Receptor-Targeted, Magneto-Mechanical Stimulation of Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells

Abstract
:1. Introduction
2. Results and Discussion
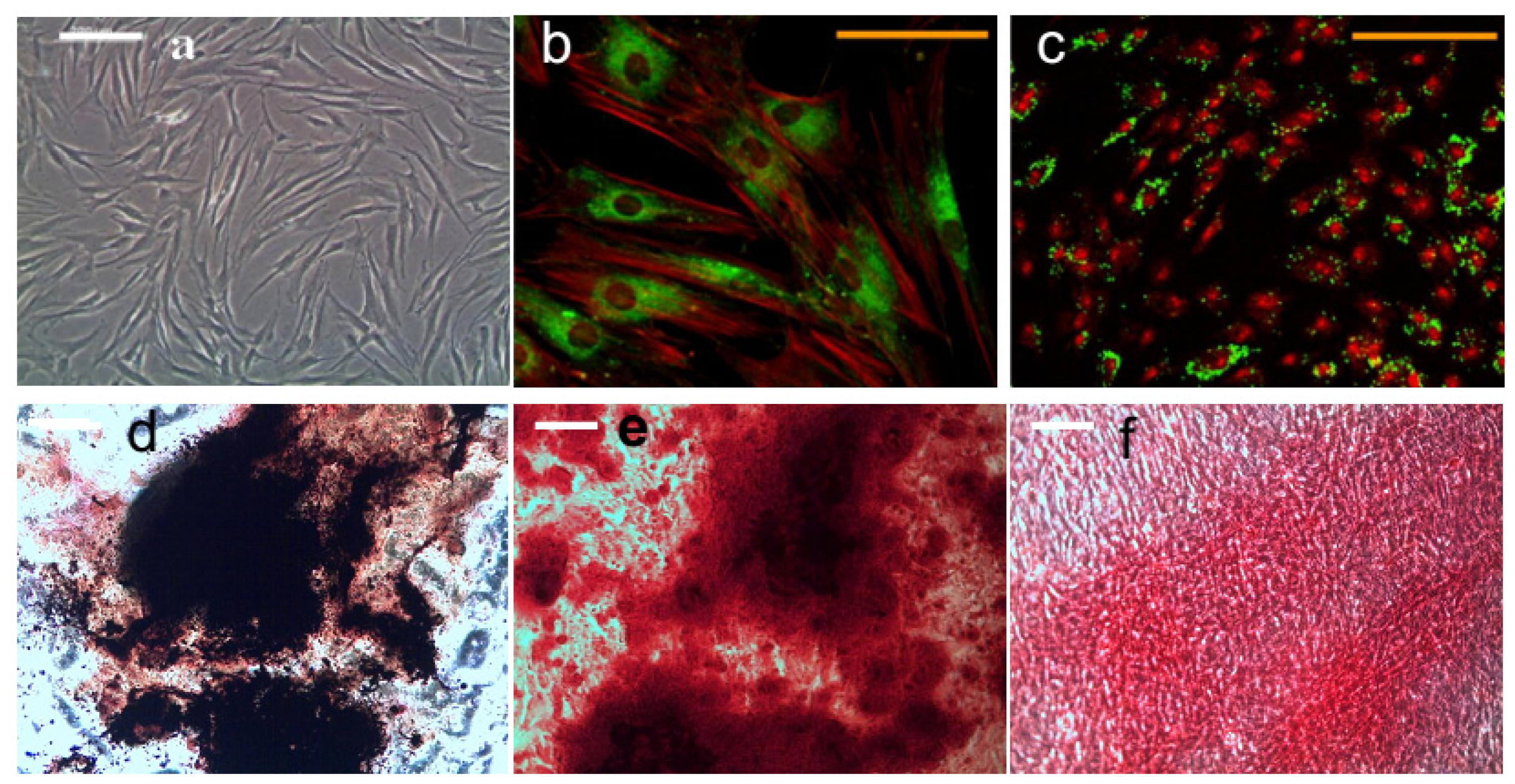
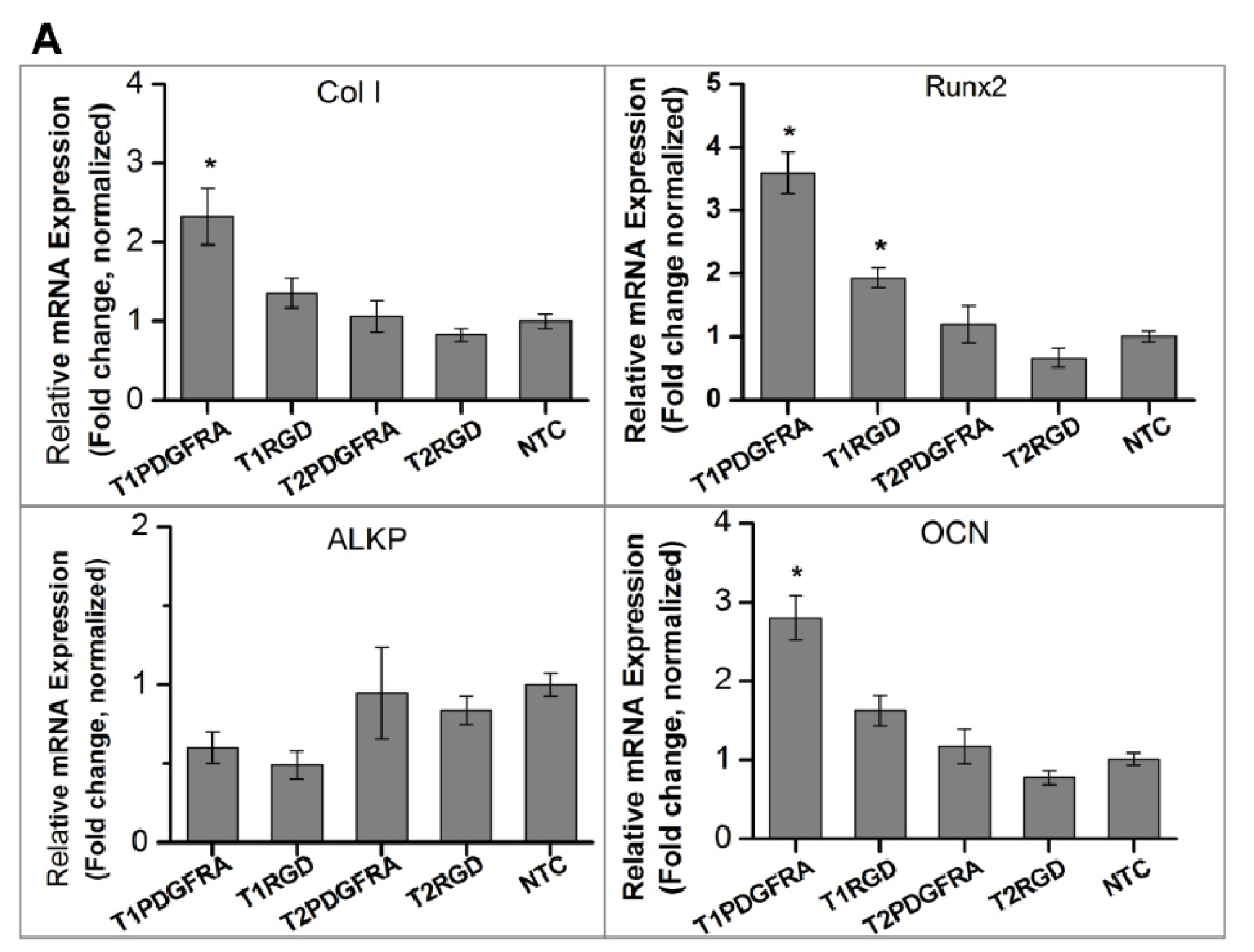
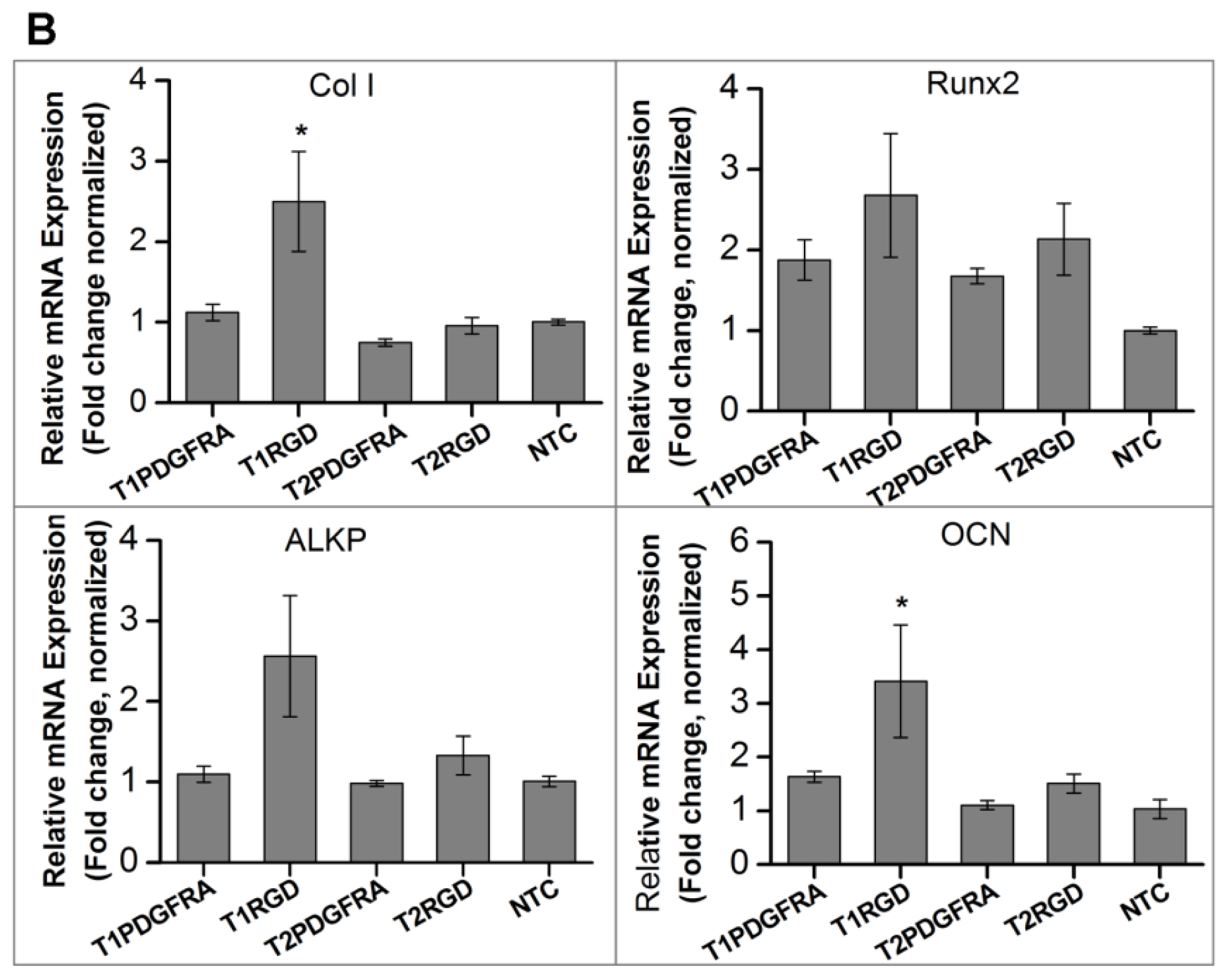
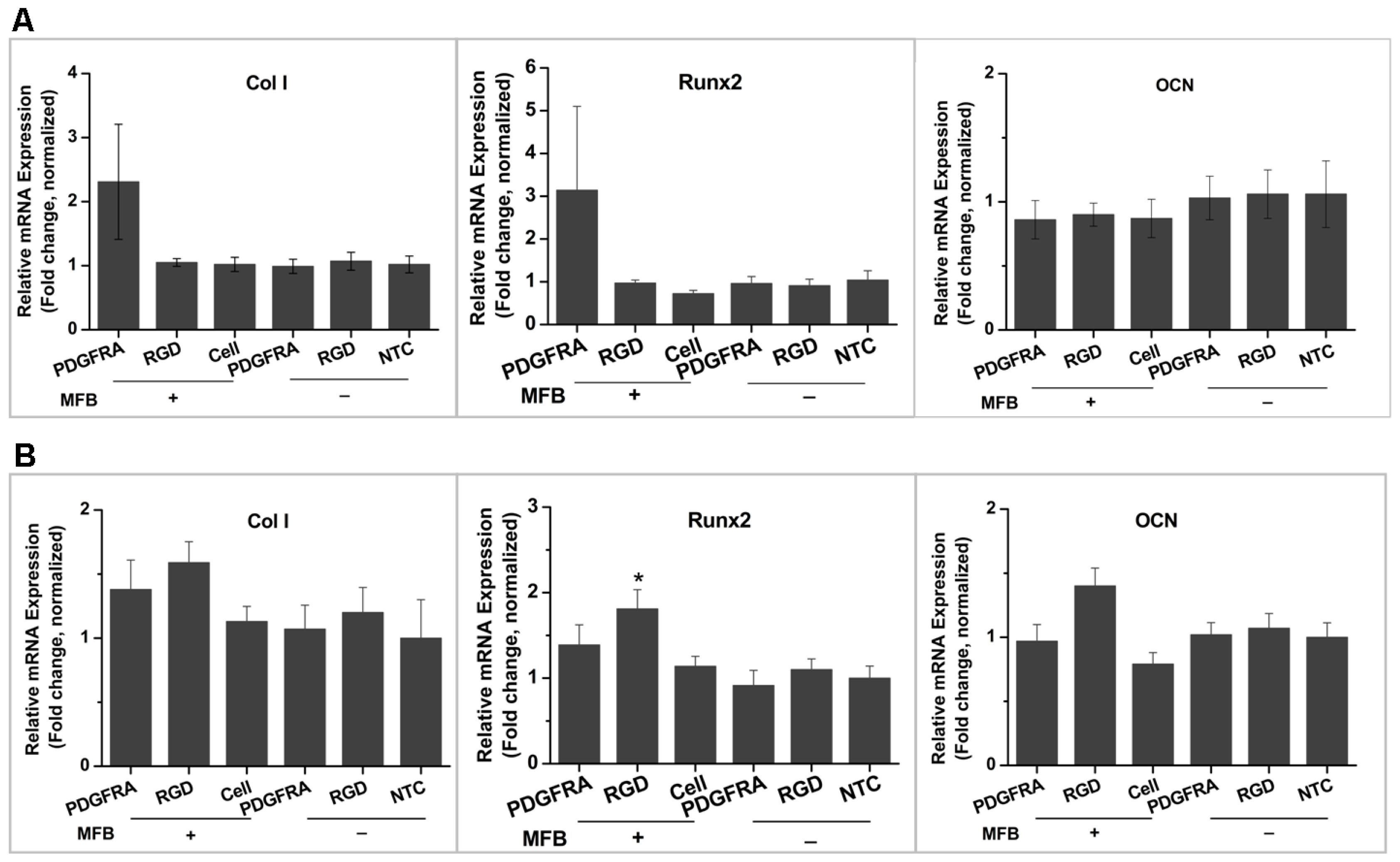
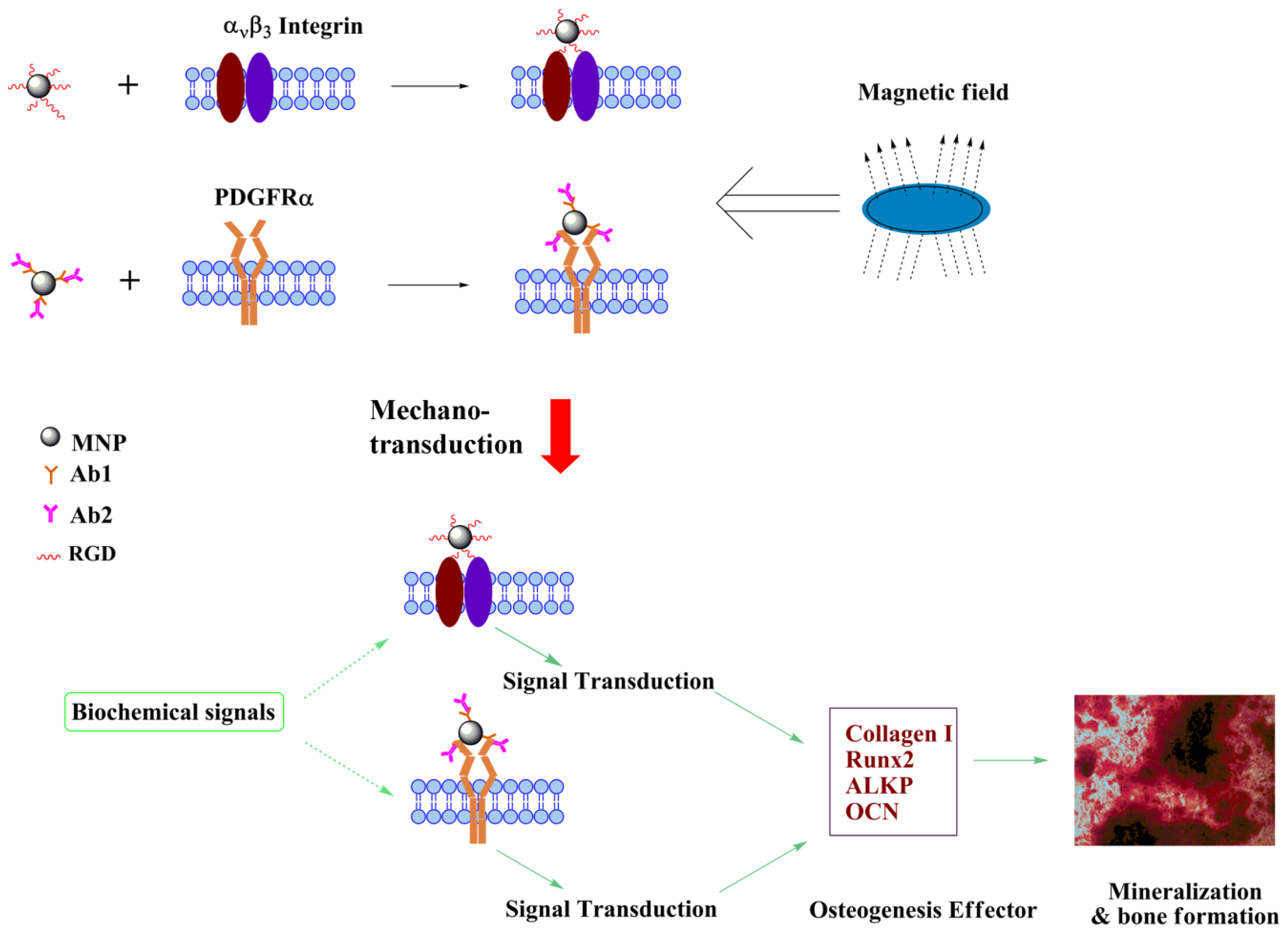
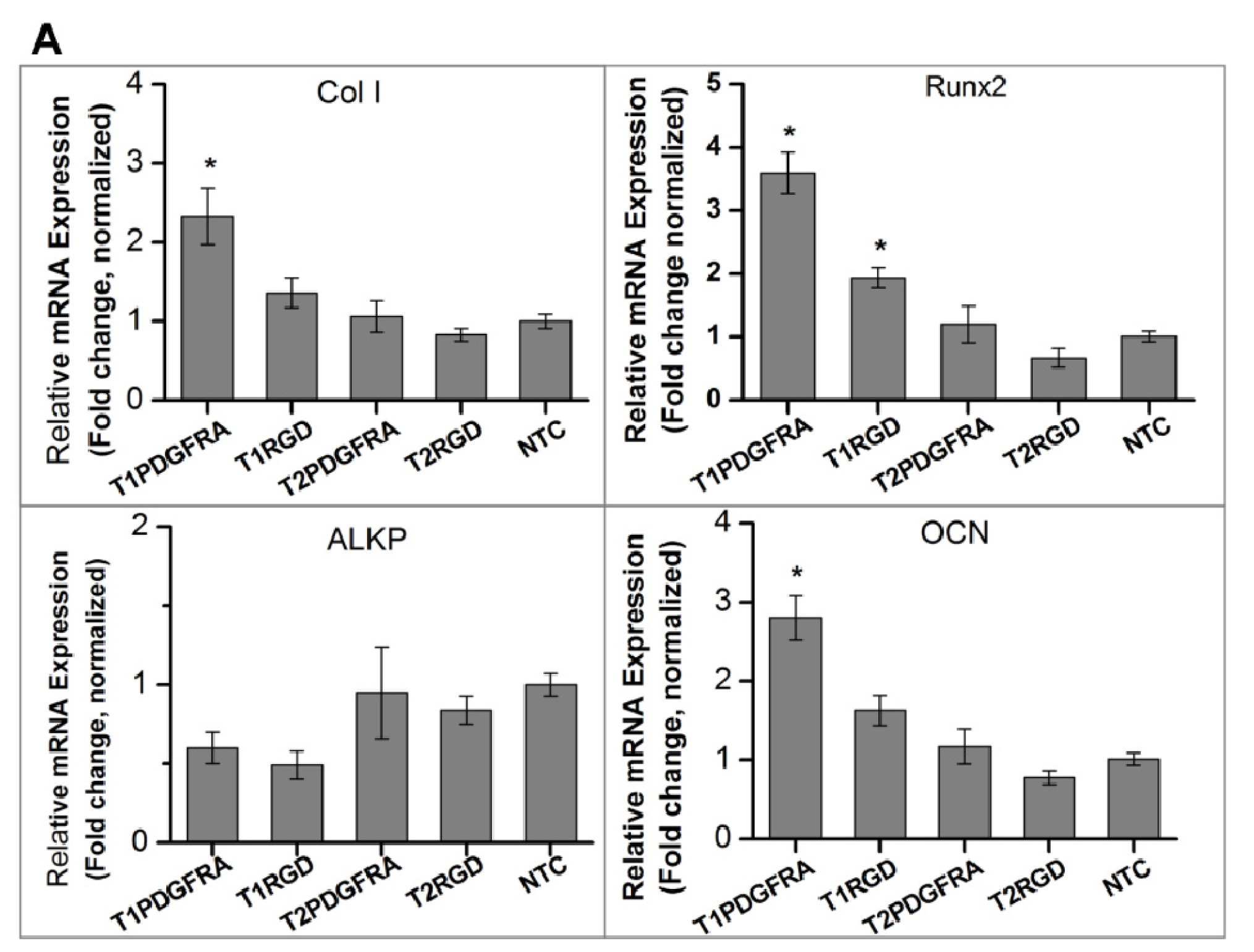
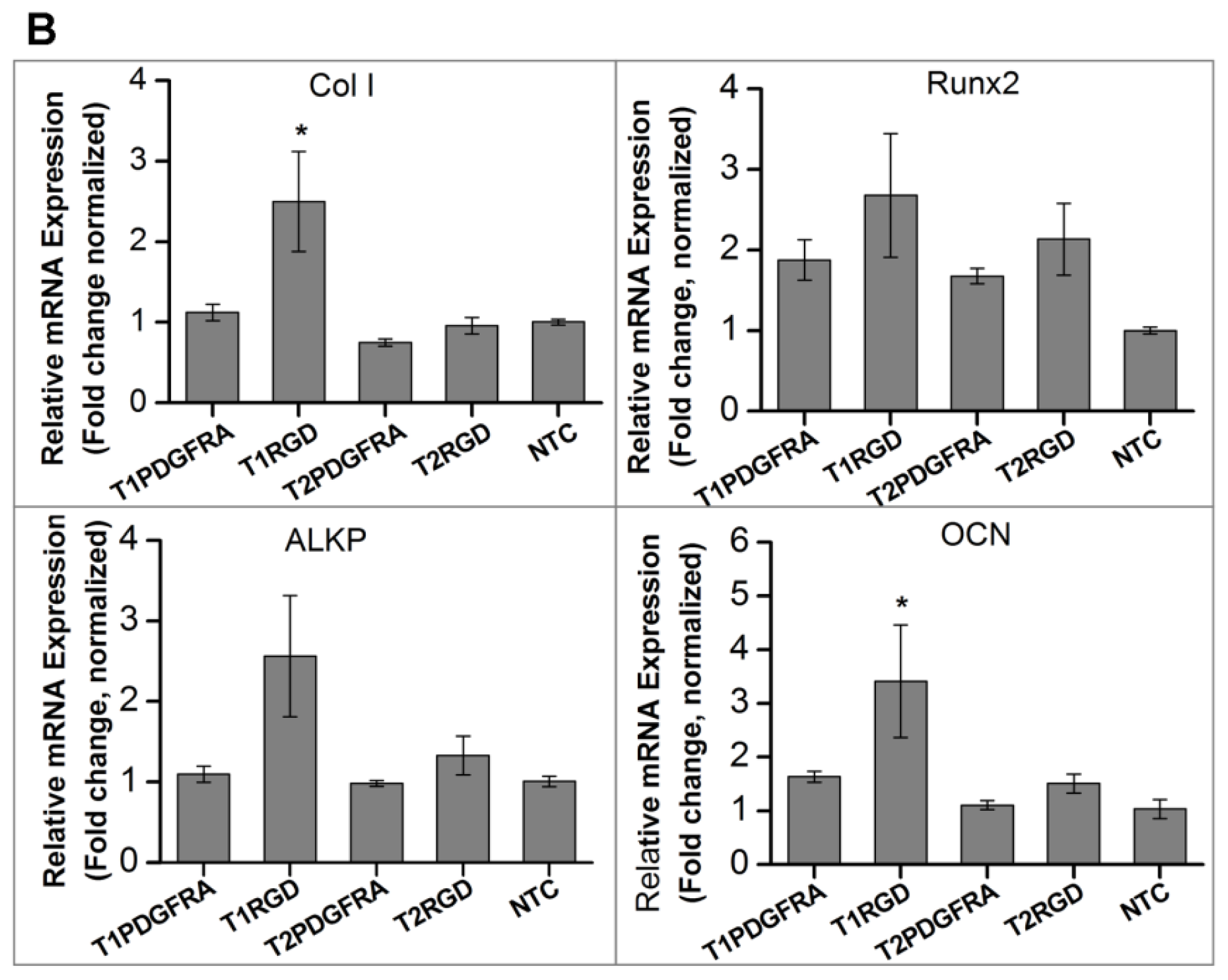
2.1. Magneto-Mechanical Stimulation Coupled with Biochemical Factors Directs the Osteogenic Differentiation of hMSCs
2.2. PDGFR and Osteogenesis of hMSCs
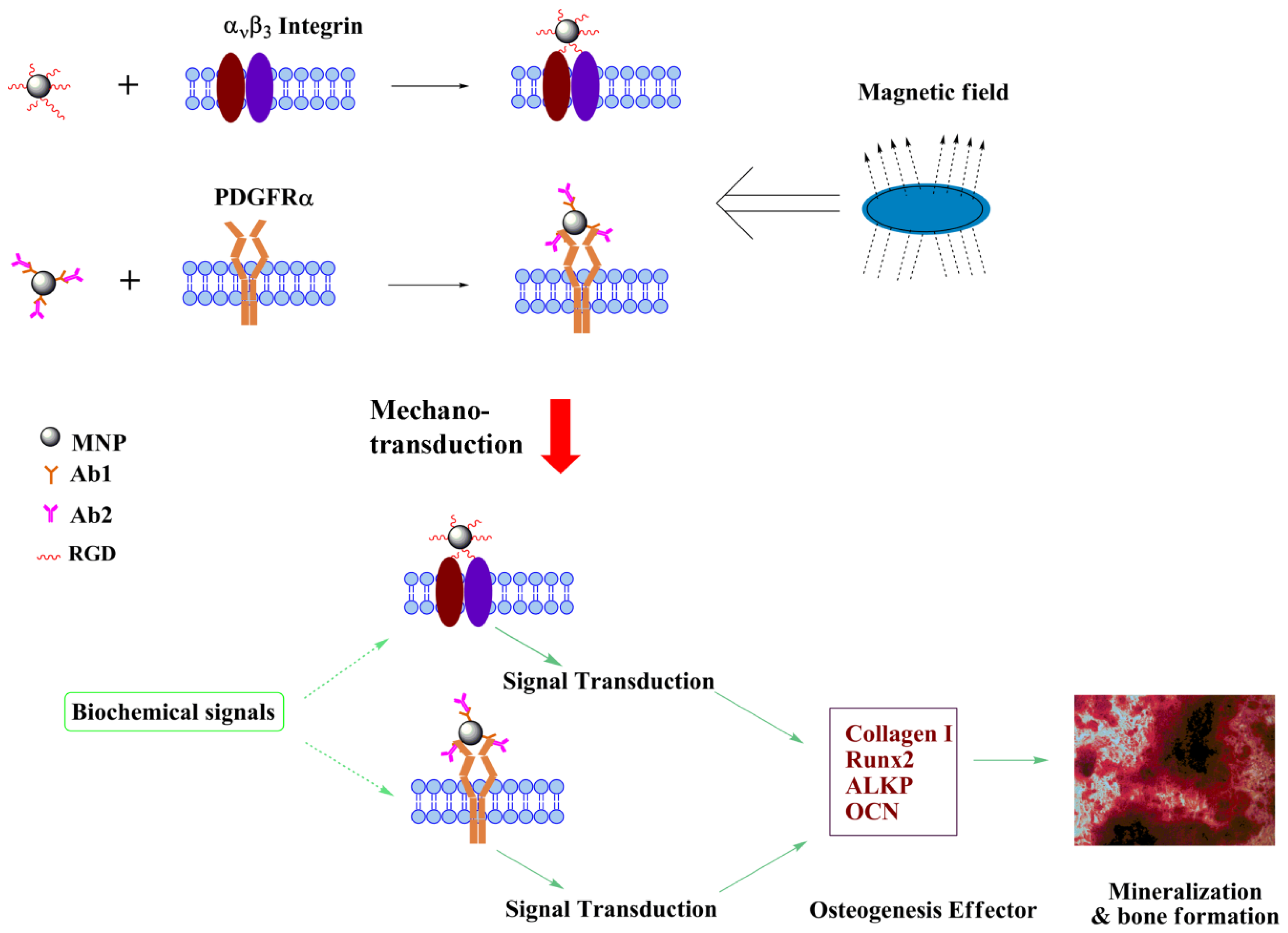
2.3. Proposed Effect of Combinational Temporal Factors and Targeting Sites of Magneto-Mechanical Stimulation on Osteogenic Induction of hMSCs
3. Experimental Section
3.1. Cell Culture
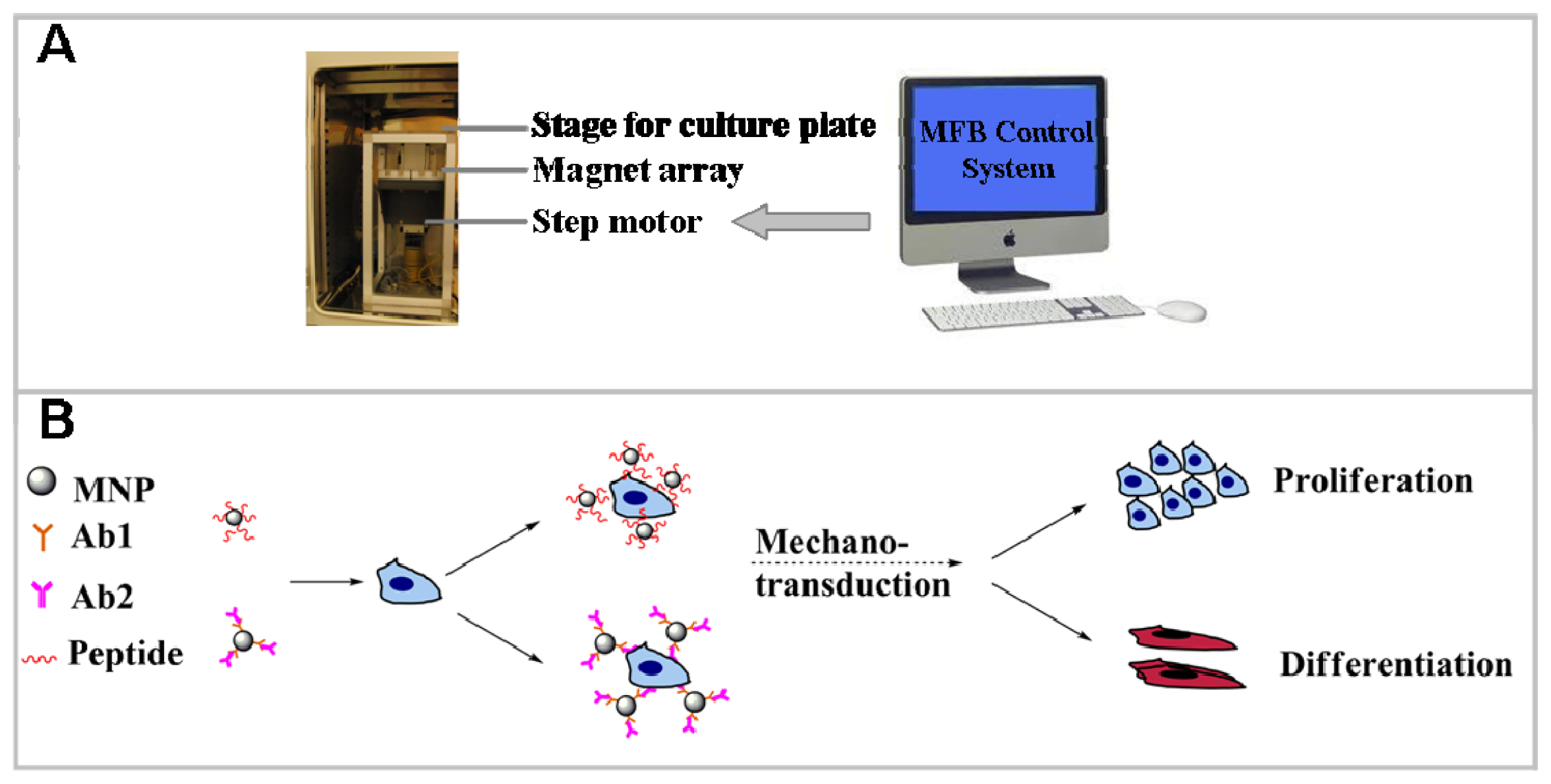
3.2. Magnetic Particle Conjugation
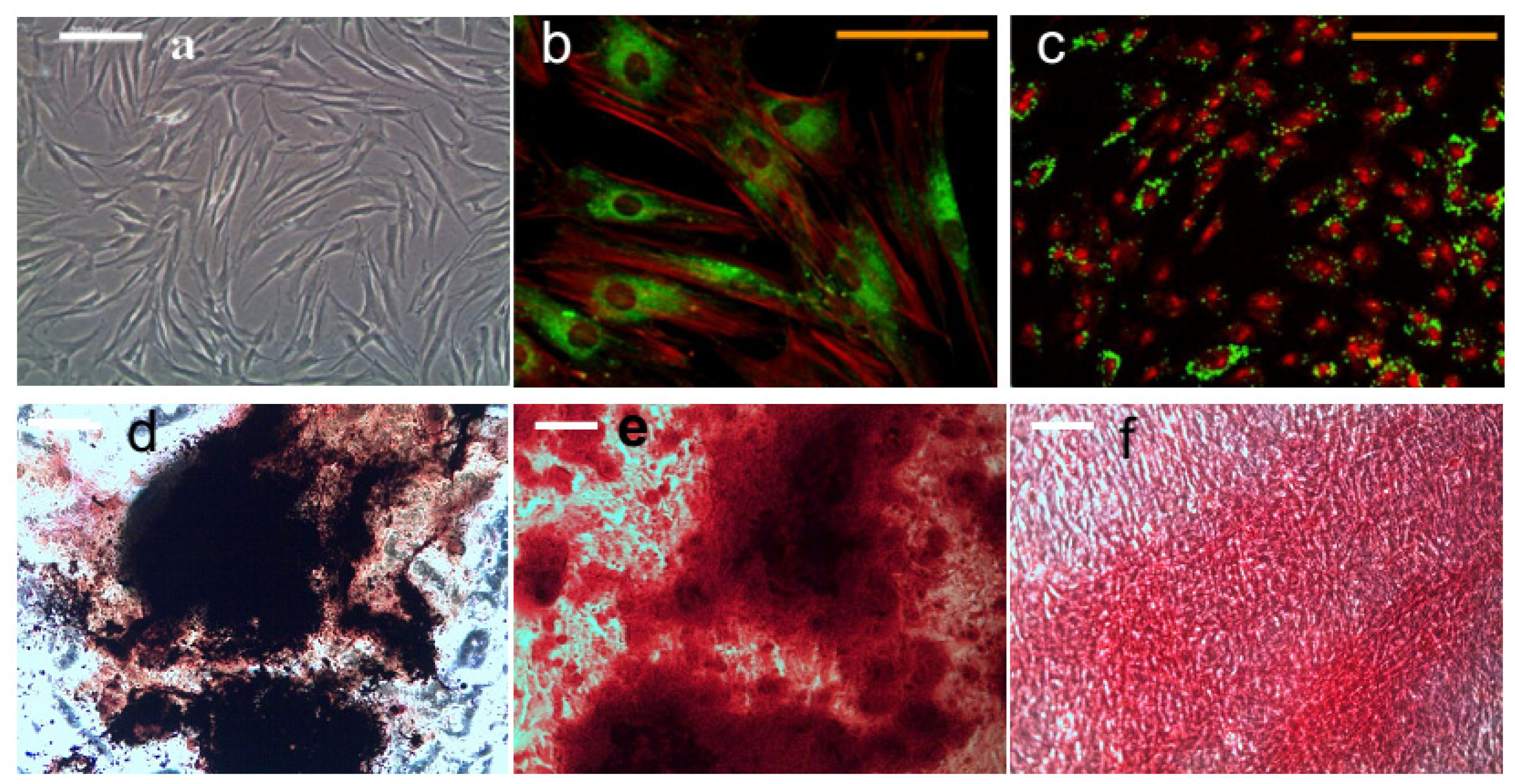
3.3. Visualization of MNP Targeting Cell Membrane
3.4. Application of MFB Technique in Osteogenesis of hMSC
3.5. Investigation of Temporal Effects of MFB on Osteogenesis of hMSCs
3.6. qRT-PCR
3.7. Histology
3.7.1. Alkaline Phosphatase and Von Kossa Double Staining
3.7.2. Calcium Staining
3.7.3. Collagen Staining
3.8. Thermal Gravimetric Analysis
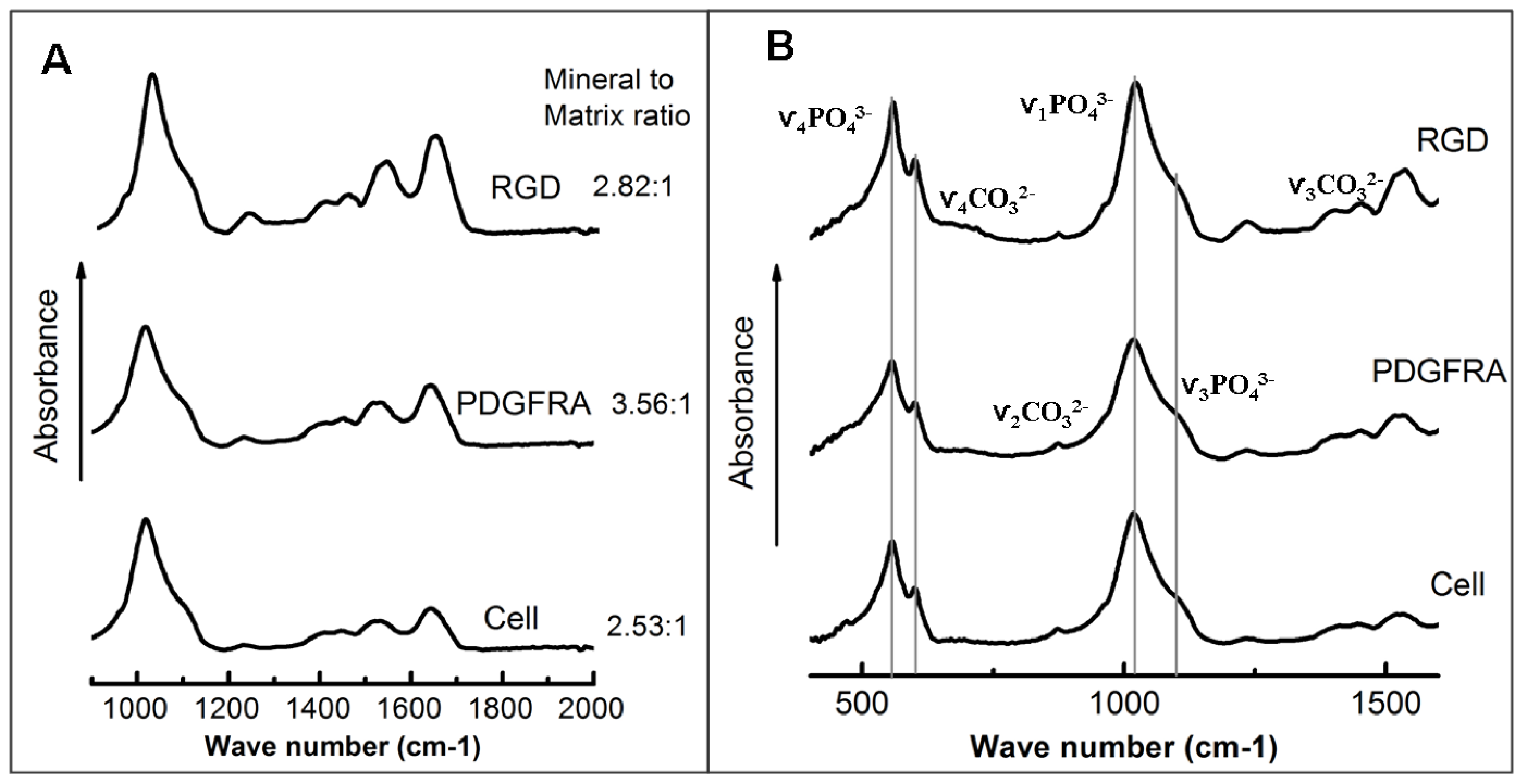
3.9. FTIR
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Langer, R.; Vacanti, J.P. Tissue engineering. Science 1993, 260, 920–926. [Google Scholar]
- Alsberg, E.; Feinstein, E.; Joy, M.P.; Prentiss, M.; Ingber, D.E. Magnetically-guided self-assembly of fibrin matrices with ordered nano-scale structure for tissue engineering. Tissue Eng 2006, 12, 3247–3256. [Google Scholar]
- Marolt, D.; Knezevic, M.; Vunjak-Novakovic, G. Bone tissue engineering with human stem cells. Stem Cell Res. Ther 2010, 1, 10. [Google Scholar]
- Baas, E.; Kuiper, J.H.; Yang, Y.; Wood, M.A.; El Haj, A.J. In vitro bone growth responds to local mechanical strain in three-dimensional polymer scaffolds. J. Biomech 2010, 43, 733–739. [Google Scholar]
- Cartmell, S.; El Haj, A.J. Mechanical Bioreactors for Tissue Engineering. In Bioreactors for Tissue Engineering, Principles, Design and Operation; Chaudhuri, J.B., Rubeai, M.A., Eds.; Springer: Berlin, Germany, 2005; pp. 193–208. [Google Scholar]
- Yamaguchi-Sekino, S.; Sekino, M.; Ueno, S. Biological effects of electromagnetic fields and recently updated safety guidelines for strong static magnetic fields. Magn. Reson. Med. Sci 2011, 10, 1–10. [Google Scholar]
- Kang, K.S.; Hong, J.M.; Kang, J.A.; Rhie, J.W.; Jeong, Y.H.; Cho, D.W. Regulation of osteogenic differentiation of human adipose-derived stem cells by controlling electromagnetic field conditions. Exp. Mol. Med 2013, 45, e6. [Google Scholar]
- Lewin, M.; Carlesso, N.; Tung, C.H.; Tang, X.W.; Cory, D.; Scadden, D.T.; Weissleder, R. Tat peptide-derivatized magnetic nanoparticles allow in vivo tracking and recovery of progenitor cells. Nat. Biotechnol 2000, 18, 410–414. [Google Scholar]
- Shimizu, K.; Ito, A.; Yoshida, T.; Yamada, Y.; Ueda, M.; Honda, H. Bone tissue engineering with human mesenchymal stem cell sheets constructed using magnetite nanoparticles and magnetic force. J. Biomed. Mater. Res. B 2007, 82, 471–480. [Google Scholar]
- Yamamoto, Y.; Ito, A.; Fujita, H.; Nagamori, E.; Kawabe, Y.; Kamihira, M. Functional evaluation of artificial skeletal muscle tissue constructs fabricated by a magnetic force-based tissue engineering technique. Tissue Eng. Part A 2011, 17, 107–114. [Google Scholar]
- Fayol, D.; Frasca, G.; Le Visage, C.; Gazeau, F.; Luciani, N.; Wilhelm, C. Use of magnetic forces to promote stem cell aggregation during differentiation, and cartilage tissue modeling. Adv. Mater 2013, 25, 2611–2616. [Google Scholar]
- Pankhurst, Q.A.; Thanh, N.T.K.; Jones, S.K.; Dobson, J. Progress in applications of magnetic nanoparticles in biomedicine. J. Phys. D 2009, 42, 224001. [Google Scholar]
- Cartmell, S.; Dobson, J.; Verschueren, S.; El Haj, A. Development of magnetic particle techniques for long-term culture of bone cells with intermittent mechanical activation. IEEE Trans. Nanobioscience 2002, 1, 92–97. [Google Scholar]
- Hu, B.; Dobson, J.; El Haj, A.J. Control of Smooth Muscle α-actin (SMA) up-regulation in HBMSCs using remote magnetic particle mechano-activation. Nanomedicine 2013. [Google Scholar] [CrossRef]
- Dobson, J.; Cartmell, S.H.; Keramane, A.; El Haj, A.J. A magnetic force mechanical conditioning bioreactor for tissue engineering, stem cell conditioning and dynamic in vitro screening. IEEE Trans. Nanobioscience 2006, 5, 173–177. [Google Scholar]
- Hughes, S.; McBain, S.; Dobson, J.; El Haj, A.J. Selective activation of mechanosensitive ion channels using magnetic particles. J. R. Soc. Interface 2008, 5, 855–863. [Google Scholar]
- Kanczler, J.M.; Sura, H.S.; Magnay, J.; Green, D.; Oreffo, R.O.; Dobson, J.P.; El Haj, A.J. Controlled differentiation of human bone marrow stromal cells using magnetic nanoparticle technology. Tissue Eng. Part A 2010, 16, 3241–3250. [Google Scholar]
- Thomas, G.P.; El Haj, A.J. Bone marrow stromal cells are load responsive in vitro. Calcif. Tissue Int 1996, 58, 101–108. [Google Scholar]
- Nii, M.; Lai, J.H.; Keeney, M.; Han, L.H.; Behn, A.; Imanbayev, G.; Yang, F. The effects of interactive mechanical and biochemical niche signaling on osteogenic differentiation of adipose-derived stem cells using combinatorial hydrogels. Acta Biomater 2013, 9, 5475–5483. [Google Scholar]
- Dobson, J. Remote control of cellular behaviour with magnetic nanoparticles. Nat. Nanotechnol 2008, 3, 139–143. [Google Scholar]
- Dobson, J. Cancer therapy: A twist on tumour targeting. Nat. Mater 2010, 9, 95–96. [Google Scholar]
- Chaudhary, L.R.; Hofmeister, A.M.; Hruska, K.A. Differential growth factor control of bone formation through osteoprogenitor differentiation. Bone 2004, 34, 402–411. [Google Scholar]
- Mitlak, B.H.; Finkelman, R.D.; Hill, E.L.; Li, J.; Martin, B.; Smith, T.; D’Andrea, M.; Antoniades, H.N.; Lynch, S.E. The effect of systemically administered PDGF-BB on the rodent skeleton. J. Bone Miner. Res 1996, 11, 238–247. [Google Scholar]
- Nash, T.J.; Howlett, C.R.; Martin, C.; Steele, J.; Johnson, K.A.; Hicklin, D.J. Effect of platelet-derived growth factor on tibial osteotomies in rabbits. Bone 1994, 15, 203–208. [Google Scholar]
- Vikjaer, D.; Blom, S.; Hjørting-Hansen, E.; Pinholt, E.M. Effect of platelet-derived growth factor-BB on bone formation in calvarial defects: An experimental study in rabbits. Eur. J. Oral Sci 1997, 105, 59–66. [Google Scholar]
- Ranly, D.M.; McMillan, J.; Keller, T.; Lohmann, C.H.; Meunch, T.; Cochran, D.L.; Schwartz, Z.; Boyan, B.D. Platelet-derived growth factor inhibits demineralised bone matrix-induced intramuscular cartilage and bone formation. A study of immuno compromised mice. J. Bone Joint Surg. Am 2005, 87, 2052–2064. [Google Scholar]
- Fierro, F.; Illmer, T.; Jing, D.; Schleyer, E.; Ehninger, G.; Boxberger, S.; Bornhäuser, M. Inhibition of platelet-derived growth factor receptorbeta by imatinib mesylate suppresses proliferation and alters differentiation of human mesenchymal stem cells in vitro. Cell Prolif 2007, 40, 355–366. [Google Scholar]
- Grey, A.; O’Sullivan, S.; Reid, I.R.; Browett, P. Imatinib mesylate, increased bone formation, and secondary hyperparathyroidism. N. Engl. J. Med 2006, 355, 2494–2495. [Google Scholar]
- O’Sullivan, S.; Naot, D.; Callon, K.; Porteous, F.; Horne, A.; Wattie, D.; Watson, M.; Cornish, J.; Browett, P.; Grey, A. Imatinib promotes osteoblast differentiation by inhibiting PDGFR signaling and inhibits osteoclastogenesis by both direct and stromal cell-dependent mechanisms. J. Bone Miner. Res 2007, 22, 1679–1689. [Google Scholar]
- Fitter, S.; Dewar, A.L.; Kostakis, P.; To, L.B.; Hughes, T.P.; Roberts, M.M.; Lynch, K.; Vernon-Roberts, B.; Zannettino, A.C. Long-term imatinib therapy promotes bone formation in CML patients. Blood 2008, 111, 2538–2547. [Google Scholar]
- Karp, J.M.; Ferreira, L.S.; Khademhosseini, A.; Kwon, A.H.; Yeh, J.; Langer, R.S. Cultivation of human embryonic stem cells without the embryoid body step enhances osteogenesis in vitro. Stem Cells 2006, 24, 835–843. [Google Scholar]
- Boskey, A.L.; Mendelsohn, R. Infrared spectroscopic characterization of mineralized tissues. Vib. Spectrosc 2005, 38, 107–114. [Google Scholar]
- Mendelsohn, R.; Paschalis, E.P.; Boskey, A.L. Infrared spectroscopy, microscopy, and microscopic imaging of mineralizing tissues. Spectra-structure correlations from human iliac crest biopsies. J. Biomed. Opt 1999, 4, 14–21. [Google Scholar]
- Paschalis, E.P.; Verdelis, K.; Mendelsohn, R.; Boskey, A.; Yamauchi, M. Spectroscopic determination of collagen cross-links in bone. J. Bone Miner. Res 2001, 16, 1821–1828. [Google Scholar]
- Komori, T. Runx2, a multifunctional transcription factor in skeletal development. J. Cell. Biochem 2002, 87, 1–8. [Google Scholar]
- Graham, S.; Leonidou, A.; Lester, M.; Heliotis, M.; Mantalaris, A.; Tsiridis, E. Investigating the role of PDGF as a potential drug therapy in bone formation and fracture healing. Expert Opin. Investig. Drugs 2009, 18, 1633–1654. [Google Scholar]
- Dobrowolski, R.; de Robertis, E.M. Endocytic control of growth factor signalling: Multivesicular bodies as signalling organelles. Nat. Rev. Mol. Cell Biol 2011, 13, 53–60. [Google Scholar]
- Kulterer, B.; Friedl, G.; Jandrositz, A.; Sanchez-Cabo, F.; Prokesch, A.; Paar, C.; Scheideler, M.; Windhager, R.; Preisegger, K.-H.; Trajanoski, Z. Gene expression profiling of human mesenchymal stem cells derived from bone marrow during expansion and osteoblast differentiation. BMC Genomics 2007, 8, 70. [Google Scholar]
- Golub, E.E.; Boesze-Battaglia, K. The role of alkaline phosphatase in mineralization. Curr. Opin. Orthop 2007, 18, 444–448. [Google Scholar]
- Kaazempur Mofrad, M.R.; Abdul-Rahim, N.A.; Karcher, H.; Mack, P.J.; Yap, B.; Kamm, R.D. Exploring the molecular basis for mechanosensation, signal transduction, and cytoskeletal remodeling. Acta Biomater 2005, 1, 281–293. [Google Scholar]
- Pedersen, J.A.; Swartz, M.A. Mechanobiology in the third dimension. Ann. Biomed. Eng 2005, 33, 1469–1490. [Google Scholar]
- Wang, N.; Butler, J.P.; Ingber, D.E. Mechanotransduction across the cell surface and through the cytoskeleton. Science 1993, 260, 1124–1127. [Google Scholar]
- Bodine, P.V.; Trailsmith, M.; Komm, B.S. Development and characterization of a conditionally transformed adult human osteoblastic cell line. J. Bone Miner. Res 1996, 11, 806–819. [Google Scholar]
- Junqueira, L.C.; Bignolas, G.; Brentani, R.R. Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histochem. J 1979, 11, 447–455. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Mineral/matrix ratio | 1660/1690 ratio | 1020/1030 ratio |
|---|---|---|---|
| PDGFRα treatment | 3.56 ± 0.52 | 2.48 ± 0.02 | 1.08 ± 0.01 |
| RGD treatment | 2.82 ± 0.56 | 2.45 ± 0.08 | 0.07 ± 0.01 |
| Cell (nontreated) | 2.53 ± 0.44 | 2.55 ± 0.04 | 1.06 ± 0.06 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hu, B.; Haj, A.J.E.; Dobson, J. Receptor-Targeted, Magneto-Mechanical Stimulation of Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells. Int. J. Mol. Sci. 2013, 14, 19276-19293. https://doi.org/10.3390/ijms140919276
Hu B, Haj AJE, Dobson J. Receptor-Targeted, Magneto-Mechanical Stimulation of Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2013; 14(9):19276-19293. https://doi.org/10.3390/ijms140919276
Chicago/Turabian StyleHu, Bin, Alicia J El Haj, and Jon Dobson. 2013. "Receptor-Targeted, Magneto-Mechanical Stimulation of Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells" International Journal of Molecular Sciences 14, no. 9: 19276-19293. https://doi.org/10.3390/ijms140919276