The Roles of Monomeric GTP-Binding Proteins in Macroautophagy in Saccharomyces cerevisiae

Abstract
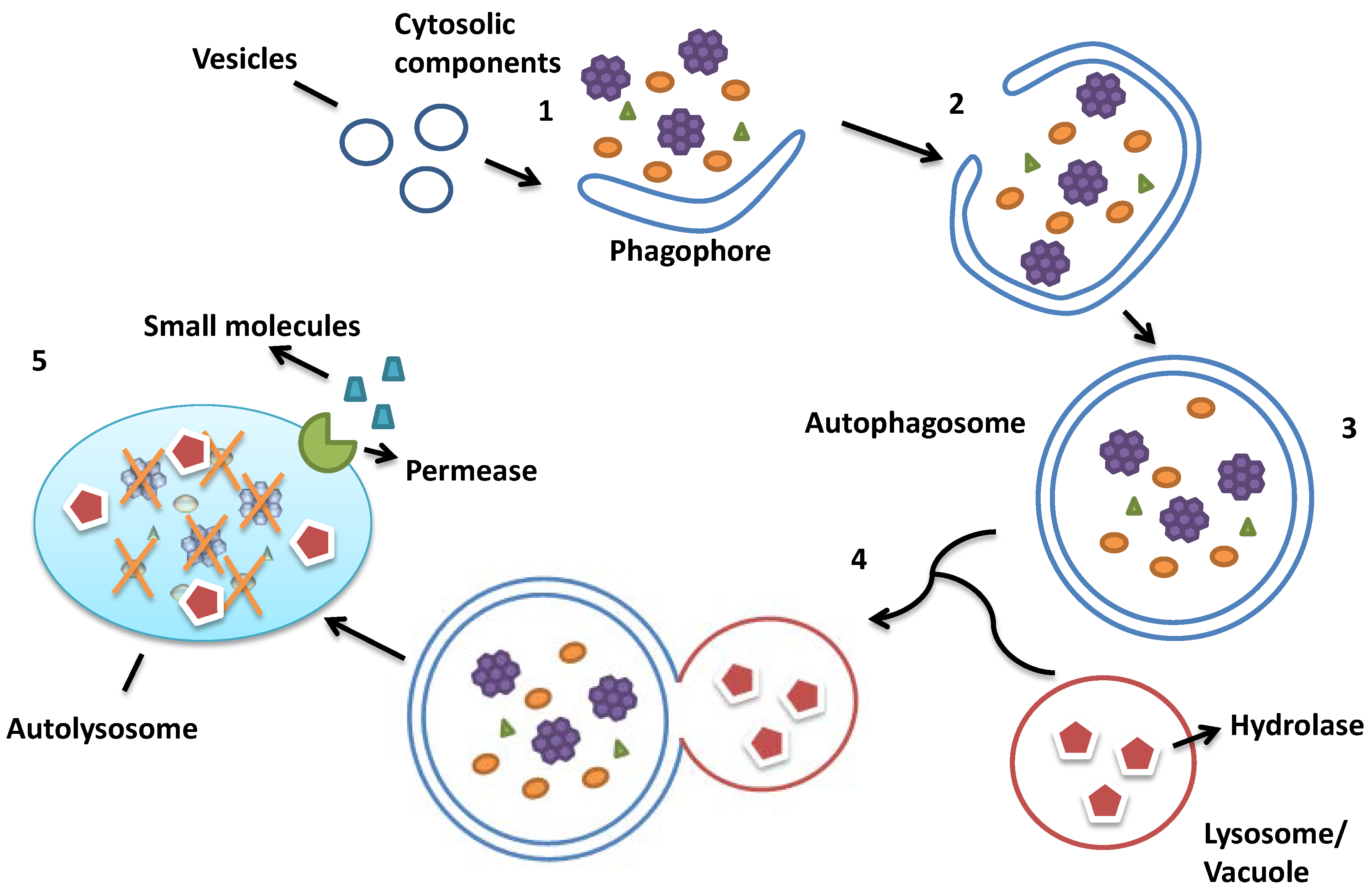
:1. Autophagy in Saccharomyces cerevisiae



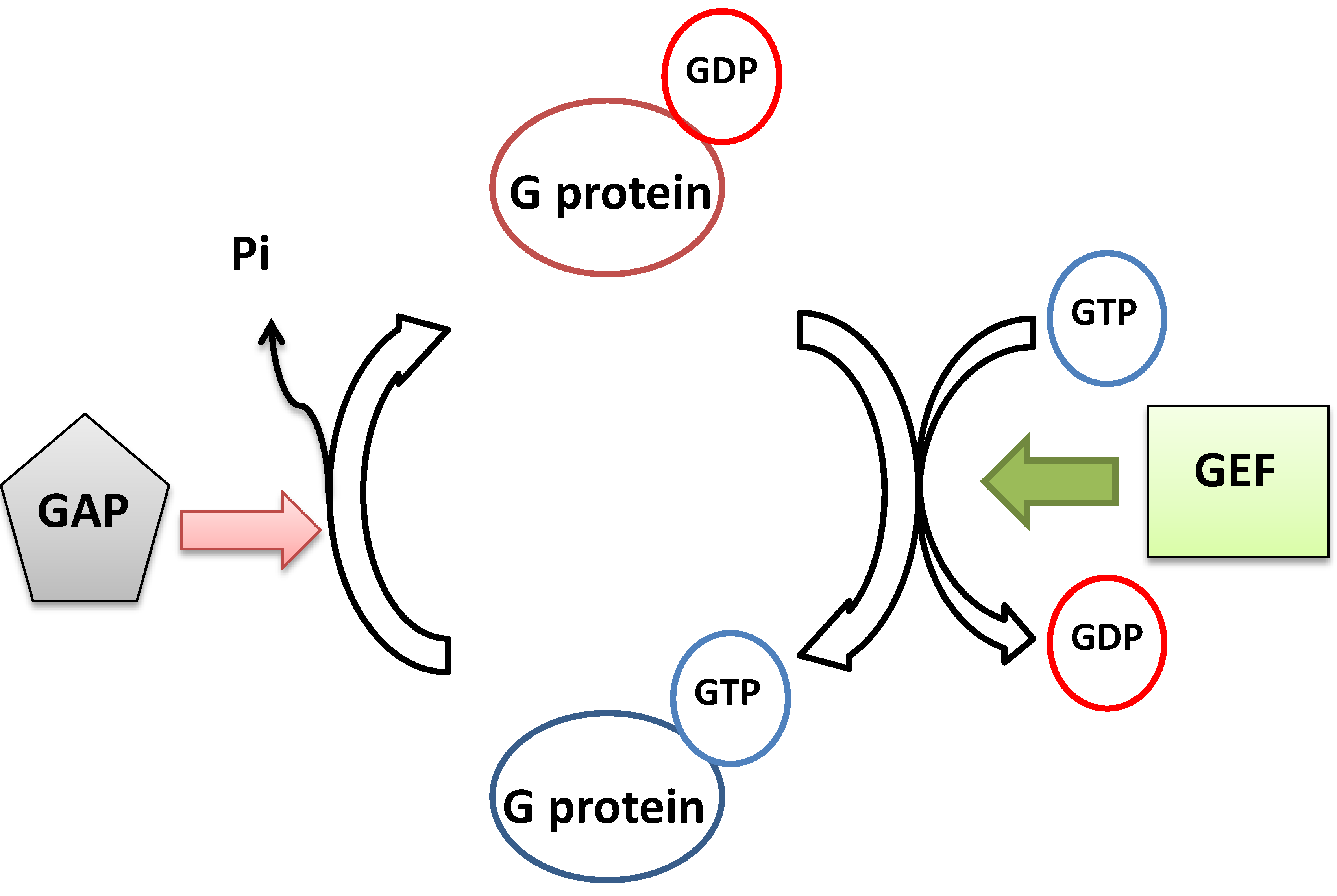
2. The Monomeric GTP-Binding Proteins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. cerevisiae | Human | Subcellular Localization | Functions in Membrane Trafficking |
|---|---|---|---|
| Ypt1 | Rab1 | ER-Golgi, intra-Golgi | Tethers and docks COP II vesicles to cis-Golgi membrane |
| Sar1 | Sar1 | ER-Golgi compartment | Assists COP II vesicles budding from ER |
| Arf1 | Arf1 | Golgi, Golgi to endosome | Regulates the budding of COP I and clathrin vesicles |
| Arl1 | Arl1 | trans-Golgi network (TGN) | Controls retrograde trafficking from endosome to TGN |
| Ypt6 | Rab6 | Golgi-endosome | Mediates fusion between endosome-derived vesicle to Golgi membrane |
| Vps21/Ypt52/Ypt53 | Rab5 | Early endosome | Regulates endocytic trafficking |
| Ypt7 | Rab7 | Late endosome | Regulates fusion between endosomes |
| Ypt31/Ypt32 | Rab11 | Post Golgi | Controls exocytic pathway |
| Sec4 | Sec4 | Post Golgi | Delivers exocytic vesicles to plasma membrane |

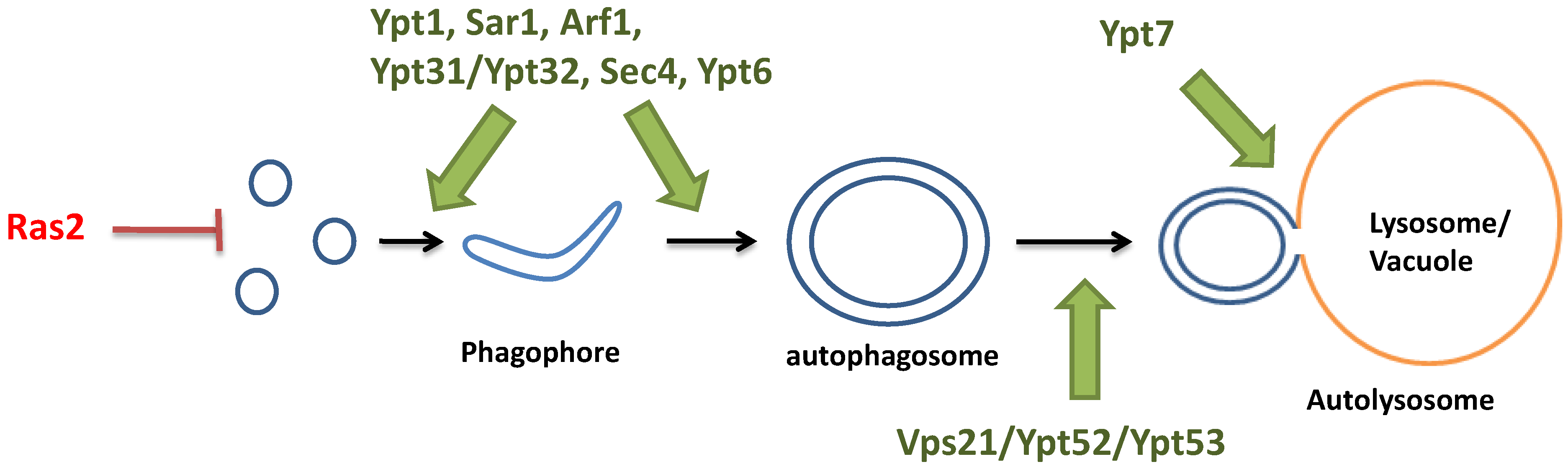
3. Monomeric GTP-Binding Proteins in Autophagy
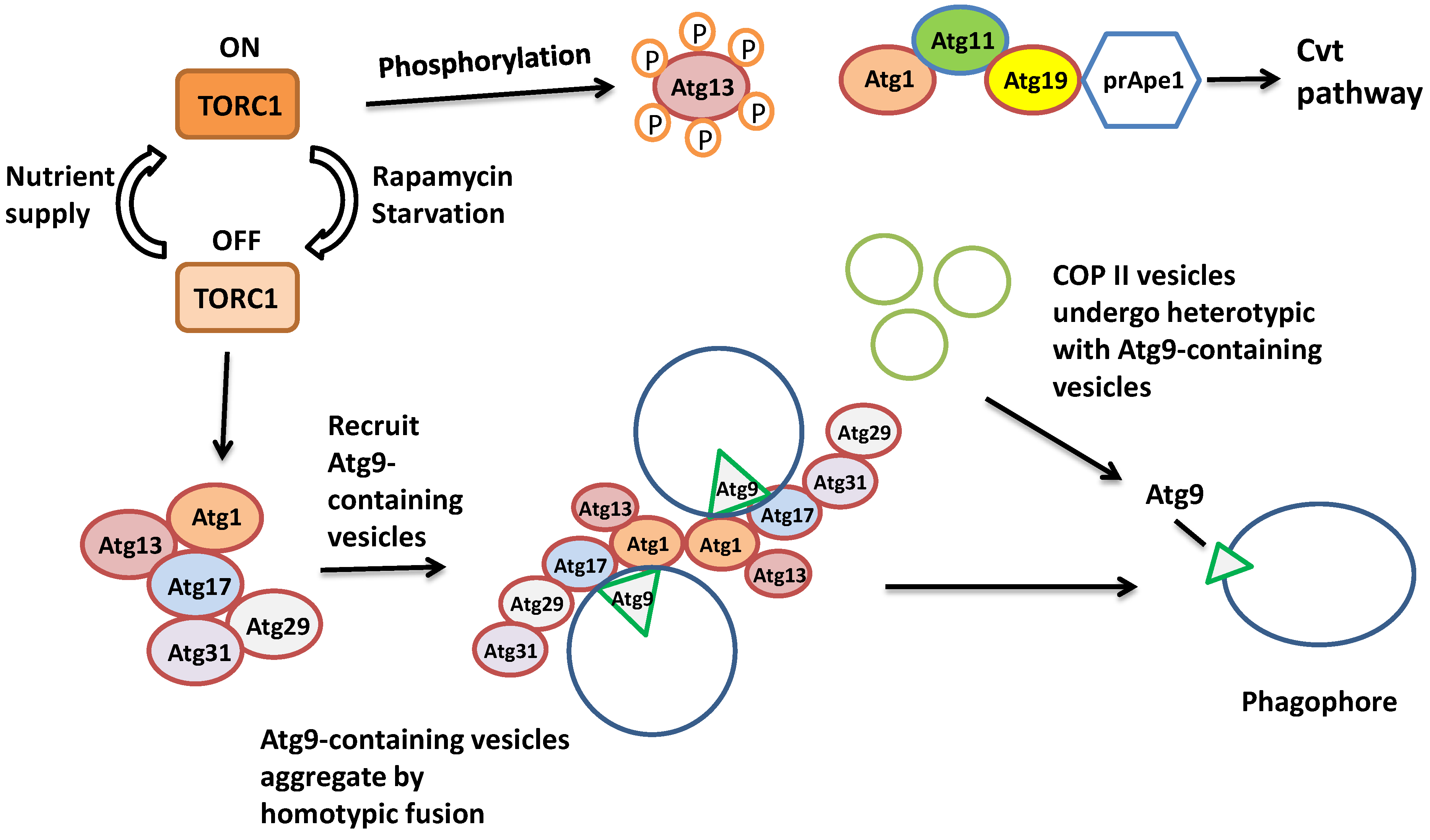
3.1. Ras Proteins in the Early Initiation of Autophagy
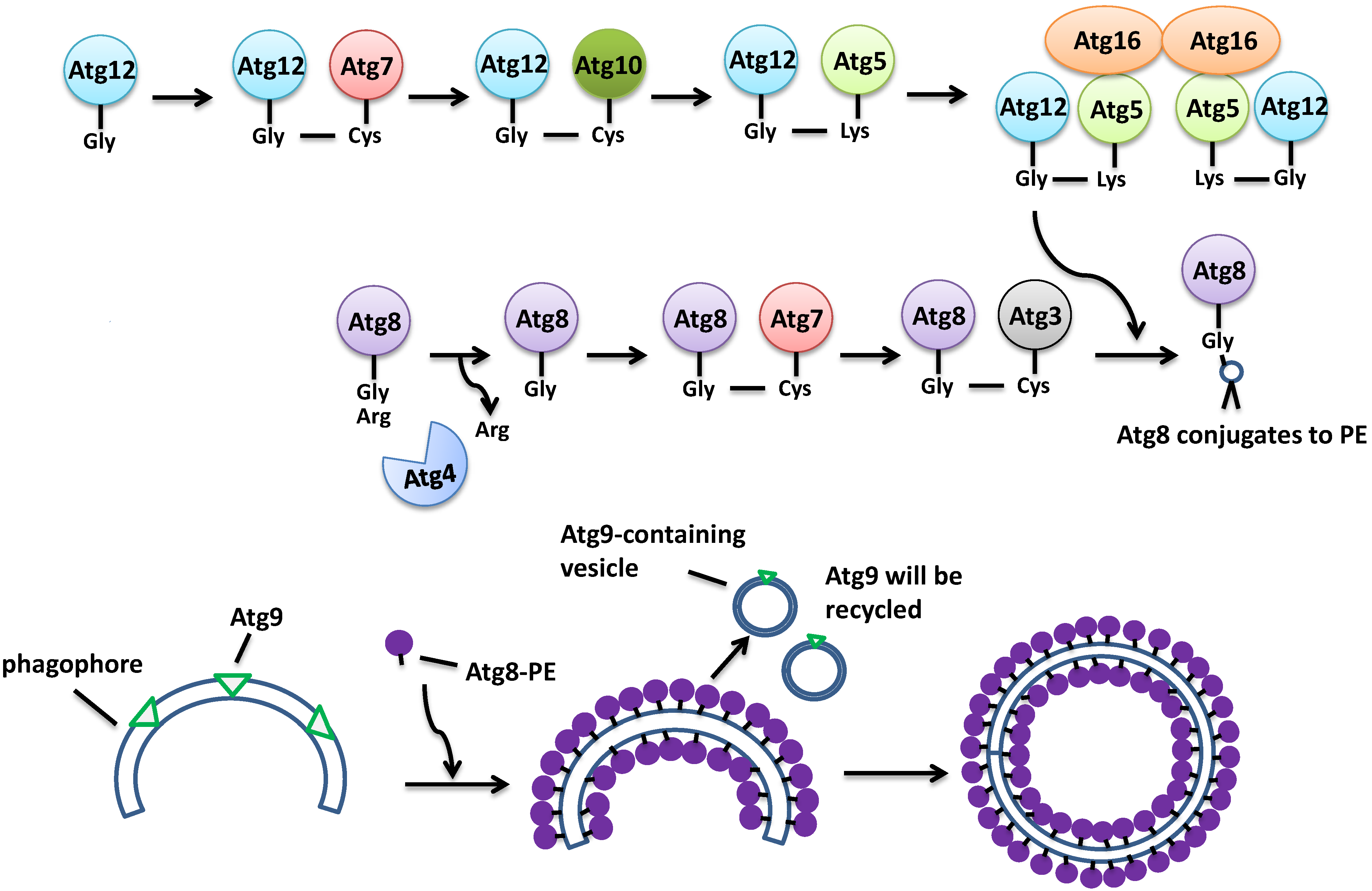
3.2. Monomeric GTP-Binding Proteins in the Formation of the Autophagosome

3.3. Monomeric GTP-Binding Proteins in the Fusion between the Autophagosome and the Vacuole
4. Summary and Conclusions; Future Directions

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Glick, D.; Barth, S.; Macleod, K.F. Autophagy: Cellular and molecular mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Klionsky, D.J. Eaten alive: A history of macroautophagy. Nat. Cell Biol. 2010, 12, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Orvedahl, A.; Levine, B. Eating the enemy within: autophagy in infectious diseases. Cell Death Differ. 2009, 16, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Klionsky, D.J. Autophagosome formation: core machinery and adaptations. Nat. Cell Biol. 2007, 9, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Ragusa, M.J.; Stanley, R.E.; Hurley, J.H. Architecture of the Atg17 complex as a scaffold for autophagosome biogenesis. Cell 2012, 151, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Cuervo, A.M.; Seglen, P.O. Methods for monitoring autophagy from yeast to human. Autophagy 2007, 3, 181–206. [Google Scholar] [CrossRef] [PubMed]
- Schimmoller, F.; Riezman, H. Involvement of Ypt7p, a small GTPase, in traffic from late endosome to the vacuole in yeast. J. Cell Sci. 1993, 106, 823–830. [Google Scholar] [PubMed]
- Eskelinen, E.L. Maturation of autophagic vacuoles in Mammalian cells. Autophagy 2005, 1, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Stephan, J.S.; Yeh, Y.Y.; Ramachandran, V.; Deminoff, S.J.; Herman, P.K. The Tor and PKA signaling pathways independently target the Atg1/Atg13 protein kinase complex to control autophagy. Proc. Natl. Acad. Sci. USA 2009, 106, 17049–17054. [Google Scholar] [CrossRef] [PubMed]
- Stephan, J.S.; Yeh, Y.Y.; Ramachandran, V.; Deminoff, S.J.; Herman, P.K. The Tor and cAMP-dependent protein kinase signaling pathways coordinately control autophagy in Saccharomyces cerevisiae. Autophagy 2010, 6, 294–295. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Cueva, R.; Yaver, D.S. Aminopeptidase I of Saccharomyces cerevisiae is localized to the vacuole independent of the secretory pathway. J. Cell Biol. 1992, 119, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Hutchins, M.U.; Klionsky, D.J. Vacuolar localization of oligomeric alpha-mannosidase requires the cytoplasm to vacuole targeting and autophagy pathway components in Saccharomyces cerevisiae. J. Biol. Chem. 2001, 276, 20491–20498. [Google Scholar] [CrossRef] [PubMed]
- Yuga, M.; Gomi, K.; Klionsky, D.J.; Shintani, T. Aspartyl aminopeptidase is imported from the cytoplasm to the vacuole by selective autophagy in Saccharomyces cerevisiae. J. Biol. Chem. 2011, 286, 13704–13713. [Google Scholar] [CrossRef] [PubMed]
- Lynch-Day, M.A.; Klionsky, D.J. The Cvt pathway as a model for selective autophagy. FEBS Lett. 2010, 584, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.; Rodriguez-Enriquez, S.; Lemasters, J.J. Selective degradation of mitochondria by mitophagy. Arch. Biochem. Biophys. 2007, 462, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W.A., Jr.; Cregg, J.M.; Kiel, J.A.; van der Klei, I.J.; Oku, M.; Sakai, Y.; Sibirny, A.A.; Stasyk, O.V.; Veenhuis, M. Pexophagy: The selective autophagy of peroxisomes. Autophagy 2005, 1, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Bernales, S.; McDonald, K.L.; Walter, P. Autophagy counterbalances endoplasmic reticulum expansion during the unfolded protein response. PLoS Biol. 2006, 4, e423. [Google Scholar] [CrossRef]
- Kraft, C.; Deplazes, A.; Sohrmann, M.; Peter, M. Mature ribosomes are selectively degraded upon starvation by an autophagy pathway requiring the Ubp3p/Bre5p ubiquitin protease. Nat. Cell Biol. 2008, 10, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Ohsumi, Y. Molecular machinery of autophagosome formation in yeast, Saccharomyces cerevisiae. FEBS Lett. 2007, 581, 2156–2161. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Codogno, P.; Cuervo, A.M.; Deretic, V.; Elazar, Z.; Fueyo-Margareto, J.; Gewirtz, D.A.; Kroemer, G.; Levine, B.; Mizushima, N.; et al. A comprehensive glossary of autophagy-related molecules and processes. Autophagy 2010, 6, 438–448. [Google Scholar]
- Nair, U.; Jotwani, A.; Geng, J.; Gammoh, N.; Richerson, D.; Yen, W.L.; Griffith, J.; Nag, S.; Wang, K.; Moss, T.; et al. SNARE proteins are required for macroautophagy. Cell 2011, 146, 290–302. [Google Scholar]
- Kakuta, S.; Yamamoto, H.; Negishi, L.; Kondo-Kakuta, C.; Hayashi, N.; Ohsumi, Y. Atg9 vesicles recruit vesicle-tethering proteins Trs85 and Ypt1 to the autophagosome formation site. J. Biol. Chem. 2012, 287, 44261–44269. [Google Scholar] [CrossRef] [PubMed]
- Lynch-Day, M.A.; Bhandari, D.; Menon, S.; Huang, J.; Cai, H.; Bartholomew, C.R.; Brumell, J.H.; Ferro-Novick, S.; Klionsky, D.J. Trs85 directs a Ypt1 GEF, TRAPPIII, to the phagophore to promote autophagy. Proc. Natl. Acad. Sci. USA 2010, 107, 7811–7816. [Google Scholar] [CrossRef] [PubMed]
- Meiling-Wesse, K.; Epple, U.D.; Krick, R.; Barth, H.; Appelles, A.; Voss, C.; Eskelinen, E.L.; Thumm, M. Trs85 (Gsg1), a component of the TRAPP complexes, is required for the organization of the preautophagosomal structure during selective autophagy via the Cvt pathway. J. Biol. Chem. 2005, 280, 33669–33678. [Google Scholar] [CrossRef] [PubMed]
- Wennerberg, K.; Rossman, K.L.; Der, C.J. The Ras superfamily at a glance. J. Cell Sci. 2005, 118, 843–846. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.S.; Blobel, G. A G protein involved in nucleocytoplasmic transport: The role of Ran. Trends Biochem. Sci. 1994, 19, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Boureux, A.; Vignal, E.; Faure, S.; Fort, P. Evolution of the Rho family of ras-like GTPases in eukaryotes. Mol. Biol. Evol. 2007, 24, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Takai, Y.; Sasaki, T.; Matozaki, T. Small GTP-binding proteins. Physiol. Rev. 2001, 81, 153–208. [Google Scholar]
- Kahn, R.A.; Volpicelli-Daley, L.; Bowzard, B.; Shrivastava-Ranjan, P.; Li, Y.; Zhou, C.; Cunningham, L. Arf family GTPases: Roles in membrane traffic and microtubule dynamics. Biochem. Soc. Trans. 2005, 33, 1269–1272. [Google Scholar] [CrossRef] [PubMed]
- Bialek-Wyrzykowska, U.; Bauer, B.E.; Wagner, W.; Kohlwein, S.D.; Schweyen, R.J.; Ragnini, A. Low levels of Ypt protein prenylation cause vesicle polarization defects and thermosensitive growth that can be suppressed by genes involved in cell wall maintenance. Mol. Microbiol. 2000, 35, 1295–1311. [Google Scholar] [CrossRef] [PubMed]
- Rak, A.; Pylypenko, O.; Durek, T.; Watzke, A.; Kushnir, S.; Brunsveld, L.; Waldmann, H.; Goody, R.S.; Alexandrov, K. Structure of Rab GDP-dissociation inhibitor in complex with prenylated YPT1 GTPase. Science 2003, 302, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, S.; Aivazian, D. Targeting Rab GTPases to distinct membrane compartments. Nat. Rev. Mol. Cell Biol. 2004, 5, 886–896. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J. Structural basis for activation of ARF GTPase: Mechanisms of guanine nucleotide exchange and GTP-myristoyl switching. Cell 1998, 95, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Amor, J.C.; Horton, J.R.; Zhu, X.; Wang, Y.; Sullards, C.; Ringe, D.; Cheng, X.; Kahn, R.A. Structures of yeast ARF2 and ARL1: Distinct roles for the N terminus in the structure and function of ARF family GTPases. J. Biol. Chem. 2001, 276, 42477–42484. [Google Scholar] [CrossRef] [PubMed]
- Pasqualato, S.; Renault, L.; Cherfils, J. Arf, Arl, Arp and Sar proteins: A family of GTP-binding proteins with a structural device for ‘front-back’ communication. EMBO Rep. 2002, 3, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Fell, G.L.; Munson, A.M.; Croston, M.A.; Rosenwald, A.G. Identification of yeast genes involved in K+ homeostasis: Loss of membrane traffic genes affects K+ uptake. G3 2011, 1, 43–56. [Google Scholar] [CrossRef]
- Manlandro, C.M.; Palanivel, V.R.; Schorr, E.B.; Mihatov, N.; Antony, A.A.; Rosenwald, A.G. Mon2 is a negative regulator of the monomeric G protein, Arl1. FEMS Yeast Res. 2012, 12, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, T.; Powers, S.; McGill, C.; Fasano, O.; Strathern, J.; Broach, J.; Wigler, M. Genetic analysis of yeast RAS1 and RAS2 genes. Cell 1984, 37, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Chen, L.; Broach, J.R.; Powers, S. Ras membrane targeting is essential for glucose signaling but not for viability in yeast. Proc. Natl. Acad. Sci. USA 1995, 92, 2984–2988. [Google Scholar] [CrossRef] [PubMed]
- Broek, D.; Samiy, N.; Fasano, O.; Fujiyama, A.; Tamanoi, F.; Northup, J.; Wigler, M. Differential activation of yeast adenylate cyclase by wild-type and mutant RAS proteins. Cell 1985, 41, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Gimeno, C.J.; Ljungdahl, P.O.; Styles, C.A.; Fink, G.R. Unipolar cell divisions in the yeast S. cerevisiae lead to filamentous growth: regulation by starvation and RAS. Cell 1992, 68, 1077–1090. [Google Scholar]
- Budovskaya, Y.V.; Stephan, J.S.; Reggiori, F.; Klionsky, D.J.; Herman, P.K. The Ras/cAMP-dependent protein kinase signaling pathway regulates an early step of the autophagy process in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 20663–20671. [Google Scholar] [CrossRef] [PubMed]
- Toda, T.; Cameron, S.; Sass, P.; Wigler, M. SCH9, a gene of Saccharomyces cerevisiae that encodes a protein distinct from, but functionally and structurally related to, cAMP-dependent protein kinase catalytic subunits. Genes Dev. 1988, 2, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Morano, K.A.; Thiele, D.J. The Sch9 protein kinase regulates Hsp90 chaperone complex signal transduction activity in vivo. EMBO J. 1999, 18, 5953–5962. [Google Scholar] [CrossRef]
- Fabrizio, P.; Pozza, F.; Pletcher, S.D.; Gendron, C.M.; Longo, V.D. Regulation of longevity and stress resistance by Sch9 in yeast. Science 2001, 292, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Toda, T.; Cameron, S.; Sass, P.; Zoller, M.; Wigler, M. Three different genes in S. cerevisiae encode the catalytic subunits of the cAMP-dependent protein kinase. Cell 1987, 50, 277–287. [Google Scholar]
- Yorimitsu, T.; Zaman, S.; Broach, J.R.; Klionsky, D.J. Protein kinase A and Sch9 cooperatively regulate induction of autophagy in Saccharomyces cerevisiae. Mol. Biol. Cell 2007, 18, 4180–4189. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Kakuta, S.; Watanabe, T.M.; Kitamura, A.; Sekito, T.; Kondo-Kakuta, C.; Ichikawa, R.; Kinjo, M.; Ohsumi, Y. Atg9 vesicles are an important membrane source during early steps of autophagosome formation. J. Cell Biol. 2012, 198, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Militello, R.D.; Colombo, M.I. A membrane is born: Origin of the autophagosomal compartment. Curr. Mol. Med. 2011, 11, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Barrowman, J.; Bhandari, D.; Reinisch, K.; Ferro-Novick, S. TRAPP complexes in membrane traffic: Convergence through a common Rab. Nat. Rev. Mol. Cell Biol. 2010, 11, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Morsomme, P.; Riezman, H. The Rab GTPase Ypt1p and tethering factors couple protein sorting at the ER to vesicle targeting to the Golgi apparatus. Dev. Cell 2002, 2, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Heitman, J.; Movva, N.R.; Hall, M.N. Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science 1991, 253, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Noda, T.; Ohsumi, Y. Tor, a phosphatidylinositol kinase homologue, controls autophagy in yeast. J. Biol. Chem. 1998, 273, 3963–3966. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Menon, S.; Yamasaki, A.; Chou, H.T.; Walz, T.; Jiang, Y.; Ferro-Novick, S. Ypt1 recruits the Atg1 kinase to the preautophagosomal structure. Proc. Natl. Acad. Sci. USA 2013, 110, 9800–9805. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Warner, J.R. Mutation of the Rab6 homologue of Saccharomyces cerevisiae, YPT6, inhibits both early Golgi function and ribosome biosynthesis. J. Biol. Chem. 1996, 271, 16813–16819. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Zou, S.; Yu, S.; Liang, Y. Ypt1 suppresses defects of vesicle trafficking and autophagy in Ypt6 related mutants. Cell Biol. Int. 2014, 38, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.G.; Raunser, S.; Munger, C.; Wagner, J.; Song, Y.L.; Cygler, M.; Walz, T.; Oh, B.H.; Sacher, M. The architecture of the multisubunit TRAPP I complex suggests a model for vesicle tethering. Cell 2006, 127, 817–830. [Google Scholar] [CrossRef] [PubMed]
- Morozova, N.; Liang, Y.; Tokarev, A.A.; Chen, S.H.; Cox, R.; Andrejic, J.; Lipatova, Z.; Sciorra, V.A.; Emr, S.D.; Segev, N. TRAPPII subunits are required for the specificity switch of a Ypt-Rab GEF. Nat. Cell Biol. 2006, 8, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Reinisch, K.; Ferro-Novick, S. Coats, Tethers, Rabs, and SNAREs work together to mediate the intracellular destination of a transport vesicle. Dev. Cell 2007, 12, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, N.; Hamasaki, M.; Yokota, S.; Suzuki, K.; Kamada, Y.; Kihara, A.; Yoshimori, T.; Noda, T.; Ohsumi, Y. Autophagosome requires specific early Sec proteins for its formation and NSF/SNARE for vacuolar fusion. Mol. Biol. Cell 2001, 12, 3690–3702. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.; Cai, Y.; Wang, J.; Zhang, J.; Menon, S.; Chou, H.T.; Ferro-Novick, S.; Reinisch, K.M.; Walz, T. The EM structure of the TRAPPIII complex leads to the identification of a requirement for COPII vesicles on the macroautophagy pathway. Proc. Natl. Acad. Sci. USA 2013, 110, 19432–19437. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Novick, P.J. A ras-like protein is required for a post-Golgi event in yeast secretion. Cell 1987, 49, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Roth, D.; Walch-Solimena, C.; Novick, P. The exocyst is an effector for Sec4p, targeting secretory vesicles to sites of exocytosis. EMBO J. 1999, 18, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Mari, M.; Griffith, J.; Rieter, E.; Krishnappa, L.; Klionsky, D.J.; Reggiori, F. An Atg9-containing compartment that functions in the early steps of autophagosome biogenesis. J. Cell Biol. 2010, 190, 1005–1022. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Nair, U.; Yasumura-Yorimitsu, K.; Klionsky, D.J. Post-Golgi Sec proteins are required for autophagy in Saccharomyces cerevisiae. Mol. Biol. Cell 2010, 21, 2257–2269. [Google Scholar] [CrossRef] [PubMed]
- Benli, M.; Doring, F.; Robinson, D.G.; Yang, X.; Gallwitz, D. Two GTPase isoforms, Ypt31p and Ypt32p, are essential for Golgi function in yeast. EMBO J. 1996, 15, 6460–6475. [Google Scholar] [PubMed]
- Ortiz, D.; Medkova, M.; Walch-Solimena, C.; Novick, P. Ypt32 recruits the Sec4p guanine nucleotide exchange factor, Sec2p, to secretory vesicles; evidence for a Rab cascade in yeast. J. Cell Biol. 2002, 157, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Shintani, T.; Reggiori, F. Fluorescence microscopy-based assays for monitoring yeast Atg protein trafficking. Methods Enzymol. 2008, 451, 43–56. [Google Scholar] [PubMed]
- Singer-Kruger, B.; Stenmark, H.; Dusterhoft, A.; Philippsen, P.; Yoo, J.S.; Gallwitz, D.; Zerial, M. Role of three rab5-like GTPases, Ypt51p, Ypt52p, and Ypt53p, in the endocytic and vacuolar protein sorting pathways of yeast. J. Cell Biol. 1994, 125, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, M.; Arlt, H.; Epp, N.; Lachmann, J.; Griffith, J.; Perz, A.; Reggiori, F.; Ungermann, C. Functional separation of endosomal fusion factors and the class C core vacuole/endosome tethering (CORVET) complex in endosome biogenesis. J. Biol. Chem. 2013, 288, 5166–5175. [Google Scholar] [CrossRef] [PubMed]
- Nickerson, D.P.; Russell, M.R.; Lo, S.Y.; Chapin, H.C.; Milnes, J.M.; Merz, A.J. Termination of isoform-selective Vps21/Rab5 signaling at endolysosomal organelles by Msb3/Gyp3. Traffic 2012, 13, 1411–1428. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, F.; Zou, S.; Yu, S.; Li, S.; Li, D.; Song, J.; Li, H.; He, Z.; Hu, B.; et al. A Vps21 endocytic module regulates autophagy. Mol. Biol. Cell 2014. [Google Scholar] [CrossRef]
- Van der Vaart, A.; Griffith, J.; Reggiori, F. Exit from the Golgi is required for the expansion of the autophagosomal phagophore in yeast Saccharomyces cerevisiae. Mol. Biol. Cell 2010, 21, 2270–2284. [Google Scholar] [CrossRef] [PubMed]
- Haas, A.; Scheglmann, D.; Lazar, T.; Gallwitz, D.; Wickner, W. The GTPase Ypt7p of Saccharomyces cerevisiae is required on both partner vacuoles for the homotypic fusion step of vacuole inheritance. EMBO J. 1995, 14, 5258–5270. [Google Scholar] [PubMed]
- Nordmann, M.; Cabrera, M.; Perz, A.; Brocker, C.; Ostrowicz, C.; Engelbrecht-Vandre, S.; Ungermann, C. The Mon1-Ccz1 complex is the GEF of the late endosomal Rab7 homolog Ypt7. Curr. Biol. 2010, 20, 1654–1659. [Google Scholar] [CrossRef] [PubMed]
- Brocker, C.; Kuhlee, A.; Gatsogiannis, C.; Balderhaar, H.J.; Honscher, C.; Engelbrecht-Vandre, S.; Ungermann, C.; Raunser, S. Molecular architecture of the multisubunit homotypic fusion and vacuole protein sorting (HOPS) tethering complex. Proc. Natl. Acad. Sci. USA 2012, 109, 1991–1996. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.K.; Darsow, T.; Emr, S.D. Vam7p, a SNAP-25-like molecule, and Vam3p, a syntaxin homolog, function together in yeast vacuolar protein trafficking. Mol. Cell Biol. 1998, 18, 5308–5319. [Google Scholar] [PubMed]
- Ungermann, C.; Price, A.; Wickner, W. A new role for a SNARE protein as a regulator of the Ypt7/Rab-dependent stage of docking. Proc. Natl. Acad. Sci. USA 2000, 97, 8889–8891. [Google Scholar] [CrossRef] [PubMed]
- Kirisako, T.; Baba, M.; Ishihara, N.; Miyazawa, K.; Ohsumi, M.; Yoshimori, T.; Noda, T.; Ohsumi, Y. Formation process of autophagosome is traced with Apg8/Aut7p in yeast. J. Cell Biol. 1999, 147, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Darsow, T.; Rieder, S.E.; Emr, S.D. A multispecificity syntaxin homologue, Vam3p, essential for autophagic and biosynthetic protein transport to the vacuole. J. Cell Biol. 1997, 138, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Rosenwald, A.G.; Rhodes, M.A.; van Valkenburgh, H.; Palanivel, V.; Chapman, G.; Boman, A.; Zhang, C.J.; Kahn, R.A. ARL1 and membrane traffic in Saccharomyces cerevisiae. Yeast 2002, 19, 1039–1056. [Google Scholar] [CrossRef] [PubMed]
- Abudugupur, A.; Mitsui, K.; Yokota, S.; Tsurugi, K. An ARL1 mutation affected autophagic cell death in yeast, causing a defect in central vacuole formation. Cell Death Differ. 2002, 9, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Reggiori, F.; Monastyrska, I.; Shintani, T.; Klionsky, D.J. The actin cytoskeleton is required for selective types of autophagy, but not nonspecific autophagy, in the yeast Saccharomyces cerevisiae. Mol. Biol. Cell 2005, 16, 5843–5856. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Rosenwald, A.G. The Roles of Monomeric GTP-Binding Proteins in Macroautophagy in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2014, 15, 18084-18101. https://doi.org/10.3390/ijms151018084
Yang S, Rosenwald AG. The Roles of Monomeric GTP-Binding Proteins in Macroautophagy in Saccharomyces cerevisiae. International Journal of Molecular Sciences. 2014; 15(10):18084-18101. https://doi.org/10.3390/ijms151018084
Chicago/Turabian StyleYang, Shu, and Anne G. Rosenwald. 2014. "The Roles of Monomeric GTP-Binding Proteins in Macroautophagy in Saccharomyces cerevisiae" International Journal of Molecular Sciences 15, no. 10: 18084-18101. https://doi.org/10.3390/ijms151018084