Exposure to Enriched Environment Decreases Neurobehavioral Deficits Induced by Neonatal Glutamate Toxicity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
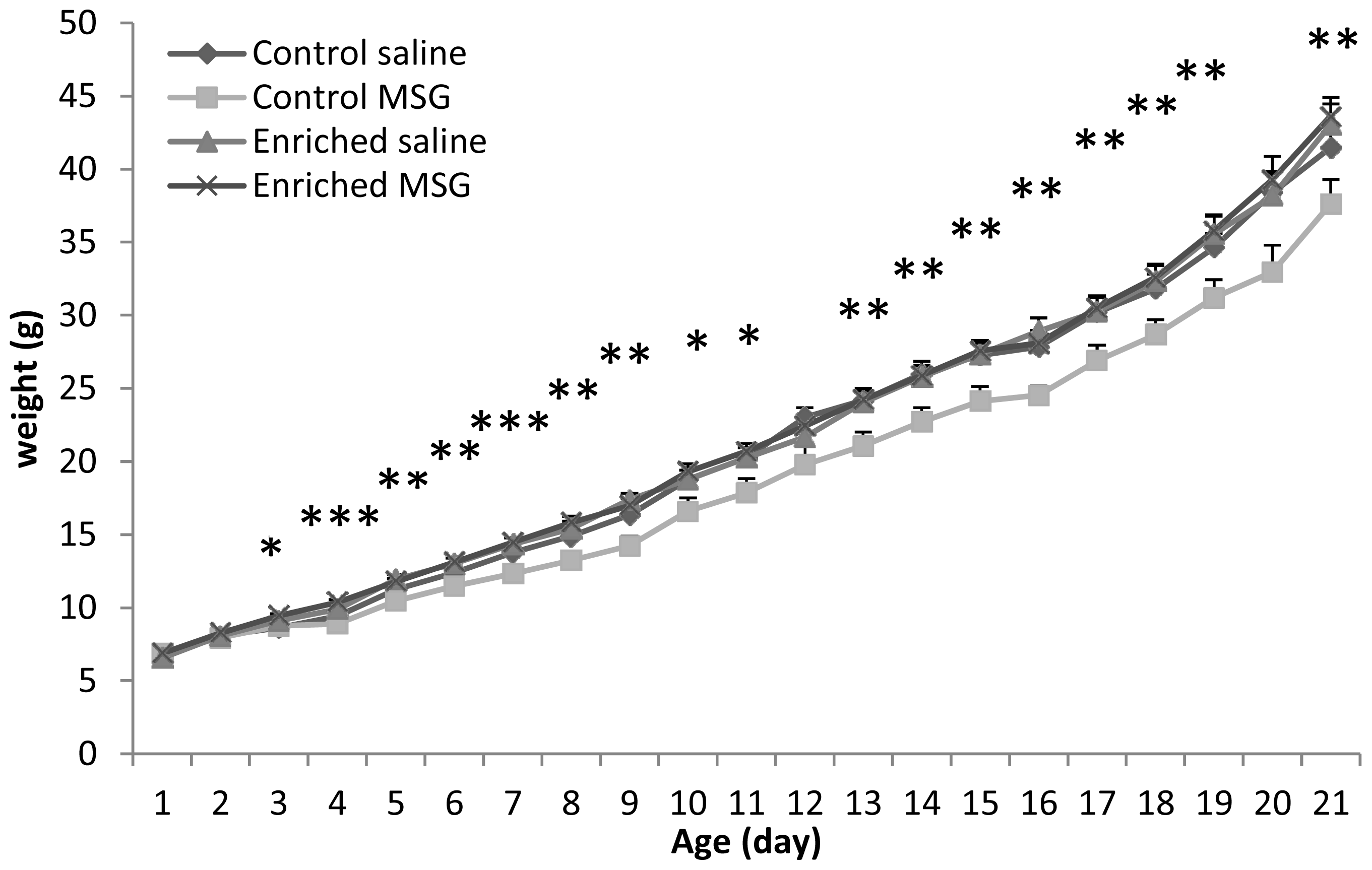
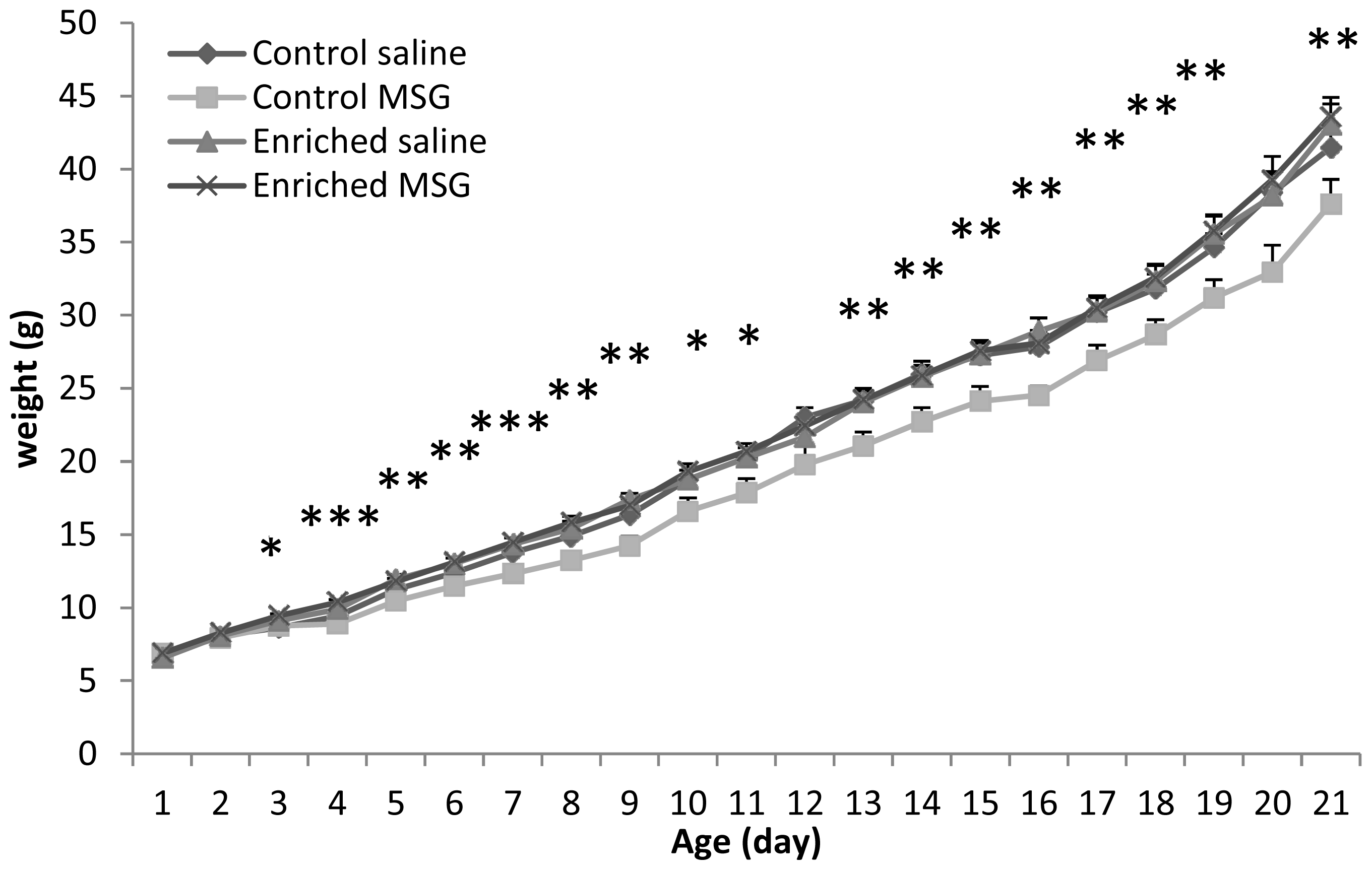
2.1. Somatic Development
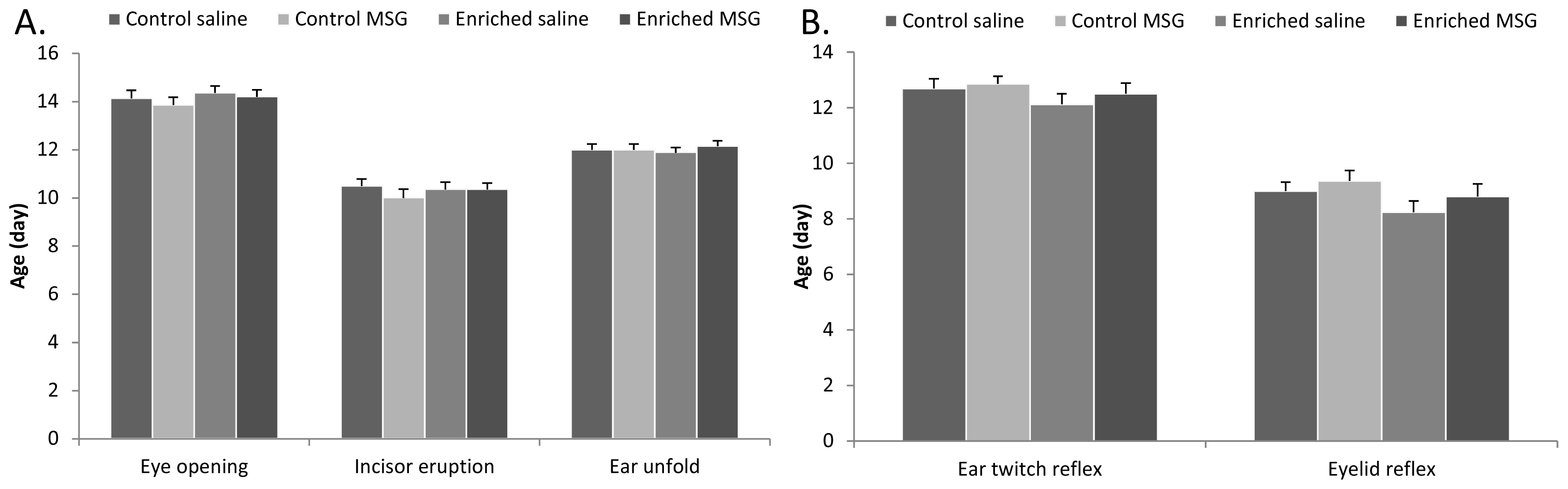
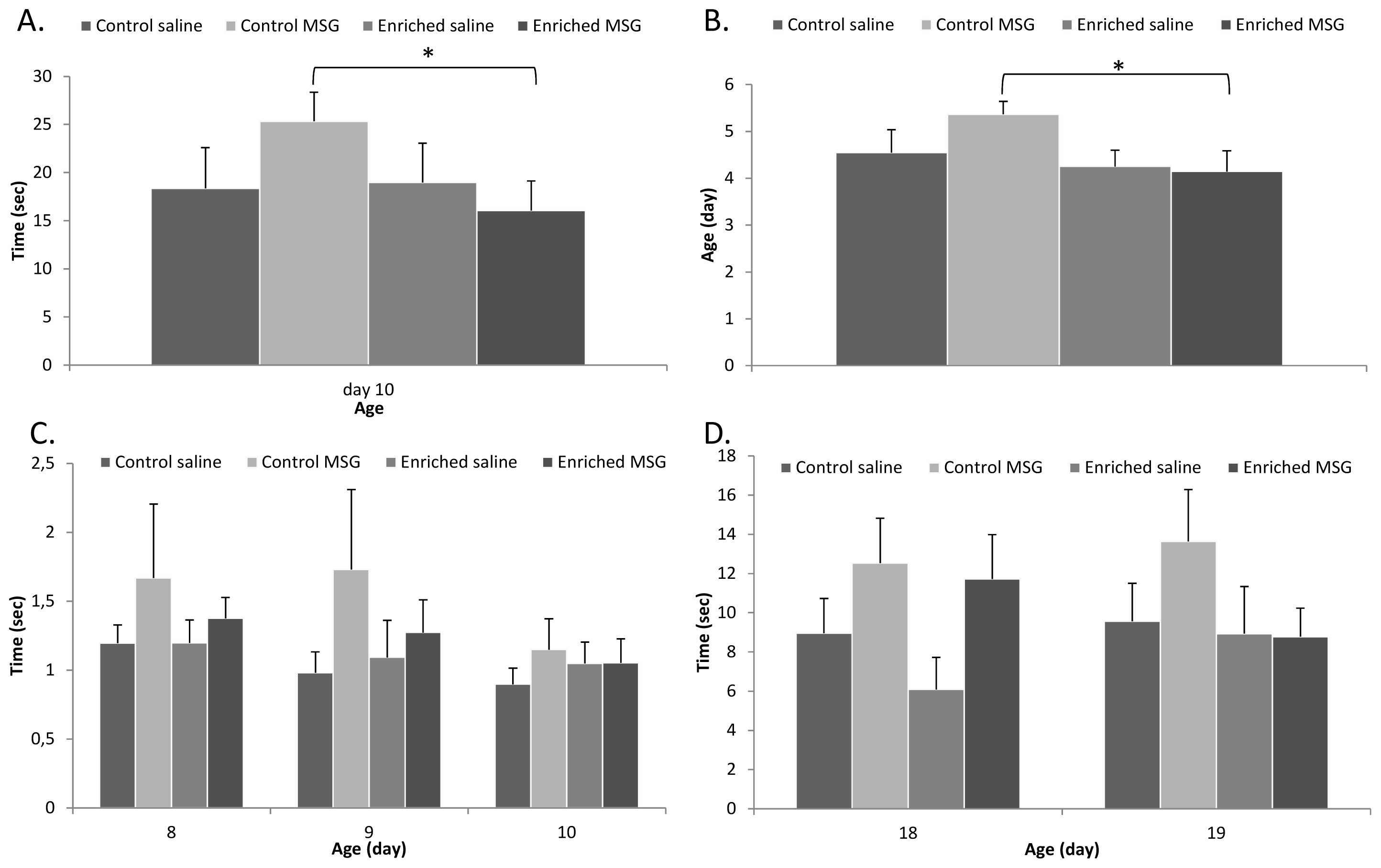
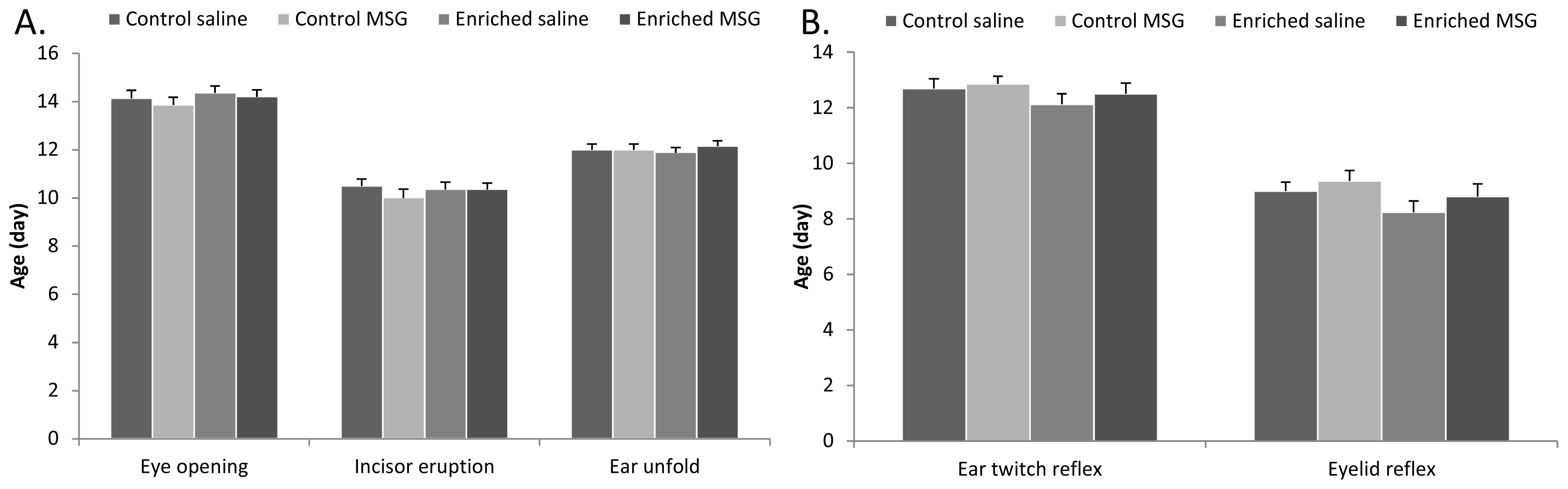
2.2. Physical Parameters and Reflex Development
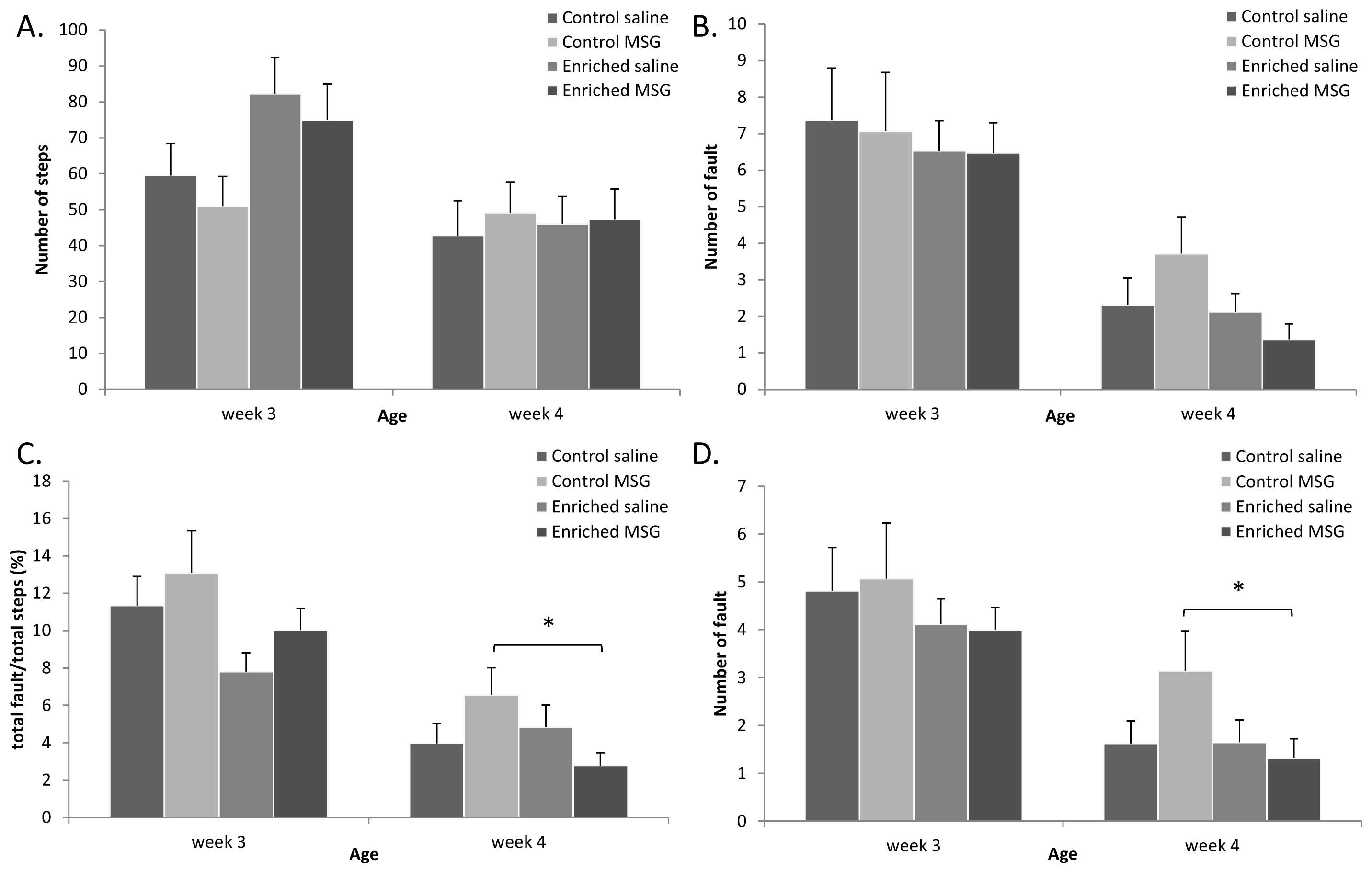
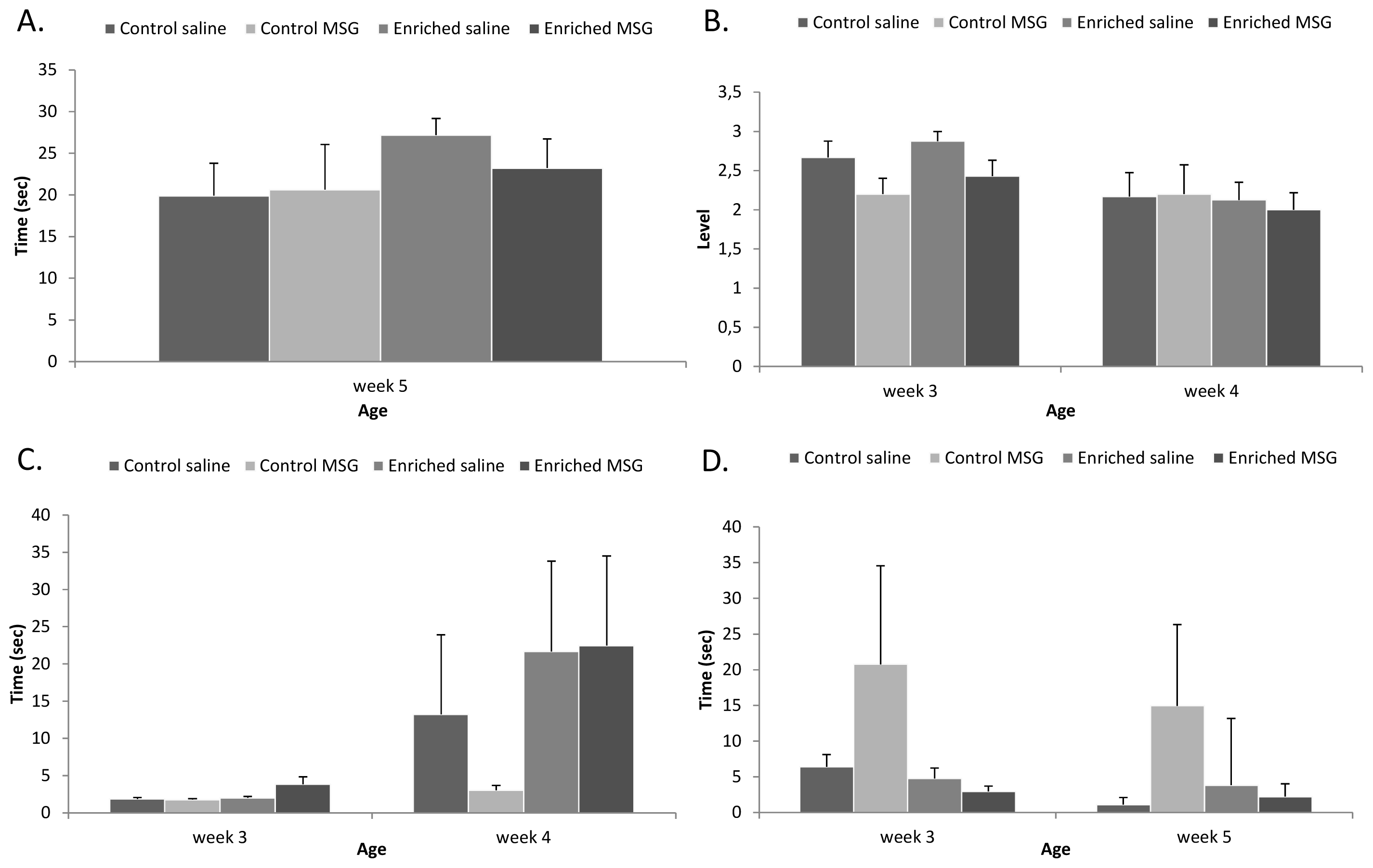
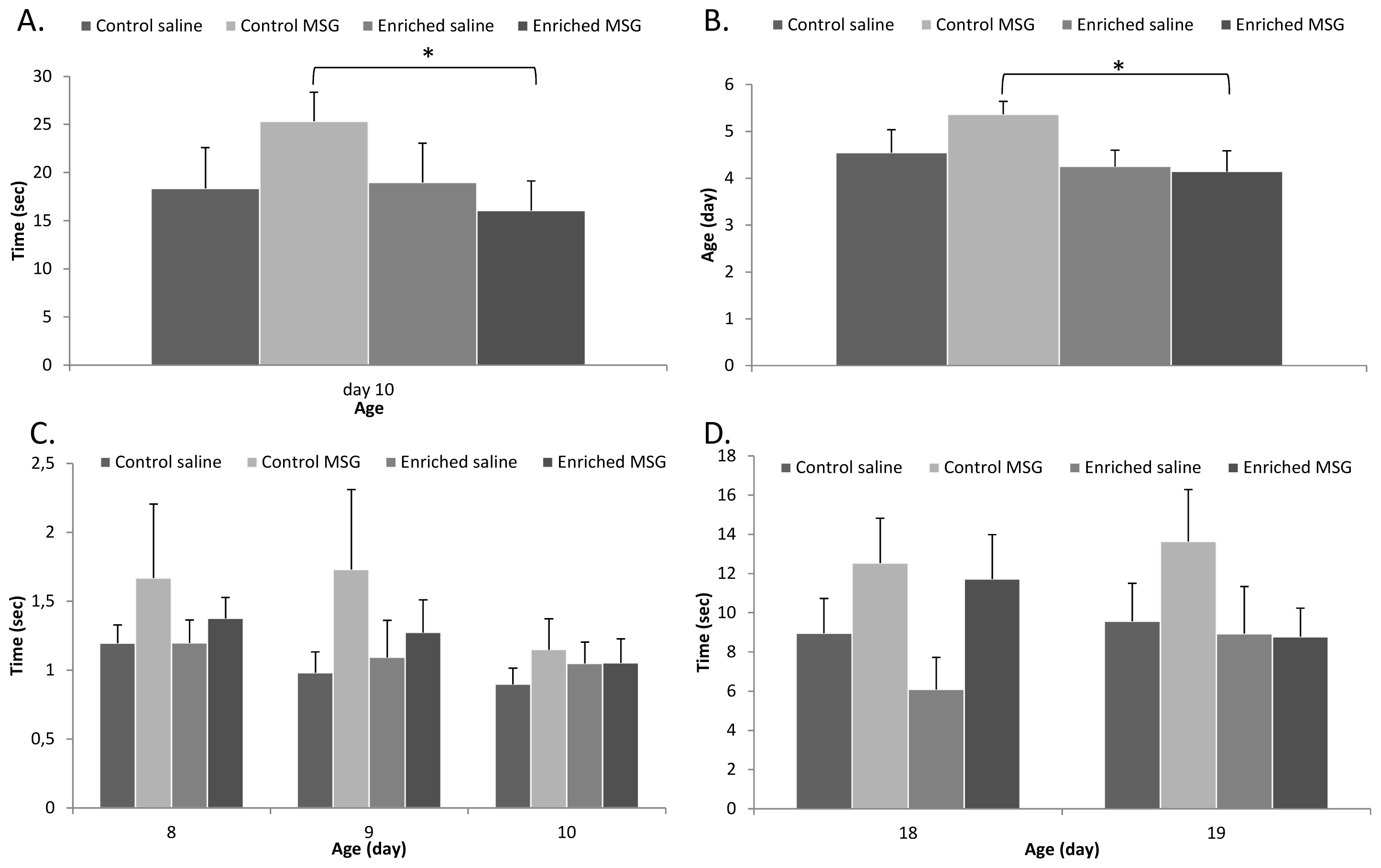
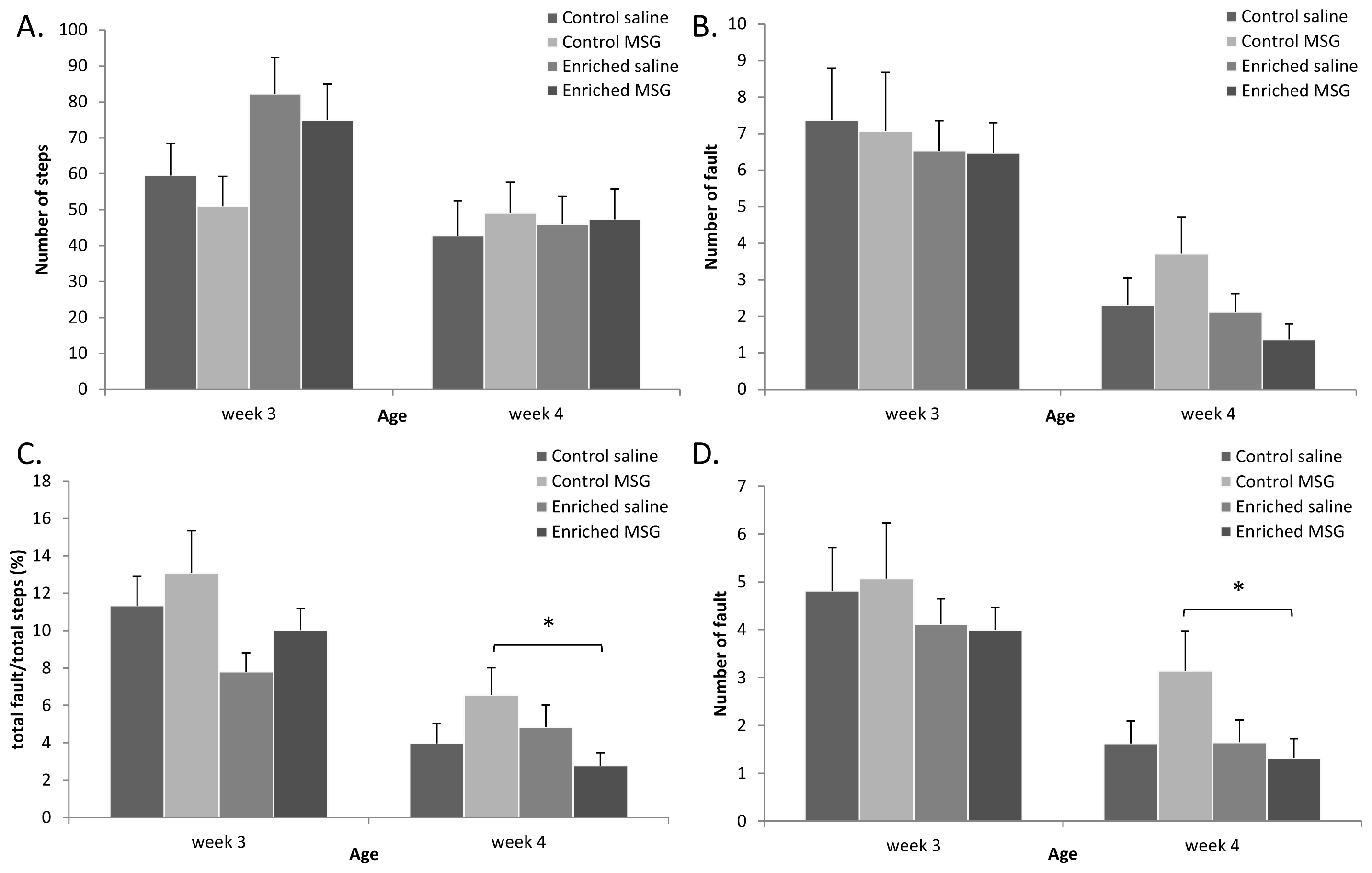
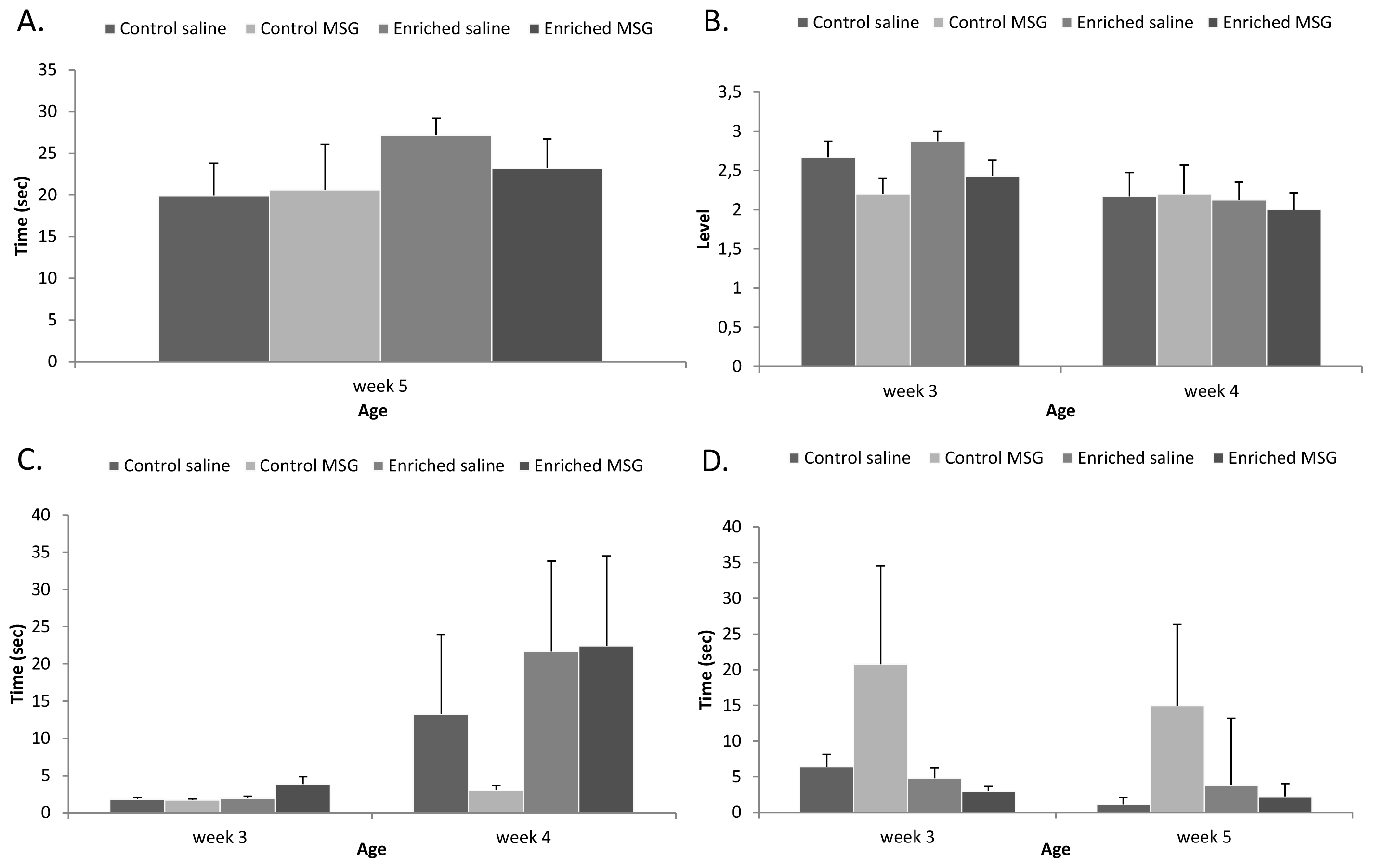
2.3. Motor Coordination
2.4. Discussion
3. Experimental Section
3.1. Experimental Animals
3.2. Environmental Enrichment
3.3. MSG Treatment
3.4. Examination of Neurobehavioral Development
3.5. Motor Coordination Tests
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Hebb, D.O. The effects of early experience on problem solving at maturity. Am. Psychol 1947, 2, 306–307. [Google Scholar]
- Landi, S.; Ciucci, F.; Maffei, L.; Berardi, N.; Cenni, M.C. Setting the pace for retinal development: Environmental enrichment acts through insulin-like growth factor 1 and brain-derived neurotrophic factor. J. Neurosci 2009, 29, 10809–10819. [Google Scholar]
- Sale, A.; Cenni, M.C.; Ciucci, F.; Putignano, E.; Chierzi, S.; Maffei, L. Maternal enrichment during pregnancy accelerates retinal development of the fetus. PLoS One 2007, 2, e1160. [Google Scholar]
- Ortuzar, N.; Argandoña, E.G.; Bengoetxea, H.; Lafuente, J.V. Combination of intracortically administered VEGF and environmental enrichment enhances brain protection in developing rats. J. Neural Transm 2011, 118, 135–144. [Google Scholar]
- Van Praag, H.; Kempermann, G.; Gage, F.H. Neural consequences of environmental enrichment. Nat. Rev. Neurosci 2000, 1, 191–198. [Google Scholar]
- Szabadfi, K.; Atlasz, T.; Horvath, G.; Kiss, P.; Hamza, L.; Farkas, J.; Tamas, A.; Lubics, A.; Gabriel, R.; Reglodi, D. Early postnatal enriched environment decreases retinal degeneration induced by monosodium glutamate treatment. Brain Res 2009, 1259, 107–112. [Google Scholar]
- Kiss, P.; Atlasz, T.; Szabadfi, K.; Horvath, G.; Griecs, M.; Farkas, J.; Matkovits, A.; Toth, G.; Lubics, A.; Tamas, A.; et al. Comparison between PACAP- and enriched environment-induced retinal protection in MSG-treated newborn rats. Neurosci. Lett 2011, 487, 400–405. [Google Scholar]
- Kazl, C.; Foote, L.T.; Kim, M.J.; Koh, S. Early-life experience alters response of developing brain to seizures. Brain Res 2009, 1285, 174–181. [Google Scholar]
- Schneider, T.; Lee, M.H.; Anderson, D.W.; Zuck, L.; Lidsky, T.I. Enriched environment during development is protective against lead-induced neurotoxicity. Brain Res 2001, 896, 48–55. [Google Scholar]
- Soeda, F.; Tanaka, A.; Shirasaki, T.; Takahama, K. An enriched environment mitigates the brain-disruptive effects of prenatal diethylstilbestrol exposure in mice. Neuroscience 2010, 169, 223–228. [Google Scholar]
- Johansson, B.B. Functional outcome in rats transferred to an enriched environment 15 days after focal brain ischemia. Stroke 1996, 27, 324–326. [Google Scholar]
- Ronnback, A.; Dahlqvist, P.; Svensson, P.A.; Jernas, M.; Carlsson, B.; Carlsson, L.M.; Olsson, T. Gene expression profiling of the rat hippocampus one month after focal cerebral ischemia followed by enriched environment. Neurosci. Lett 2005, 385, 173–178. [Google Scholar]
- Zhu, H.; Zhang, J.; Sun, H.; Zhang, L.; Liu, H.; Zeng, X.; Yang, Y.; Yao, Z. An enriched environment reverses the synaptic plasticity deficit induced by chronic cerebral hypoperfusion. Neurosci. Lett 2011, 502, 71–75. [Google Scholar]
- Kovesdi, E.; Gyorgy, A.B.; Kwon, S.K.; Wingo, D.L.; Kamnaksh, A.; Long, J.B.; Kasper, C.E.; Agoston, D.V. The effect of enriched environment on the outcome of traumatic brain injury: A behavioral, proteomics, and histological study. Front. Neurosci 2011, 5. [Google Scholar] [CrossRef]
- Baldini, S.; Restani, L.; Baroncelli, L.; Coltelli, M.; Franco, R.; Cenni, M.C.; Maffei, L.; Berardi, N. Enriched early life experiences reduce adult anxiety-like behavior in rats: A role for insulin-like growth factor 1. J. Neurosci 2013, 33, 11715–11723. [Google Scholar]
- Vazquez-Sanroman, D.; Sanchis-Segura, C.; Toledo, R.; Hernandez, M.E.; Manzo, J.; Miquel, M. The effects of enriched environment on BDNF expression in the mouse cerebellum depending on the length of exposure. Behav. Brain Res 2013, 243, 118–128. [Google Scholar]
- Cao, X.; Huang, S.; Ruan, D. Enriched environment restores impaired hippocampal long-term potentiation and water maze performance induced by developmental lead exposure in rats. Dev. Psychobiol 2008, 50, 307–313. [Google Scholar]
- Beas-Zarate, C.; Perez-Vega, M.I.; Gonzalez-Burgos, I. Neonatal exposure to monosodium l-glutamate induces loss of neurons and cytoarchitectural alterations in hippocampal CA1 pyramidal neurons of adult rats. Brain Res 2002, 952, 275–281. [Google Scholar]
- Chaparro-Huerta, V.; Rivera-Cervantes, M.C.; Torres-Mendoza, B.M.; Beas-Zarate, C. Neuronal death and tumor necrosis factor-α response to glutamate-induced excitotoxicity in the cerebral cortex of neonatal rats. Neurosci. Lett 2002, 333, 95–98. [Google Scholar]
- Kiss, P.; Tamas, A.; Lubics, A.; Szalai, M.; Szalontay, L.; Lengvari, I.; Reglodi, D. Development of neurological reflexes and motor coordination in rats neonatally treated with monosodium glutamate. Neurotox. Res 2005, 8, 235–244. [Google Scholar]
- Kiss, P.; Hauser, D.; Tamas, A.; Lubics, A.; Racz, B.; Horvath, Z.; Farkas, J.; Zimmermann, F.; Stepien, A.; Lengvari, I.; et al. Changes in open-field activity and novelty-seeking behavior in periadolescent rats neonatally treated with monosodium glutamate. Neurotox. Res 2007, 12, 85–93. [Google Scholar]
- Tamas, A.; Gabriel, R.; Racz, B.; Denes, V.; Kiss, P.; Lubics, A.; Lengvari, I.; Reglodi, D. Effects of pituitary adenylate cyclase activating polypeptide in retinal degeneration induced by monosodium-glutamate. Neurosci. Lett 2004, 372, 110–113. [Google Scholar]
- Farkas, J.; Reglodi, D.; Gaszner, B.; Szogyi, D.; Horvath, G.; Lubics, A.; Tamas, A.; Frank, F.; Besirevic, D.; Kiss, P. Effects of maternal separation on the neurobehavioral development of newborn Wistar rats. Brain Res. Bull 2009, 79, 208–214. [Google Scholar]
- Kiss, P.; Szogyi, D.; Reglodi, D.; Horvath, G.; Farkas, J.; Lubics, A.; Tamas, A.; Atlasz, T.; Szabadfi, K.; Babai, N.; et al. Effects of perinatal asphyxia on the neurobehavioral and retinal development of newborn rats. Brain. Res 2009, 1255, 42–50. [Google Scholar]
- Lubics, A.; Reglodi, D.; Tamas, A.; Kiss, P.; Szalai, M.; Szalontay, L.; Lengvari, I. Neurological reflexes and early motor behavior in rats subjected to neonatal hypoxic/ischemic injury. Behav. Brain Res 2005, 157, 157–165. [Google Scholar]
- Mesa-Gresa, P.; Pérez-Martinez, A.; Redolat, R. Behavioral effects of combined environmental enrichment and chronic nicotine administration in male NMRI mice. Physiol. Behav. 2013, 114–115, 65–76. [Google Scholar]
- Bodnar, I.; Göõz, P.; Okamura, H.; Toth, B.E.; Vecsernye, M.; Halasz, B.; Nagy, G.M. Effect of neonatal treatment with monosodium glutamate on dopaminergic and L-DOPA-ergic neurons of the medial basal hypothalamus and on prolactin and MSH secretion of rats. Brain Res. Bull 2001, 55, 767–774. [Google Scholar]
- Dawson, R., Jr. Developmental and sex-specific effects of low dose neonatal monosodium glutamate administration on mediobasal hypothalamic chemistry. Neuroendocrinology 1986, 42, 158–166. [Google Scholar]
- Dubovicky, M.; Tokarev, D.; Skultetyova, I.; Jezova, D. Changes of exploratory behaviour and its habituation in rats neonatally treated with monosodium glutamate. Pharmacol. Biochem. Behav 1997, 56, 565–569. [Google Scholar]
- Miquel, M.; Font, L.; Sanchis-Segura, C.; Aragon, C.M. Neonatal administration of monosodium glutamate prevents the development of ethanol- but not psychostimulant-induced sensitization: A putative role of the arcuate nucleus. Eur. J. Neurosci 2003, 17, 2163–2170. [Google Scholar]
- Hlinak, Z.; Gandalovicova, D.; Krejci, I. Behavioral deficits in adult rats treated neonatally with glutamate. Neurotoxicol. Teratol 2005, 27, 465–473. [Google Scholar]
- Szabadfi, K.; Atlasz, T.; Reglodi, D.; Kiss, P.; Danyadi, B.; Fekete, E.M.; Zorrilla, E.P.; Tamas, A.; Szabo, K.; Gabriel, R. Urocortin 2 protects against retinal degeneration following bilateral common carotid artery occlusion in the rat. Neurosci. Lett 2009, 455, 42–45. [Google Scholar]
- Atlasz, T.; Babai, N.; Reglodi, D.; Kiss, P.; Tamas, A.; Bari, F.; Domoki, F.; Gabriel, R. Diazoxide is protective in the rat retina against ischemic injury induced by bilateral carotid occlusion and glutamate-induced degeneration. Neurotox. Res 2007, 12, 105–111. [Google Scholar]
- Reglodi, D.; Kiss, P.; Tamas, A.; Lengvari, I. The effects of PACAP and PACAP antagonist on the neurobehavioral development of newborn rats. Behav. Brain Res 2003, 140, 131–139. [Google Scholar]
- Reynolds, S.; Urruela, M.; Devine, D.P. Effects of environmental enrichment on repetitive behaviors in the BTBR T + tf/J mouse model of autism. Autism Res. 2013. [Google Scholar] [CrossRef]
- De Carvalho Mendes, F.; de Almeida, M.N.; Felício, A.P.; Fadel, A.C.; de Jesus Silva, D.; Borralho, T.G.; da Silva, R.P.; Bento-Torres, J.; Vasconcelos, P.F.; Perry, V.H.; et al. Enriched environment and masticatory activity rehabilitation recover spatial memory decline in aged mice. BMC Neurosci. 2013, 14. [Google Scholar] [CrossRef]
- Richter, S.H.; Zeuch, B.; Riva, M.A.; Gass, P.; Vollmayr, B. Environmental enrichment ameliorates depressive-like symptoms in young rats bred for learned helplessness. Behav. Brain Res 2013, 252, 287–292. [Google Scholar]
- Ravenelle, R.; Byrnes, E.M.; Byrnes, J.J.; McInnis, C.; Park, J.H.; Donaldson, S.T. Environmental enrichment effects on the neurobehavioral profile of selective outbred trait anxiety rats. Behav. Brain Res 2013, 252, 49–57. [Google Scholar]
- Pattison, K.F.; Laude, J.R.; Zentall, T.R. Environmental enrichment affects suboptimal, risky, gambling-like choice by pigeons. Anim. Cogn 2013, 16, 429–434. [Google Scholar]
- Johnson, E.M.; Traver, K.L.; Hoffman, S.W.; Harrison, C.R.; Herman, J.P. Environmental enrichment protects against functional deficits caused by traumatic brain injury. Front. Behav. Neurosci. 2013, 7. [Google Scholar] [CrossRef]
- Urakawa, S.; Hida, H.; Masuda, T.; Misumi, S.; Kim, T.S.; Nishino, H. Environmental enrichment brings a beneficial effect on beam walking and enhances the migration of doublecortin-positive cells following striatal lesions in rats. Neuroscience 2007, 144, 920–933. [Google Scholar]
- Monaco, C.M.; Mattiola, V.V.; Folweiler, K.A.; Tay, J.K.; Yelleswarapu, N.K.; Curatolo, L.M.; Matter, A.M.; Cheng, J.P.; Kline, A.E. Environmental enrichment promotes robust functional and histological benefits in female rats after controlled cortical impact injury. Exp. Neurol 2013, 247, 410–418. [Google Scholar]
- Rojas, J.J.; Deniz, B.F.; Miguel, P.M.; Diaz, R.; do Hermel, E.E.; Achaval, M.; Nettó, C.A.; Pereira, L.O. Effects of daily environmental enrichment on behavior and dendritic spine density in hippocampus following neonatal hypoxia-ischemia in the rat. Exp. Neurol 2013, 241, 25–33. [Google Scholar]
- Baroncelli, L.; Braschi, C.; Maffei, L. Visual depth perception in normal and deprived rats: Effects of environmental enrichment. Neuroscience 2013, 236, 313–319. [Google Scholar]
- Thiriet, N.; Gennequin, B.; Lardeux, V.; Chauvet, C.; Decressac, M.; Janet, T.; Jaber, M.; Solinas, M. Environmental enrichment does not reduce the rewarding and neurotoxic effects of methamphetamine. Neurotox. Res 2011, 19, 172–182. [Google Scholar]
- Lestaevel, P.; Airault, F.; Racine, R.; Bensoussan, H.; Dhieux, B.; Delissen, O.; Manens, L.; Aigueperse, J.; Voisin, P.; Souidi, M. Influence of environmental enrichment and depleted uranium on behaviour, cholesterol and acetylcholine in apolipoprotein E-deficient mice. J. Mol. Neurosci. 2013, in press. [Google Scholar]
- Fisher, K.N.; Turner, R.A.; Pineault, G.; Kleim, J.; Saari, M.J. The postweaning housing environment determines expression of learning deficit associated with neonatal monosodium glutamate (MSG). Neurotoxicol. Teratol 1991, 13, 507–513. [Google Scholar]
- Vivinetto, A.L.; Suarez, M.M.; Rivarola, M.A. Neurobiological effects of neonatal maternal separation and post-weaning environmental enrichment. Behav. Brain Res 2013, 240, 110–118. [Google Scholar]
- Babai, N.; Atlasz, T.; Tamas, A.; Reglodi, D.; Toth, G.; Kiss, P.; Gabriel, R. Degree of damage compensation by various PACAP treatments in monosodium glutamate-induced retinal degeneration. Neurotox. Res 2005, 8, 227–233. [Google Scholar]
- Altman, J.; Sudarshan, K. Postnatal development of locomotion in the laboratory rat. Anim. Behav 1975, 23, 896–920. [Google Scholar]
- Hill, J.M.; Gozes, I.; Hill, J.L.; Fridkin, M.; Brenneman, D.E. Vasoactive intestinal peptide antagonist retards the development of neonatal behaviors in the rat. Peptides 1991, 12, 187–192. [Google Scholar]
- Smart, J.L.; Dobbing, J. Vulnerability of developing brain. II. Effects of early nutritional deprivation on reflex ontogeny and development on behavior in the rat. Brain Res 1971, 28, 85–95. [Google Scholar]
- Smart, J.L.; Dobbing, J. Vulnerability of developing brain. VI. Relative effects of foetal and early postnatal undernutrition on reflex ontogeny and development of behavior in the rat. Brain Res 1971, 33, 303–314. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Horvath, G.; Reglodi, D.; Vadasz, G.; Farkas, J.; Kiss, P. Exposure to Enriched Environment Decreases Neurobehavioral Deficits Induced by Neonatal Glutamate Toxicity. Int. J. Mol. Sci. 2013, 14, 19054-19066. https://doi.org/10.3390/ijms140919054
Horvath G, Reglodi D, Vadasz G, Farkas J, Kiss P. Exposure to Enriched Environment Decreases Neurobehavioral Deficits Induced by Neonatal Glutamate Toxicity. International Journal of Molecular Sciences. 2013; 14(9):19054-19066. https://doi.org/10.3390/ijms140919054
Chicago/Turabian StyleHorvath, Gabor, Dora Reglodi, Gyongyver Vadasz, Jozsef Farkas, and Peter Kiss. 2013. "Exposure to Enriched Environment Decreases Neurobehavioral Deficits Induced by Neonatal Glutamate Toxicity" International Journal of Molecular Sciences 14, no. 9: 19054-19066. https://doi.org/10.3390/ijms140919054