Temperature-Responsive Poly(ε-caprolactone) Cell Culture Platform with Dynamically Tunable Nano-Roughness and Elasticity for Control of Myoblast Morphology



Abstract
:1. Introduction
2. Results and Discussion
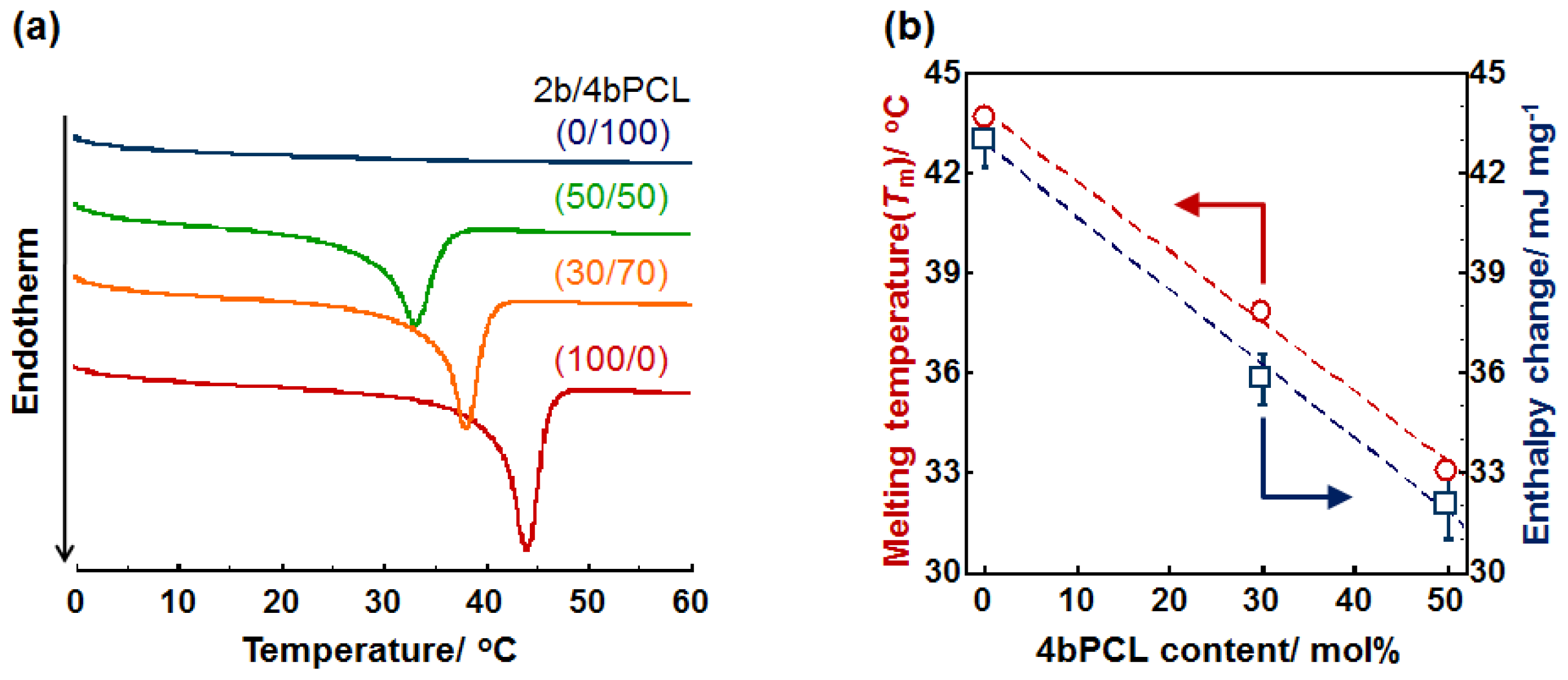
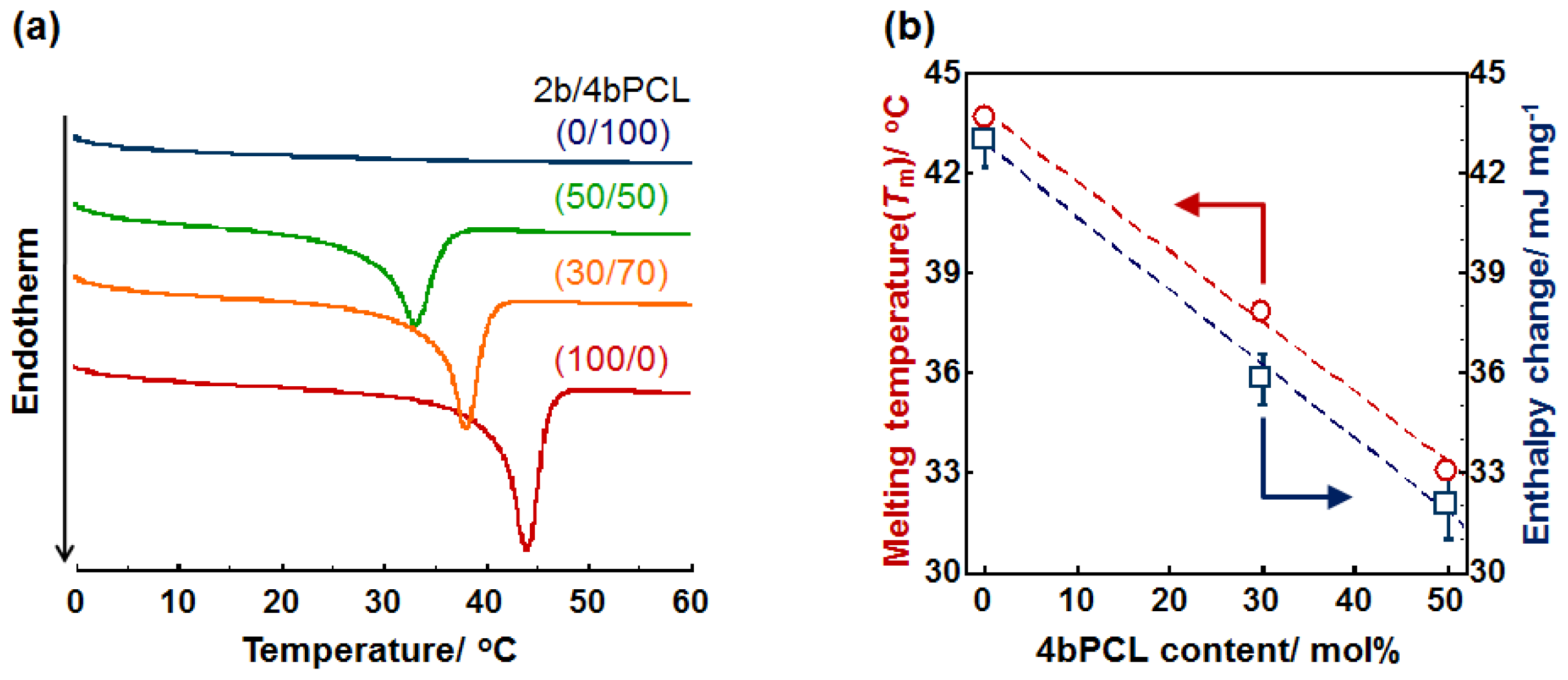
2.1. Crystal-Amorphous Transition Temperature
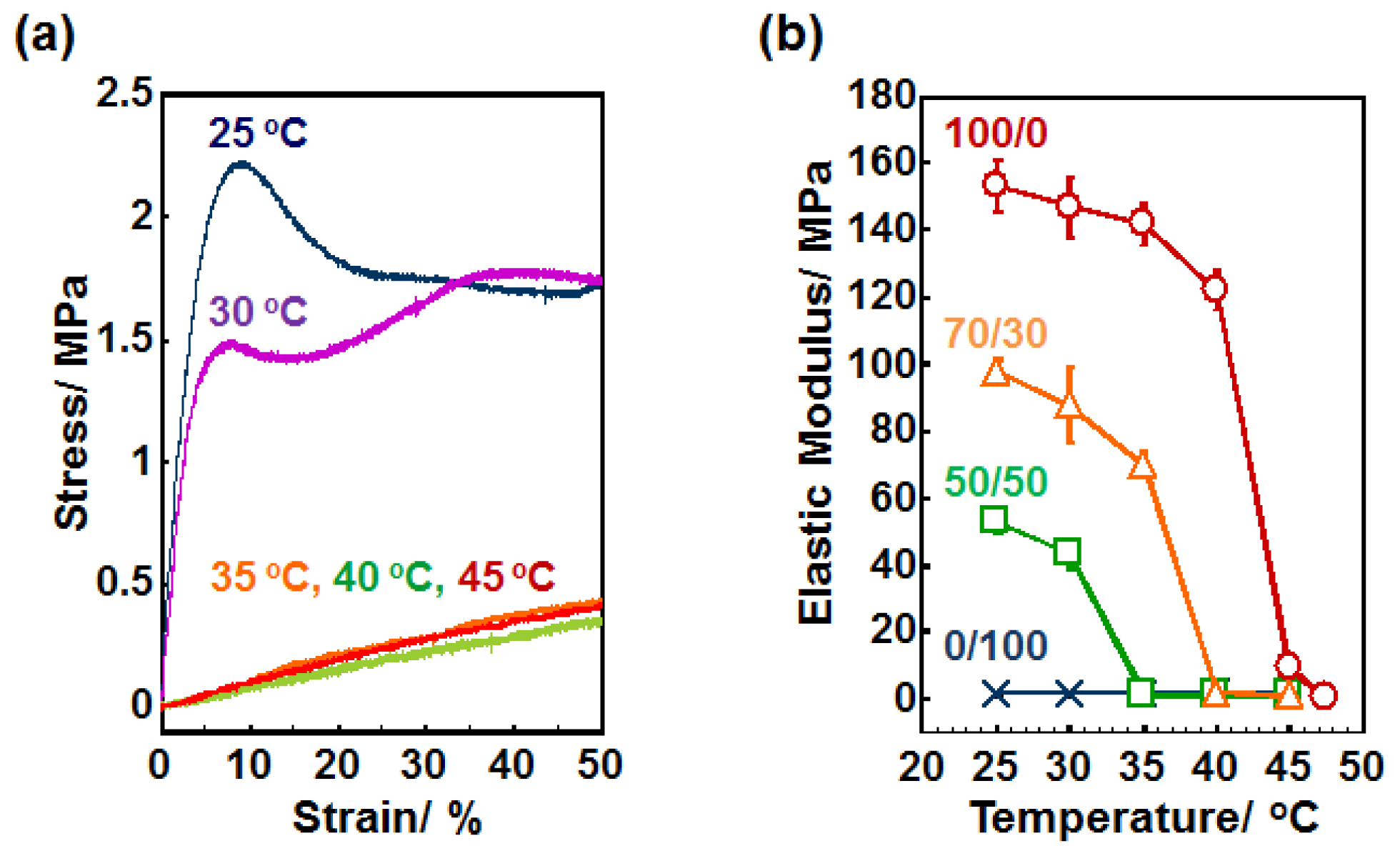
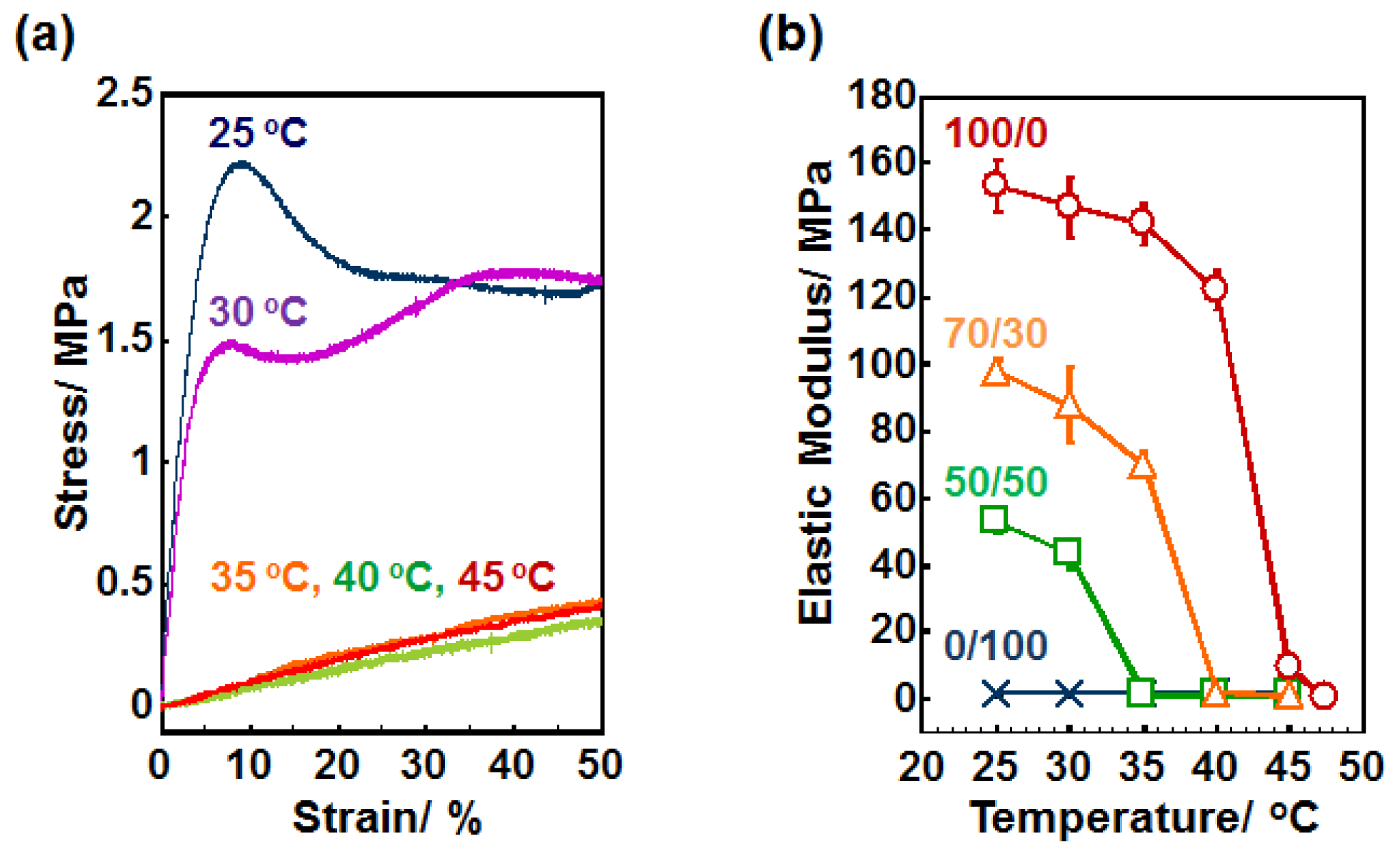
2.2. Elastic Modulus
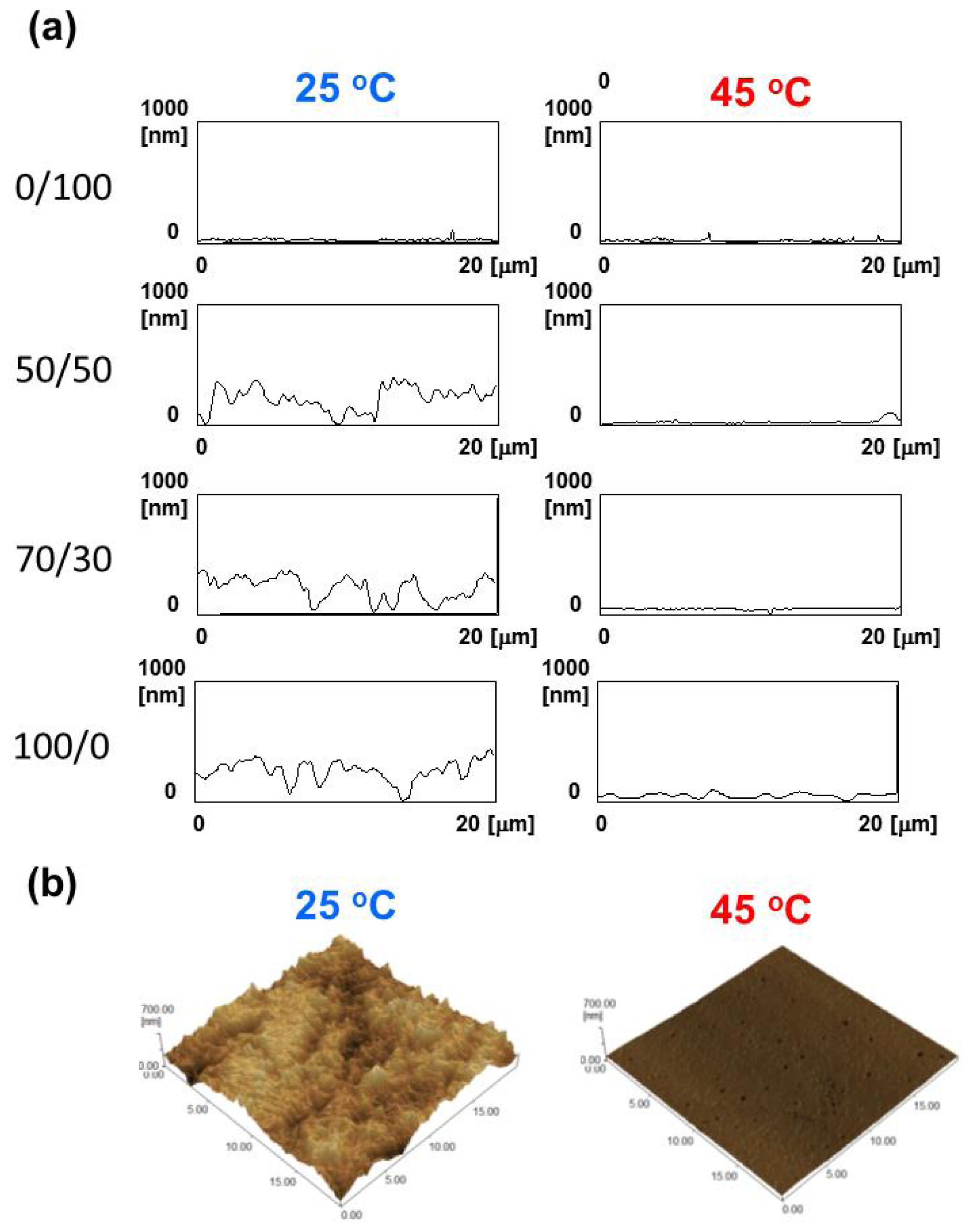
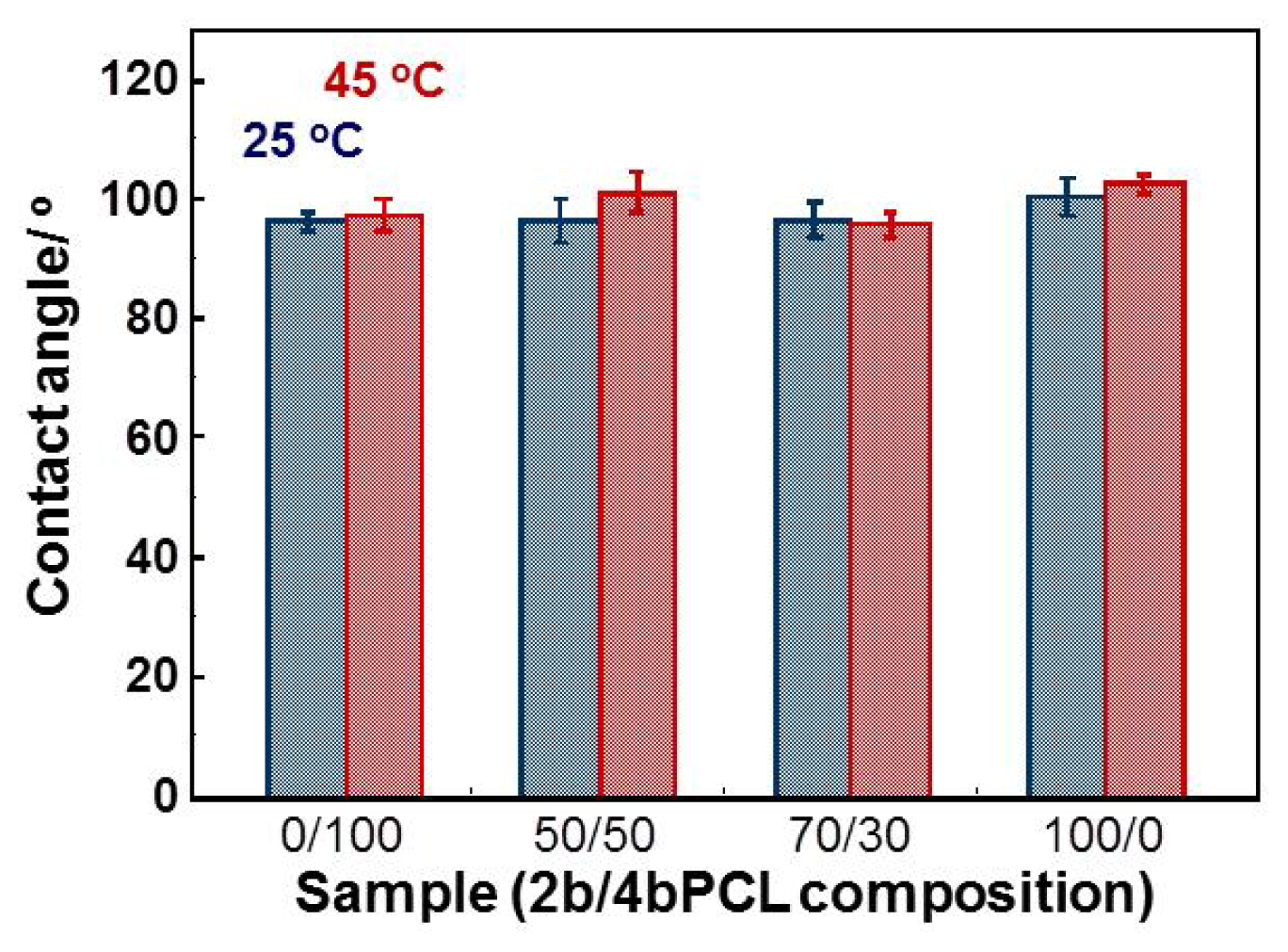
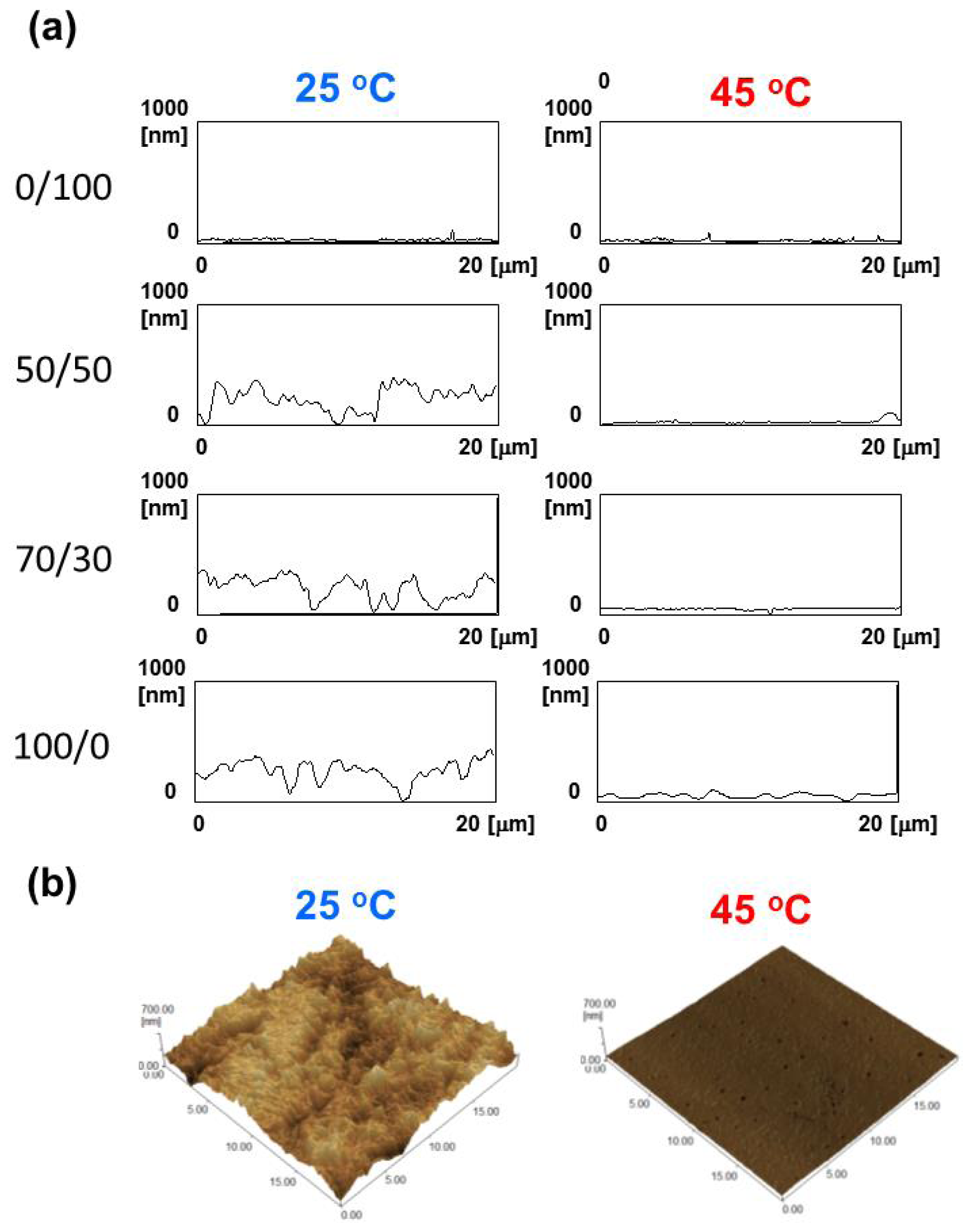
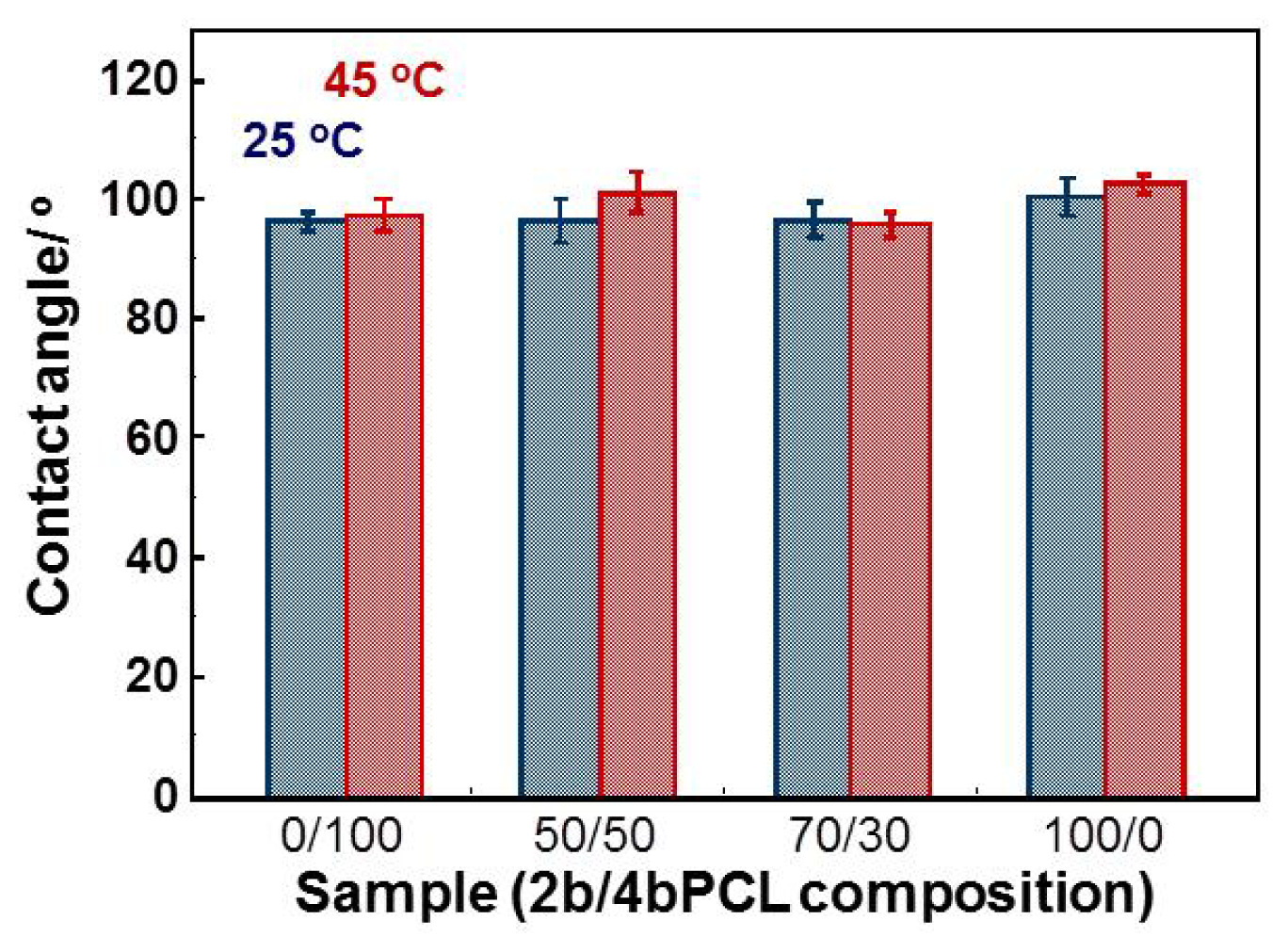
2.3. Surface Morphology and Wettability
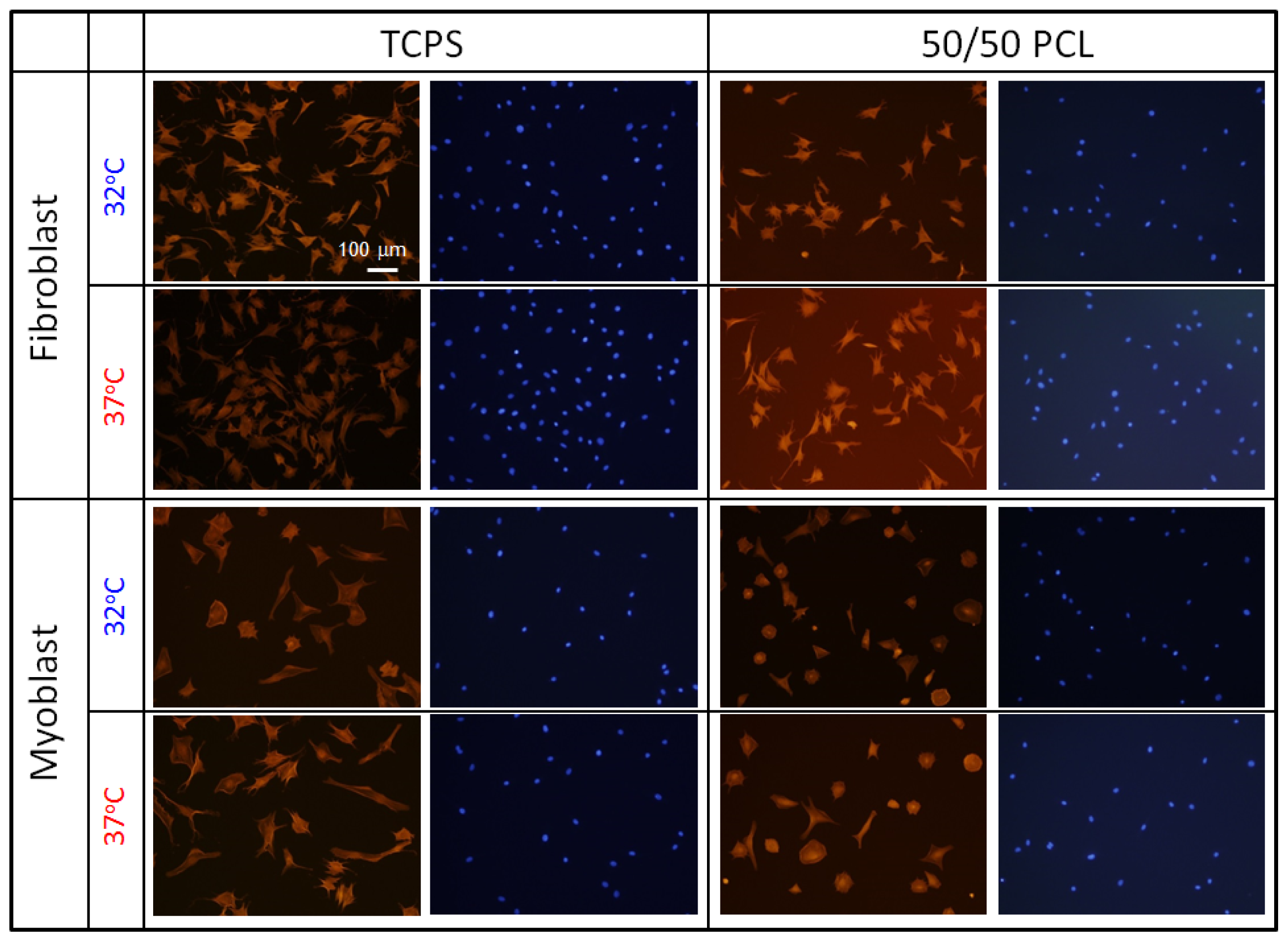
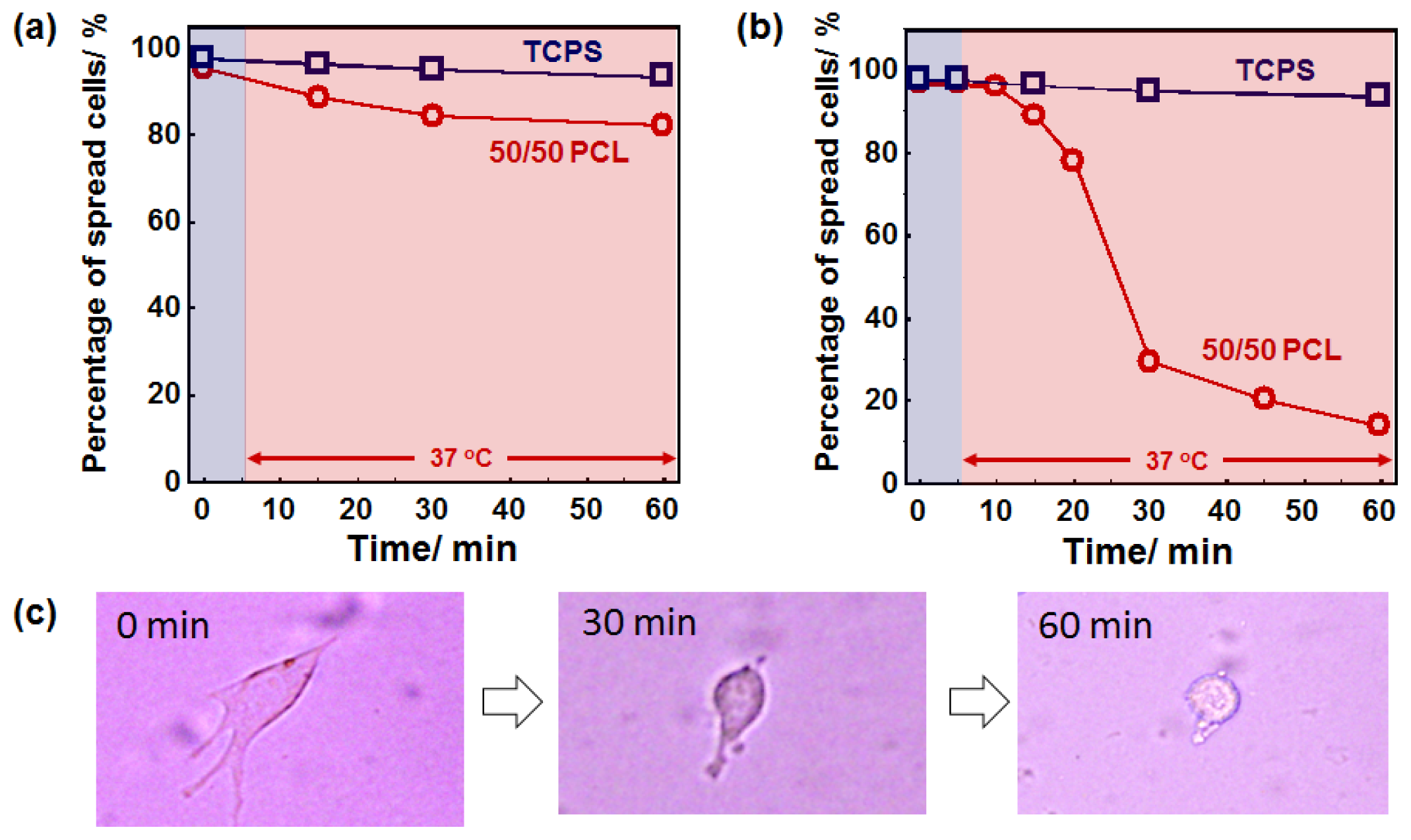
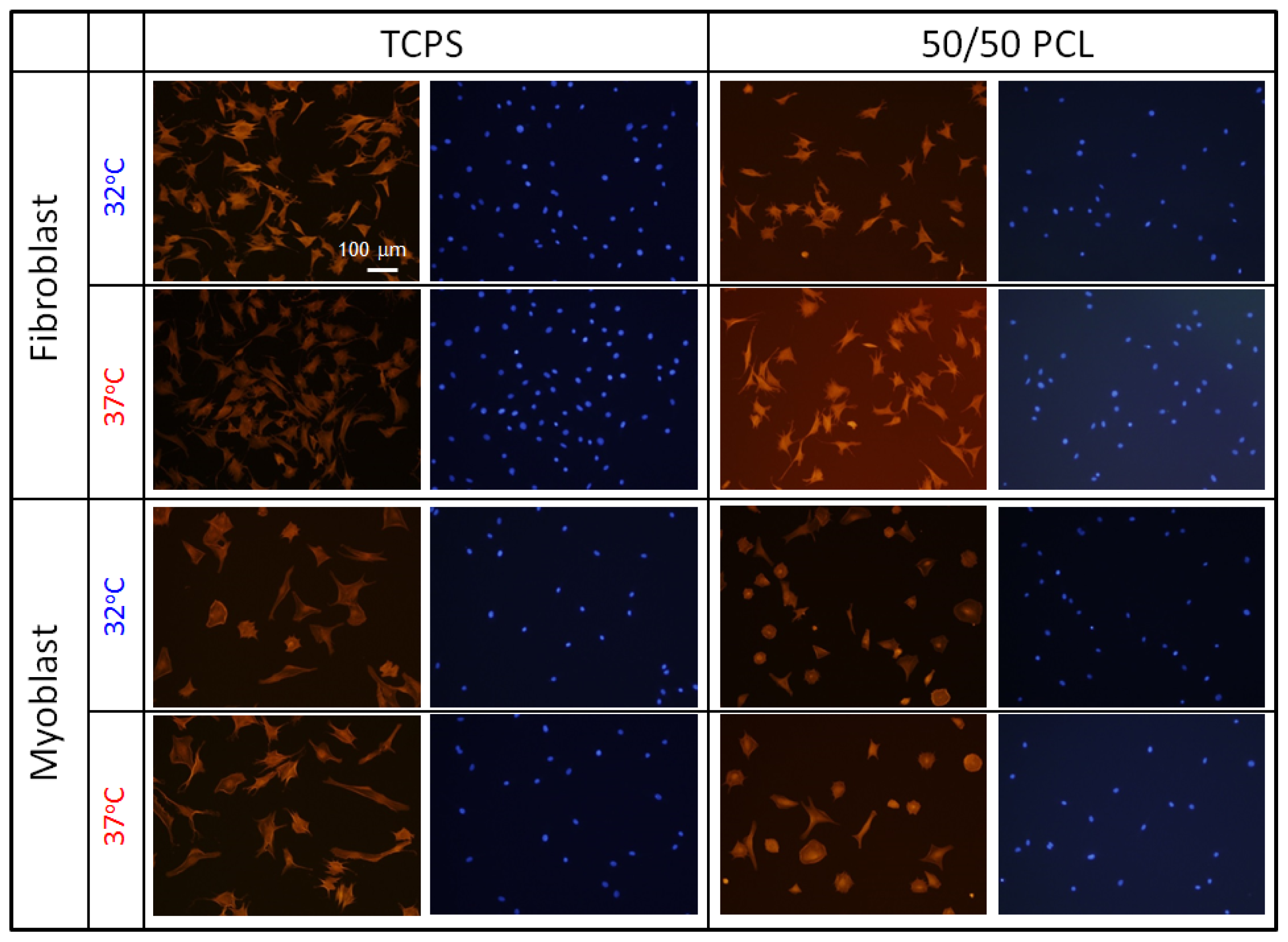
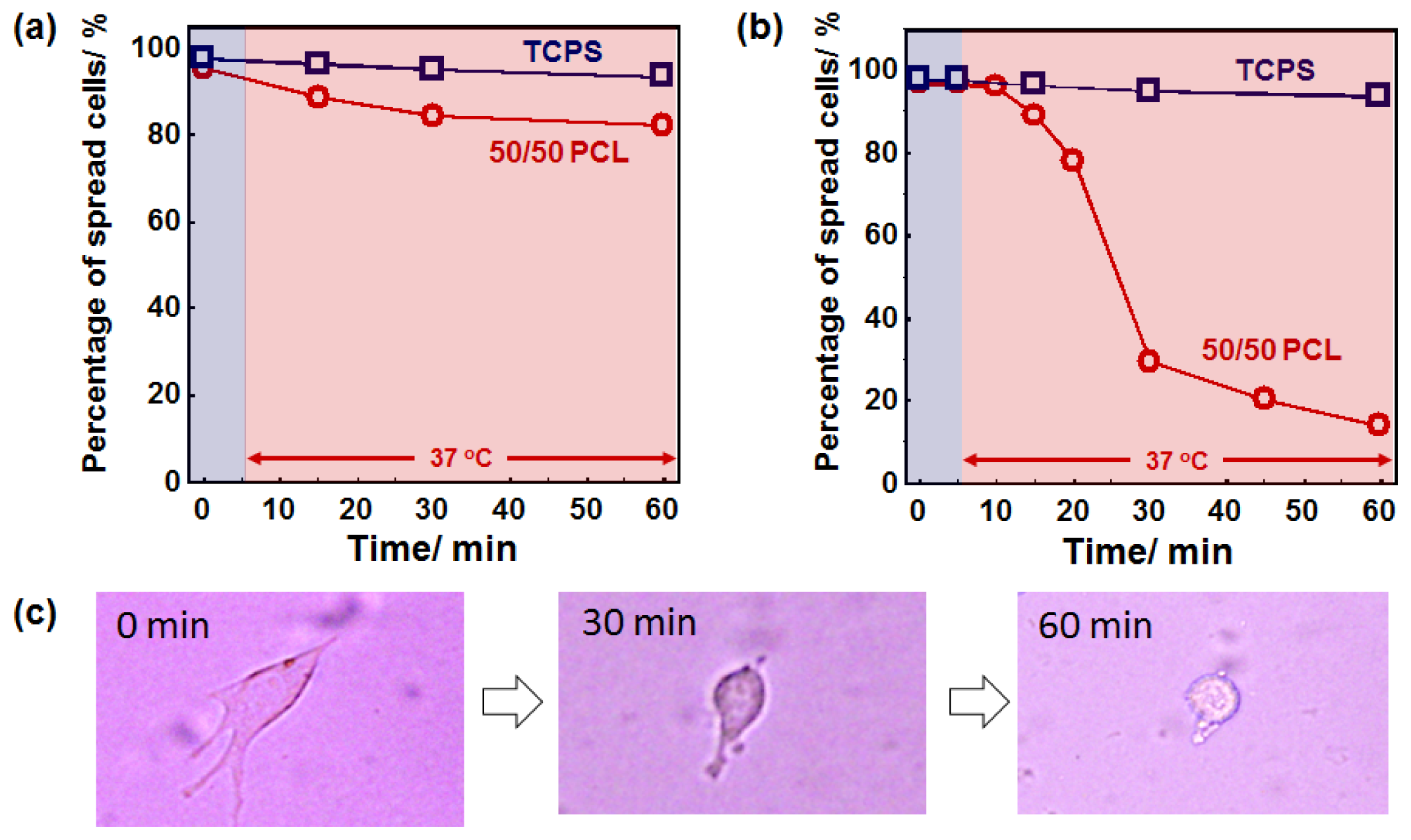
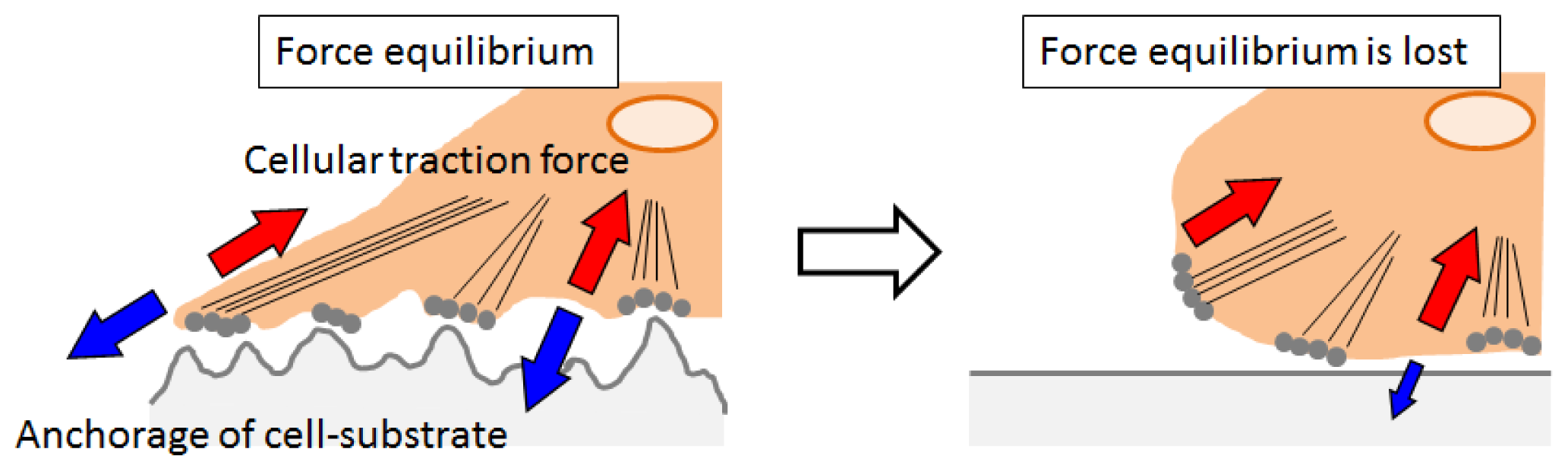
2.4. Cell Behavior
3. Experimental Section
3.1. Fabrication of Crosslinked PCL Films
3.2. Thermo-Mechanical Properties
3.3. Topographical Observation by Atomic Force Microscope (AFM)
3.4. Contact Angles
3.5. Cell Culture
4. Conclusions
Supplementary Information
ijms-15-01511-s001.pdfAcknowledgments
Conflicts of Interest
References
- Wolf, K.; Mazo, I.; Leung, H.; Engelke, K.; von Andrian, U.H.; Deryugina, E.I.; Strongin, A.Y.; Bröcker, E.-B.; Friedl, P. Compensation mechanism in tumor cell migration: Mesenchymal-amoeboid transition after blocking of pericellular proteolysis. J. Cell Biol 2003, 160, 267–277. [Google Scholar]
- Wells, R.G. The role of matrix stiffness in regulating cell behavior. Hepatology 2008, 47, 1394–1400. [Google Scholar]
- Kim, J.; Hayward, R.C. Mimicking dynamic in vivo environments with stimuli-responsive materials for cell culture. Trends Biotechnol 2012, 30, 426–439. [Google Scholar]
- Ingber, D.E.; Mammoto, T. Mechanical control of tissue and organ development. Development 2010, 137, 1407–1420. [Google Scholar]
- Kim, D.H.; Seo, C.H.; Han, K.; Kwon, K.W.; Levchenko, A.; Suh, K.Y. Guided cell migration on microtextured substrates with variable local density and anisotropy. Adv. Funct. Mater 2009, 19, 1579–1586. [Google Scholar]
- Kim, D.Y.; You, M.H.; Kwak, M.K.; Kim, D.H.; Kim, K.; Levchenko, A.; Suh, K.Y. Synergistically enhanced osteogenic differentiation of human mesenchymal stem cells by culture on nanostructured surfaces with induction media. Biomacromolecules 2010, 11, 1856–1862. [Google Scholar]
- Georges, P.C.; Janmey, P.A. Cell type-specific response to growth on soft materials. J. Appl. Physiol 2005, 98, 1547–1553. [Google Scholar]
- Tibbitt, M.W.; Anseth, K.S. Dynamic microenvironments: The fourth dimension. Sci. Transl. Med 2012, 4. [Google Scholar] [CrossRef]
- Burdick, J.A.; Murphy, W.L. Moving from static to dynamic complexity in hydrogel design. Nat. Commun 2012, 3, 1269–1276. [Google Scholar]
- Kloxin, A.M.; Kasko, A.M.; Salinas, C.N.; Anseth, K.S. Photodegradable hydrogels for dynamic tuning of physical and chemical properties. Science 2009, 324, 59–63. [Google Scholar]
- Khetan, S.; Guvendiren, M.; Legant, W.R.; Cohen, D.M.; Chen, C.S.; Burdick, J.A. Degradation-mediated cellular traction directs stem cell fate in covalently crosslinked three-dimensional hydrogels. Nat. Mater 2013, 12, 458–465. [Google Scholar]
- Yamato, M.; Akiyama, Y.; Kobayashi, J.; Yang, J.; Kikuchi, A.; Okano, T. Temperature-responsive cell culture surfaces for regenerative medicine with cell sheet engineering. Prog. Polym. Sci 2007, 32, 1123–1133. [Google Scholar]
- Matsuda, N.; Shimizu, T.; Yamato, M.; Okano, T. Tissue engineering based on cell sheet technology. Adv. Mater 2007, 19, 3089–3099. [Google Scholar]
- Ebara, M.; Yamato, M.; Aoyagi, T.; Kikuchi, A.; Sakai, K.; Okano, T. A novel approach to observing synergy effects of PHSRN on integrin-RGD binding using intelligent surfaces. Adv. Mater 2008, 20, 3034–3038. [Google Scholar]
- Idota, N.; Ebara, M.; Kotsuchibashi, Y.; Narain, R.; Aoyagi, T. Novel temperature-responsive polymer brushes with carbohydrate residues facilitate selective adhesion and collection of hepatocytes. Sci. Technol. Adv. Mat 2012, 13, 064206. [Google Scholar]
- Nakanishi, J.; Kikuchi, Y.; Takarada, T.; Nakayama, H.; Yamaguchi, K.; Maeda, M. Photoactivation of a substrate for cell adhesion under standard fluorescence microscopes. J. Am. Chem. Soc 2004, 126, 16314–16315. [Google Scholar]
- Rolli, C.G.; Nakayama, H.; Yamaguchi, K.; Spatz, J.P.; Kemkemer, R.; Nakanishi, J. Switchable adhesive substrates: Revealing geometry dependence in collective cell behavior. Biomaterials 2012, 33, 2409–2418. [Google Scholar]
- Kloxin, A.M.; Tibbitt, M.W.; Anseth, K.S. Synthesis of photodegradable hydrogels as dynamically tunable cell culture platforms. Nat. Protocols 2010, 5, 1867–1887. [Google Scholar]
- Yoshikawa, H.Y.; Rossetti, F.F.; Kaufmann, S.; Kaindl, T.; Madsen, J.; Engel, U.; Lewis, A.L.; Armes, S.P.; Tanaka, M. Quantitative evaluation of mechanosensing of cells on dynamically tunable hydrogels. J. Am. Chem. Soc 2011, 133, 1367–1374. [Google Scholar]
- Ebara, M.; Uto, K.; Idota, N.; Hoffman, J.M.; Aoyagi, T. Shape-memory surface with dynamically tunable nano-geometry activated by body heat. Adv. Mater 2012, 24, 273–278. [Google Scholar]
- Ebara, M.; Uto, K.; Idota, N.; Hoffman, J.M.; Aoyagi, T. The Taming of the Cell; Shape-memory nanopatterns direct cell orientation. Int. J. Nanomed 2013, in press. [Google Scholar]
- Lendlein, A.; Kelch, S. Shape-memory polymers. Angew. Chem. Int. Ed 2002, 41, 2035–2057. [Google Scholar]
- Lendlein, A.; Langer, R. Biodegradable, elastic shape-memory polymers for potential biomedical applications. Science 2002, 296, 1673–1676. [Google Scholar]
- Forte, G.; Pagliari, S.; Ebara, M.; Uto, K.; Tam, J.K.; Romanazzo, S.; Escobedo-Lucea, C.; Romano, E.; Di Nardo, P.; Traversa, E.; et al. Substrate stiffness modulates gene expression and phenotype in neonatal cardiomyocytes in vitro. Tissue Eng. Part A 2012, 18, 1837–1848. [Google Scholar]
- Tam, J.K.V.; Uto, K.; Ebara, M.; Pagliari, S.; Forte, G.; Aoyagi, T. Mesenchymal stem cell adhesion but not plasticity is affected by high substrate stiffness. Sci. Technol. Adv. Mat 2012, 13, 064205. [Google Scholar]
- Romanazzo, S.; Forte, G.; Ebara, M.; Uto, K.; Pagliari, S.; Aoyagi, T.; Traversa, E.; Taniguchi, A. Substrate stiffness affects skeletal myoblast differentiation in vitro. Sci. Technol. Adv. Mat. 2012, 13, 064211. [Google Scholar]
- Webb, K.; Hlady, V.; Tresco, P.A. Relative importance of surface wettability and charged functional groups on NIH3T3 fibroblast attachment, spreading, and cytoskeletal organization. J. Biomed. Mater. Res 1998, 41, 422–430. [Google Scholar]
- Mrksich, M. A surface chemistry approach to studying cell adhesion. Chem. Soc. Rev 2000, 29, 267–273. [Google Scholar]
- Wurm, A.; Zhuravlev, E.; Eckstein, K.; Jehnichen, D.; Pospiech, D.; Androsch, R.; Wunderlich, B.; Schick, C. Crystallization and homogeneous nucleation kinetics of Poly(ɛ-caprolactone) (PCL) with different molar masses. Macromolecules 2012, 45, 3816–3828. [Google Scholar]
- Martínez, F.N.; García, R. Measuring phase shifts and energy dissipation with amplitude modulation atomic force microscopy. Nanotechnology 2006, 17, S167. [Google Scholar]
- Modulevsky, D.J.; Tremblay, D.; Gullekson, C.; Bukoreshtliev, N.V.; Pelling, A.E. The physical interaction of myoblasts with the microenvironment during remodeling of the cytoarchitecture. PLoS One 2012, 7, e45329. [Google Scholar]
- Yamato, M.; Okuhara, M.; Karikusa, F.; Kikuchi, A.; Sakurai, Y.; Okano, T. Signal transduction and cytoskeletal reorganization are required for cell detachment from cell culture surfaces grafted with a temperature-responsive polymer. J. Biomed. Mater. Res 1999, 44, 44–52. [Google Scholar]
- Kushida, A.; Yamato, M.; Konno, C.; Kikuchi, A.; Sakurai, Y.; Okano, T. Temperature-responsive culture dishes allow nonenzymatic harvest of differentiated Madin-Darby canine kidney (MDCK) cell sheets. J. Biomed. Mater. Res 2000, 51, 216–223. [Google Scholar]
- Yoshikawa, H.Y.; Kawano, T.; Matsuda, T.; Kidoaki, S.; Tanaka, M. Morphology and adhesion strength of myoblast cells on photocurable gelatin under native and non-native micromechanical environments. J. Phys. Chem. B 2013, 117, 4081–4088. [Google Scholar]
- Ren, K.; Crouzier, T.; Roy, C.; Picart, C. Polyelectrolyte multilayer films of controlled stiffness modulate myoblast cell differentiation. Adv. Funct. Mater 2008, 18, 1378–1389. [Google Scholar]
- Al-Rekabi, Z.; Pelling, A.E. Cross talk between matrix elasticity and mechanical force regulates myoblast traction dynamics. Phys. Biol 2013, 10, 066003. [Google Scholar]
- Uto, K.; Yamamoto, K.; Hirase, S.; Aoyagi, T. Temperature-responsive cross-linked poly(epsilon-caprolactone) membrane that functions near body temperature. J. Control. Release 2006, 110, 408–413. [Google Scholar]
- Ebara, M.; Uto, K.; Idota, N.; Hoffman, J.M.; Aoyagi, T. Rewritable and shape-memory soft matter with dynamically tunable microchannel geometry in a biological temperature range. Soft Matter 2013, 9, 3074–3080. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2b/4bPCL | Tm (°C) | Elasticity (MPa) | Roughness (nm) | Contact angle (°) | |||
|---|---|---|---|---|---|---|---|
| 25 °C | 45 °C | 25 °C | 45 °C | 25 °C | 45 °C | ||
| 0/100 | - | 1.4 (±0.4) | 1.5 (±0.1) | 6.2 | 6.1 | 96.5 (±1.6) | 97.4 (±2.4) |
| 50/50 | 33.0 (±0.1) | 52.9 (±3.0) | 1.1 (±0.1) | 63.4 | 12.4 | 96.3 (±3.7) | 100.9 (±3.3) |
| 70/30 | 37.8 (±0.1) | 97.6 (±3.6) | 0.9 (±0.1) | 83.6 | 13.4 | 95.8 (±2.1) | 96.6 (±2.8) |
| 100/0 | 43.7 (±0.3) | 153.4 (±8.0) | 9.3 (±2.7) | 85.5 | 27.6 | 100.5 (±3.1) | 102.5 (±1.5) |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Uto, K.; Ebara, M.; Aoyagi, T. Temperature-Responsive Poly(ε-caprolactone) Cell Culture Platform with Dynamically Tunable Nano-Roughness and Elasticity for Control of Myoblast Morphology. Int. J. Mol. Sci. 2014, 15, 1511-1524. https://doi.org/10.3390/ijms15011511
Uto K, Ebara M, Aoyagi T. Temperature-Responsive Poly(ε-caprolactone) Cell Culture Platform with Dynamically Tunable Nano-Roughness and Elasticity for Control of Myoblast Morphology. International Journal of Molecular Sciences. 2014; 15(1):1511-1524. https://doi.org/10.3390/ijms15011511
Chicago/Turabian StyleUto, Koichiro, Mitsuhiro Ebara, and Takao Aoyagi. 2014. "Temperature-Responsive Poly(ε-caprolactone) Cell Culture Platform with Dynamically Tunable Nano-Roughness and Elasticity for Control of Myoblast Morphology" International Journal of Molecular Sciences 15, no. 1: 1511-1524. https://doi.org/10.3390/ijms15011511