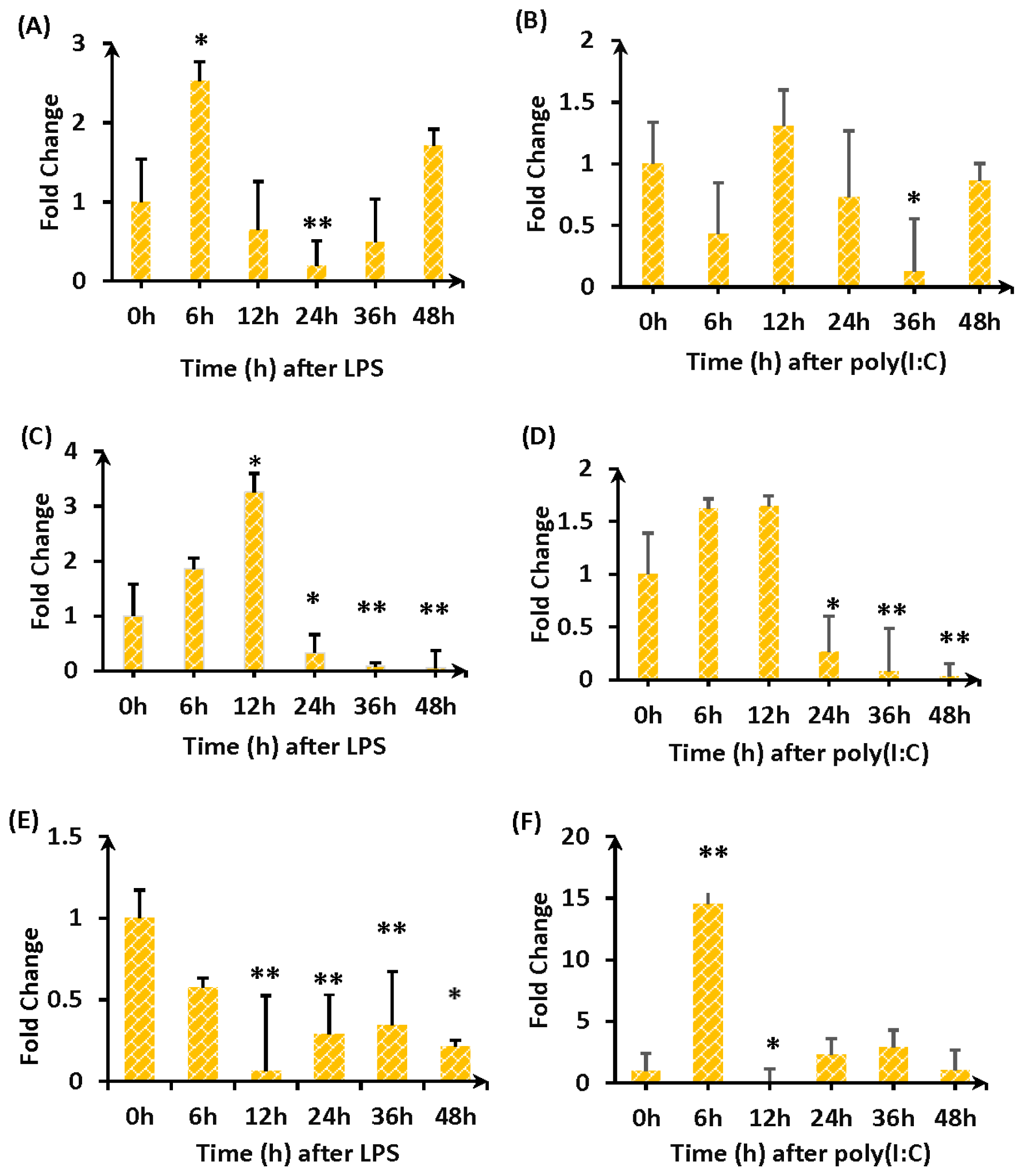
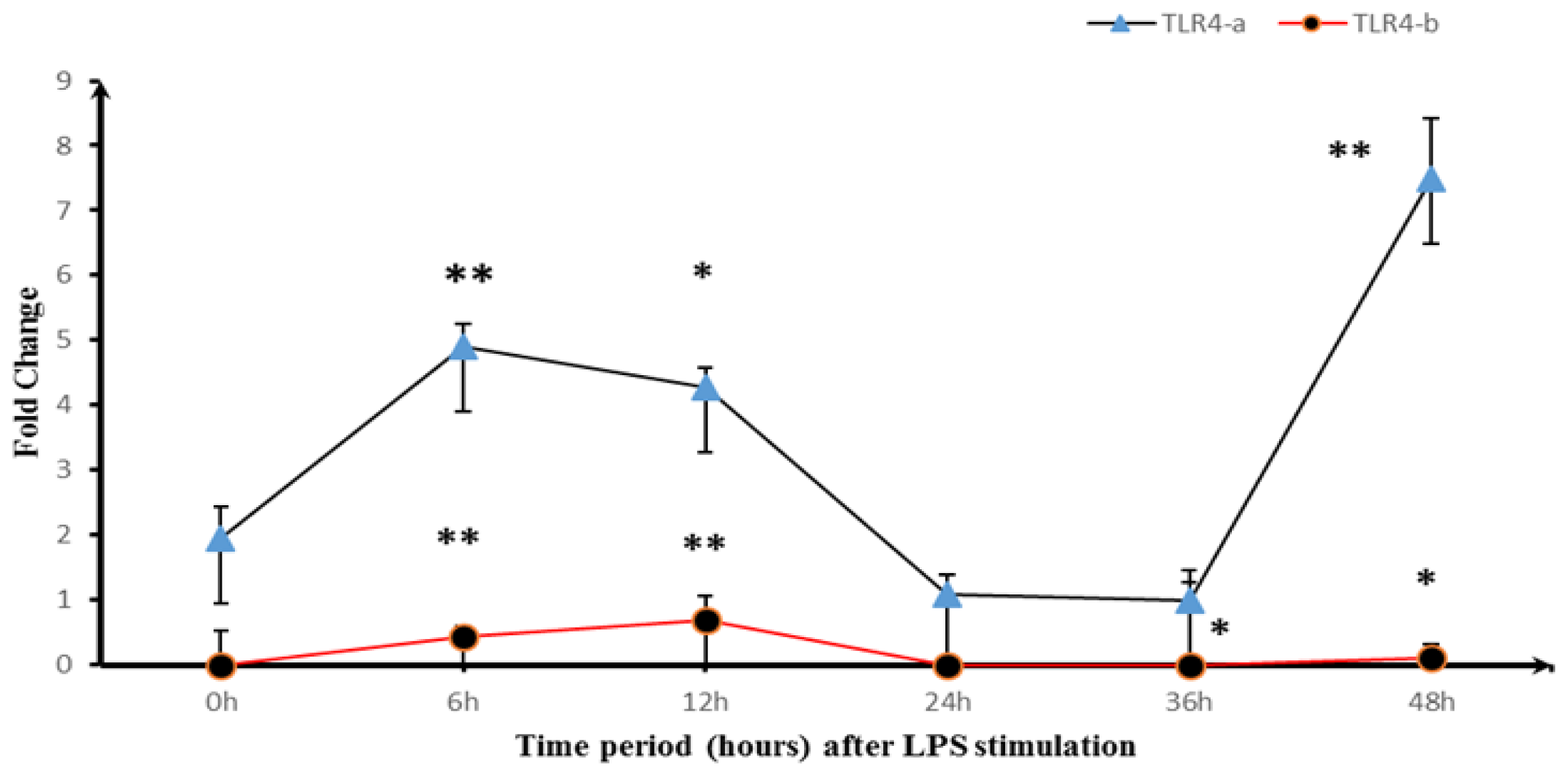
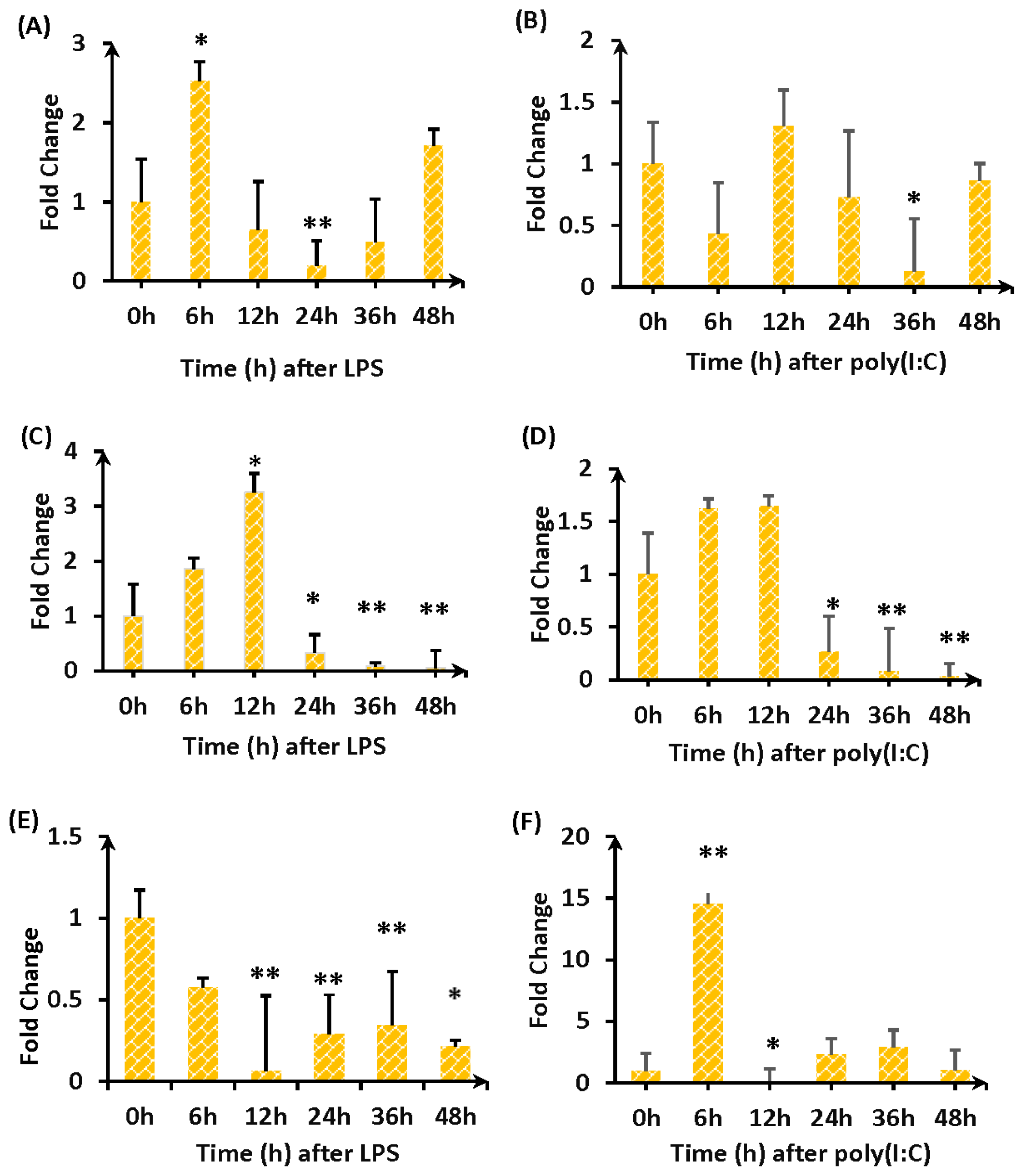
2.3. Expression of TLR4-a and TLR4-b Following Stimulation with LPS
3. Discussion
4. Experimental Section
4.1. Animals and Tissue Sample Collection
4.2. DNA and RNA Isolation and cDNA Synthesis
4.3. Cloning of the TLR4 Gene
4.4. Construction of pcDNA 3.1(+)-TLR4 Overexpression Vector
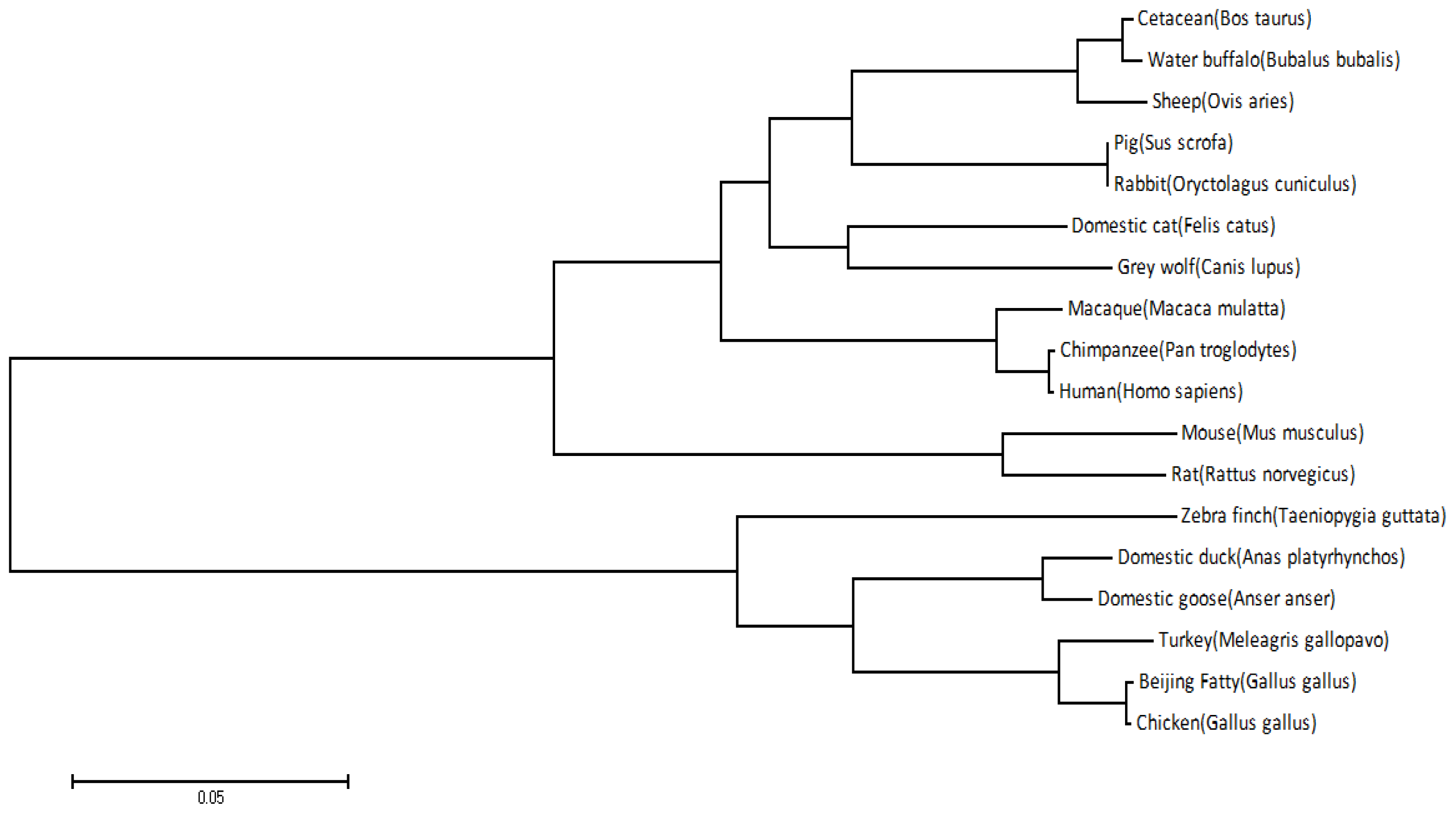
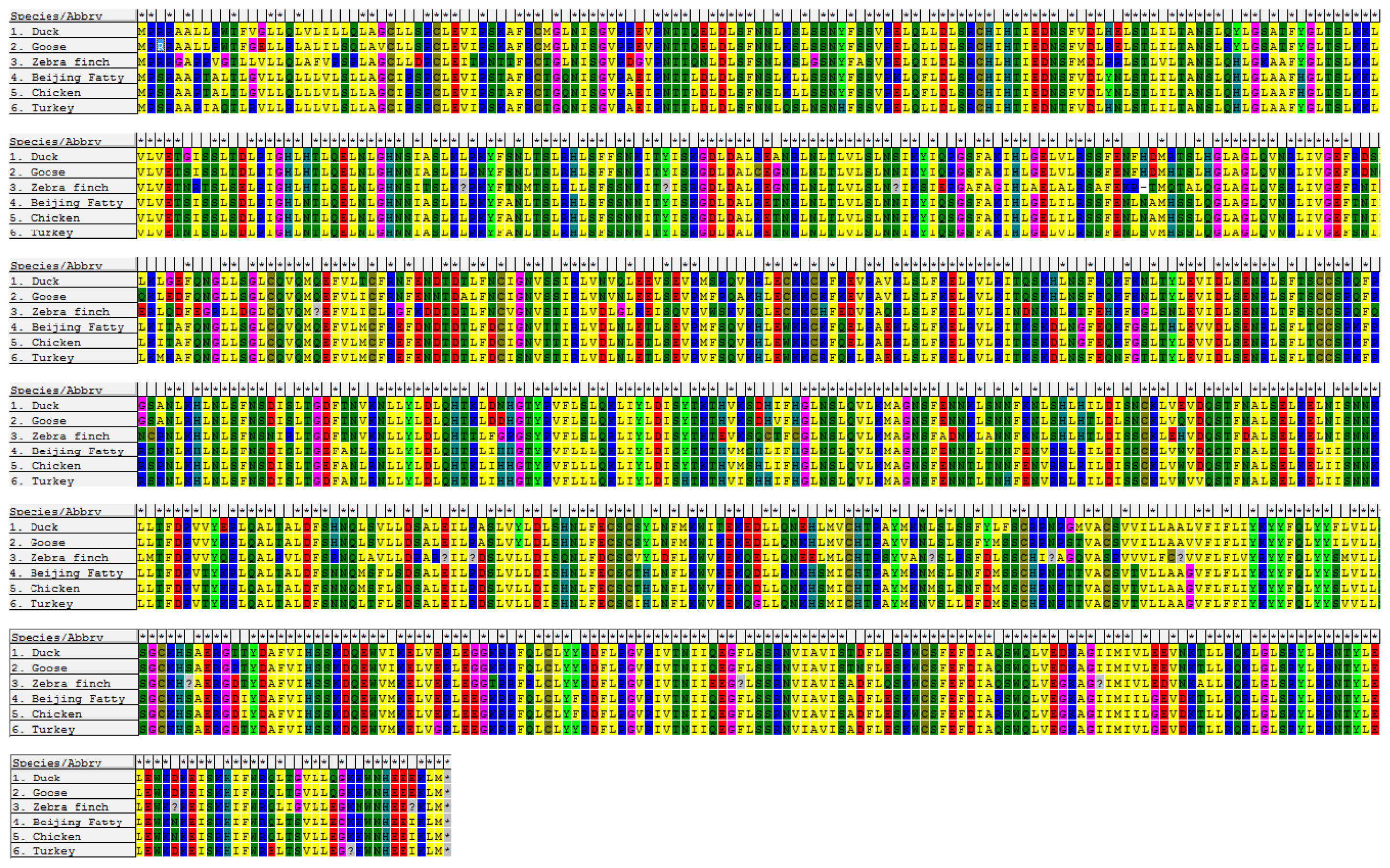
4.5. Sequence Analysis, Multiple Sequence Alignment, and Phylogenetic Analysis
4.6. Real-Time Quantitative PCR
4.7. Expression of TLR4 in Different Tissues
4.8. duTLR4 Expression in Duck Embryo Cells after LPS and Poly (I:C) Challenge
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Akira, S.; Takeda, K.; Kaisho, T. Toll-like receptors: Critical proteins linking innate and acquired immunity. Nat. Immunol 2001, 2, 675–680. [Google Scholar]
- Du, X.; Poltorak, A.; Wei, Y.G.; Beutler, B. Three novel mammalian toll-like receptors: Gene structure, expression, and evolution. Eur. Cytokine Network 2000, 11, 362–371. [Google Scholar]
- Tabeta, K.; Georgel, P.; Janssen, E.; Du, X.; Hoebe, K.; Crozat, K. Toll-like receptors 9 and 3 as essential components of innate immune defense against mouse cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2004, 101, 3516–3521. [Google Scholar]
- Werling, D.; Jungi, T.W. Toll-like receptors linking innate and adaptive immune response. Vet. Immunol. Immunopathol 2003, 91, 1–12. [Google Scholar]
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immunol 2004, 5, 987–995. [Google Scholar]
- Takeda, K.; Akira, S. Toll-like receptors in innate immunity. Int. Immunol 2005, 17, 1–14. [Google Scholar]
- Underhill, D.M.; Ozinsky, A. Toll-like receptors: Key mediators of microbe detection. Curr. Opin. Immunol 2002, 14, 103–110. [Google Scholar]
- He, H.B.; Wu, X.L.; Yu, B.; Liu, K.L.; Zhou, G.X.; Qian, G.Q.; Chen, X.Y. The effect of desacetyluvaricin on the expression of TLR4 and P53 protein in Hepg 2.2.15. Hepat. Mon 2011, 11, 5. [Google Scholar]
- Zhang, B.L.; Du, J.L.; Han, H.B.; Lian, Z.X.; Li, N. Two novel splice variants of the ovis aries Toll-like receptor 4. Asian J. Anim. Vet. Adv 2011, 6, 155–165. [Google Scholar]
- Asahina, Y.; Yoshioka, N.; Kano, R.; Morimoto, T.; Hasegawa, A. Full-length cDNA cloning of Toll-like receptor 4 in dogs and cats. Vet. Immunol. Immunopathol. 2003, 96, 159–167. [Google Scholar]
- Jault, C.; Pichon, L.; Chluba, J. Toll-like receptor gene family and TIR-domain adapters in Danio rerio. Mol. Immunol 2004, 40, 759–771. [Google Scholar]
- Leveque, G.; Forgetta, V.; MorrolL, S.; Smith, A.L.; Bumstead, N.; Barrow, P.; Loredo-OSTI, J.C.; Morgan, K.; Malo, D. Allelic variation in TLR4 is linked to susceptibility to Salmonella enterica serovar Typhimurium infection in chickens. Infect. Immun 2003, 71, 1116–1124. [Google Scholar]
- Teghanemt, A.; Zhang, D.; Levis, E.N.; Weiss, J.P.; Gioannini, T.L. Molecular basis of reduced potency of underacylated endotoxins. J. Immunol 2005, 175, 4669–4676. [Google Scholar]
- Zhang, D.; Zhang, G.; hayden, M.S.; greenblatt, M.B.; Bussey, C.; Flavell, R.A.; Ghosh, S. A toll-like receptor that prevents infection by uropathogenic bacteria. Science 2004, 303, 1522–1526. [Google Scholar]
- Shi, H.; Kokoeva, M.V.; Inouye, K.; Tzameli, I.; Yin, H.; Flier, J.S. TLR4 links innate immunity and fatty acid induced insulin resistance. J. Clin. Invest 2006, 116, 3015–3025. [Google Scholar]
- Yilmaz, A.; Shen, S.X.; Adelson, D.L.; Xavier, S.; Zhu, J.J. Identification and sequence analysis of chicken toll-like receptors. Immunogenetics 2005, 56, 743–753. [Google Scholar]
- Wells, C.A.; Chalk, A.M.; Forrest, A.; Taylor, D.; Waddell, N.; Schroder, K.; Himes, S.R.; Faulkner, G.; Lo, S.; Kasukawa, T.; et al. Alternate transcription of the toll-like receptor signaling cascade. Genome Biol 2006, 7, R10. [Google Scholar]
- Lqbal, M.; Philbin, V.J.; Smith, A.L. Expression patterns of chicken toll-like receptor mRNA in tissues, immune cell subsets and cell lines. Vet. Immunol. Immunopathol 2005, 104, 117–127. [Google Scholar]
- Temperley, N.D.; Berlin, S.; Paton, I.R.; Griffin, D.K.; Burt, D.W. Evolution of the chicken toll-like receptor gene family: A story of gene gain and gene loss. BMC Genomics 2008, 9, 62–73. [Google Scholar]
- Meijer, A.H.; Gabby Krens, S.F.; Medina Rodriguez, I.A.; He, S.; Bitter, W.; Ewa Snaar-Jagalska, B.; Spaink, H.P. Expression analysis of the toll-like receptor and TIR domain adaptor families of zebrafish. Mol. Immunol 2004, 40, 773–783. [Google Scholar]
- Hajjar, A.M.; Ernst, R.K.; Tsai, J.H.; Wilson, C.B.; Miller, S.I. Human Toll-like receptor 4 recognizes host-specific LPS modifications. Nat. Immunol 2002, 3, 354–359. [Google Scholar]
- Muzio, M.; Bosisio, D.; Polentarutti, N.; D’amico, G.; Toppacciaro, A.; Mancinelli, R.; van’t Veer, C.; Penton-Rol, G.; Ruco, L.P.; Allavena, P.; et al. Differential expression and regulation of toll-like receptors (TLR) in human leukocytes: Selective expression of TLR3 in dendritic cells. J. Immunol 2000, 164, 5998–6004. [Google Scholar]
- Park, B.S.; Song, B.H.; Kim, H.M.; Choi, B.; Lee, H.; Lee, J. The structural basis of lipopolysaccharide recognition by the TLR4-MD-2 complex. Nature 2009, 458, 1191–1195. [Google Scholar]
- Visintin, A.; Halmen, K.A.; Latz, E.; Monks, B.G.; Golenbock, D.T. Pharmacological inhibition of endotoxin responses is achieved by targeting the TLR4 coreceptor, MD-2. J. Immunol 2005, 175, 6465–6472. [Google Scholar]
- Chen, H.; Lunney, J.K.; Cheng, L.; Li, X.; Cao, J.; Zhu, M.; Zhao, S. Porcine S100A8 and S100A9: Molecular characterizations and crucial functions in response to Haemophilus parasuis infection. Dev. Comp. Immunol 2011, 35, 490–500. [Google Scholar]
- Saberi, M.; Woods, N.B.; de Luca, C.; Schenk, S.; Lu, J.C.; Bandyopadhyay, G.; Verma, I.M.; Olefsky, J.M. Hematopoietic cell-specific deletion of toll-like receptor 4 ameliorates hepatic and adipose tissue insulin resistance in high-fat-fed mice. Cell Metab 2009, 10, 419–429. [Google Scholar]
- Kogut, M.H.; Iqbal, M.; He, H.; Philbin, V.; Kaiser, P.; Smith, A. Expression and function of toll-like receptors in chicken heterophils. Dev. Comp. Immunol 2005, 29, 791–807. [Google Scholar]
- Remer, K.A.; Brcic, M.; Jungi, T.W. Toll-like receptor-4 is involved in eliciting an LPS-induced oxidative burst in neutrophils. Immunol. Lett 2003, 85, 75–80. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 1997, 25, 3389–3402. [Google Scholar]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol 1999, 112, 531–552. [Google Scholar]
- Pearson, W.R.; Robins, G.; Zhang, T. Generalized neighbor-joining: More reliable phylogenetictree reconstruction. Mol. Biol. Evol 1999, 16, 806–816. [Google Scholar]
- Gale, C.P.; Grant, P.J. The characterisation and functional analysis of the human glyoxalase-1 gene using methods of bioinformatics. Gene 2004, 340, 251–260. [Google Scholar]
- Xia, C.; Lin, C.Y.; Xu, G.X.; Hu, T.J.; Yang, T.Y. cDNA cloning and genomic structure of the duck (Anas platyrhynchos) MHC class I gene. Immunogenetics 2004, 56, 304–309. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer name | Sequence (5′→3′) | Tm (°C) | Note |
|---|---|---|---|
| P1F | CACTTCCCCTTGTTCTCTGC | 65 | coding sequence amplification |
| P1R | AGAGGCAGACAAATGGATGG | ||
| IL-1β-F | CCATGGTGCTTTCTCCTGTGT | 60 | RT-qPCR |
| IL-1β-R | GAGCTTGTAGCCCTTGATGC | ||
| IL-6-F | GAGCTTGTAGCCCTTGATGC | 60 | RT-qPCR |
| IL-6-R | TTCGACGAGGAGAAATGCTT | ||
| MHC II-F | CCACCTTTACCAGCTTCGAG | 60 | RT-qPCR |
| MHC II-R | CCGTTCTTCATCCAGGTGAT | ||
| P3F | CCATGGTGCTTTCTCCTGTGT | 60 | RT-qPCR |
| P3R | CTGGGTGGTGTTTGGGACTT | ||
| P4F | CCCTCTGCTTGGGAGATTTG | 60 | RT-qPCR |
| P4R | GCTTGTTCTGTTTCTCAGGTGTTT | ||
| GAPDH-F | TGCTAAGCGTGTCATCATCT | 60 | RT-qPCR |
| GAPDH-R | AGTGGTCATAAGACCCTCCA |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhao, W.; Huang, Z.; Chen, Y.; Zhang, Y.; Rong, G.; Mu, C.; Xu, Q.; Chen, G. Molecular Cloning and Functional Analysis of the Duck TLR4 Gene. Int. J. Mol. Sci. 2013, 14, 18615-18628. https://doi.org/10.3390/ijms140918615
Zhao W, Huang Z, Chen Y, Zhang Y, Rong G, Mu C, Xu Q, Chen G. Molecular Cloning and Functional Analysis of the Duck TLR4 Gene. International Journal of Molecular Sciences. 2013; 14(9):18615-18628. https://doi.org/10.3390/ijms140918615
Chicago/Turabian StyleZhao, Wenming, Zhengyang Huang, Yang Chen, Yang Zhang, Guanghui Rong, Chunyu Mu, Qi Xu, and Guohong Chen. 2013. "Molecular Cloning and Functional Analysis of the Duck TLR4 Gene" International Journal of Molecular Sciences 14, no. 9: 18615-18628. https://doi.org/10.3390/ijms140918615