2.1. Inhibition of in Vitro ACE and Human Recombinant Renin Activities by Flaxseed Protein Hydrolysates (FPH)
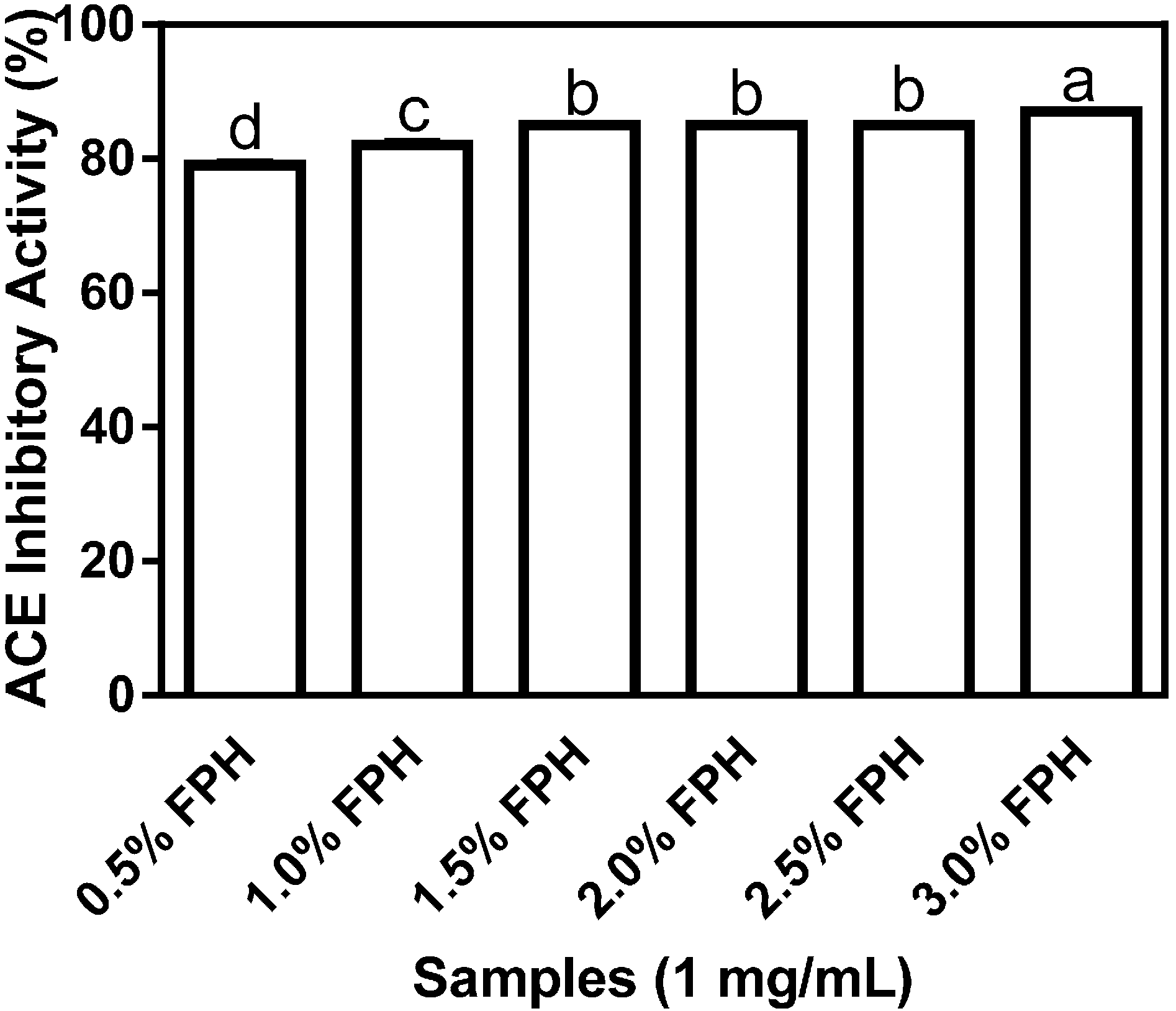
Figure 1 shows the differential inhibition of ACE by flaxseed protein hydrolysates (FPH) samples obtained following digestion with various concentrations of thermoase GL-30. The highest inhibitory value of 87% was recorded for the FPH sample obtained at the 3% thermoase concentration and this level of ACE inhibition was significantly (
p < 0.05) higher than the values obtained for all the other protein hydrolysate samples. While the increases in enzyme concentration from 0.5% to 1%, and from 1% to 1.5% resulted in statistically significant differences in the percentage of ACE inhibition, neither increasing the enzyme concentration from 1.5% to 2%, nor from 2% to 2.5% produced any difference in ACE inhibition of statistical significance. The results suggest that increasing enzyme concentration between 0.5% and 1.5% was important for the production of antihypertensive peptides whereas a similar increase in enzyme concentration between 1.5% and 2% yielded negligible or no differences in ACE-inhibiting peptides.
Figure 1.
Percentage (mean ± standard error) angiotensin I-converting enzyme- (ACE-) inhibitory activity of flaxseed protein hydrolysates (FPH) digested with thermoase GL-30 (0.5%–3%). Bars with different letters have mean values that are significantly different (p < 0.05).
Figure 1.
Percentage (mean ± standard error) angiotensin I-converting enzyme- (ACE-) inhibitory activity of flaxseed protein hydrolysates (FPH) digested with thermoase GL-30 (0.5%–3%). Bars with different letters have mean values that are significantly different (p < 0.05).
Flaxseed proteins are known to have a high concentration of BCAA [
17], which have been shown to be important in the inhibition of ACE activity [
18], and thus in potentiating the antihypertensive property of bioactive peptides. Since thermoase GL-30 is an isoform of thermolysin which is known for its specificity in cleaving at the
N-terminal regions of aromatic, bulky and hydrophobic amino acid residues and thus releasing BCAA (leucine, isoleucine and valine) [
18,
19], it was carefully chosen to yield a high proportion of BCAA-enriched hydrolysates. Interestingly, the FPH sample at the 3% enzyme concentration which inhibited ACE activity the most has the highest percentage (15.80%) of BCAA as shown in
Table 1 which is comparable to the BCAA content of 16.79% obtained by Udenigwe and Aluko [
18] after sequential hydrolysis of flaxseed protein isolate (FPI) with thermolysin and pronase. Given the previously reported capacity of hydrophobic amino acids (HAA) for inhibiting ACE activity [
9,
20], their relatively high content in all the FPH samples suggests important
in vitro antihypertensive properties and a potential role in the reduction of elevated blood pressure. Percentage ACE-inhibitory activity by the six different FPH samples, which ranged from 79% to 87% is comparable to the percentage ACE-inhibitory activities of about 84% and 82%, respectively, at the same final assay concentration of 1 mg/mL reported for ACE inhibition by thermolysin-digested (pH 8.0, 50 °C, 4 h) rapeseed protein samples [
21] and pepsin-digested (pH 3.0, 37 °C, 4 h) canola protein samples [
20] using the
N-(3-[2-furyl]acryloyl)-phenylalanylglycylglycine (FAPGG)-based spectrophotometric method (described in
Section 3.2.3.). However, the observed values in this work are significantly (
p < 0.05) higher than the 70.4% ± 0.4% ACE inhibition obtained using a hippuryl-
l-histidyl-
l-leucine (HHL)-based chromatographic method, which was reported for thermolysin-digested (pH 8.0, 60 °C, 3 h) rapeseed protein samples [
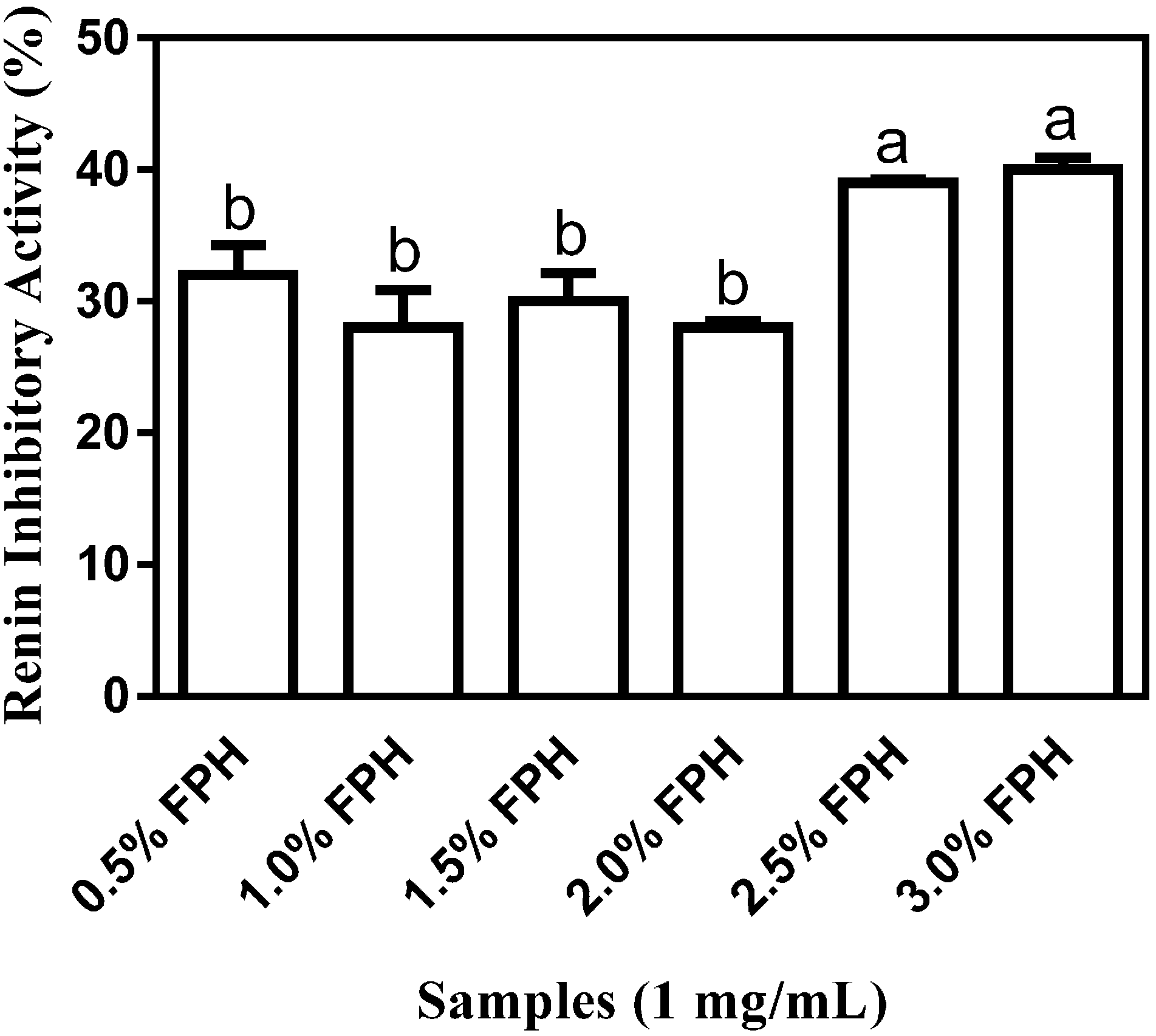
10]. All the FPH samples showed at least 28.0% ± 0.46% renin inhibition (
Figure 2) with the percentages of renin-inhibitory activities of 39% and 40% at the 2.5% and 3% enzyme concentrations, respectively, being significantly (
p < 0.05) higher than the
in vitro percentage inhibition ranging from 28% to 32% which was observed at the 0.5%–2% enzyme concentrations. The highest renin-inhibitory activity of 40.0% ± 0.94% which was observed with FPH at the 3% enzyme concentration suggests that just as with ACE inhibition, increasing the concentration of thermoase to 3% resulted in a marked increase in renin inhibition, probably because greater amounts of peptides were liberated from the native protein at higher enzyme concentration.
Figure 2.
Percentage (mean ± standard error) renin-inhibitory activity of flaxseed protein hydrolysates (FPH) after sample hydrolysis with a range (0.5%–3%) of thermoase GL-30 concentrations. Bars with different letters have mean values that are significantly different (p < 0.05).
Figure 2.
Percentage (mean ± standard error) renin-inhibitory activity of flaxseed protein hydrolysates (FPH) after sample hydrolysis with a range (0.5%–3%) of thermoase GL-30 concentrations. Bars with different letters have mean values that are significantly different (p < 0.05).
Table 1.
Percentage amino acid (AA) compositions of flaxseed protein meal (FPM), isolate (FPI), and hydrolysates (FPH) samples digested at different thermoase-GL 30 concentrations (0.5%–3%).
Table 1.
Percentage amino acid (AA) compositions of flaxseed protein meal (FPM), isolate (FPI), and hydrolysates (FPH) samples digested at different thermoase-GL 30 concentrations (0.5%–3%).
| AA | FPM | FPI | 0.5% FPH | 1.0% FPH | 1.5% FPH | 2.0% FPH | 2.5% FPH | 3.0% FPH |
|---|
| ASX | 9.76 | 9.39 | 11.61 | 11.53 | 11.01 | 11.74 | 11.59 | 11.41 |
| THR | 3.70 | 3.24 | 3.50 | 3.51 | 3.44 | 3.55 | 3.50 | 3.37 |
| SER | 5.05 | 4.73 | 5.21 | 5.19 | 5.14 | 5.29 | 5.19 | 4.92 |
| GLX | 20.92 | 23.44 | 22.18 | 21.47 | 22.56 | 21.91 | 21.59 | 21.44 |
| PRO | 3.95 | 3.73 | 4.49 | 4.43 | 4.29 | 4.50 | 4.31 | 4.36 |
| GLY | 6.14 | 6.00 | 5.47 | 5.37 | 5.56 | 5.41 | 5.34 | 5.27 |
| ALA | 4.59 | 4.04 | 4.52 | 4.56 | 4.28 | 4.57 | 4.51 | 4.54 |
| CYS | 1.80 | 2.13 | 1.35 | 1.27 | 1.58 | 1.34 | 1.35 | 1.32 |
| VAL | 5.17 | 4.85 | 5.02 | 5.18 | 4.97 | 5.07 | 5.02 | 5.53 |
| MET | 1.70 | 1.77 | 1.22 | 1.20 | 1.28 | 1.09 | 1.17 | 1.14 |
| ILE | 4.15 | 3.80 | 4.19 | 4.35 | 4.11 | 4.19 | 4.16 | 4.61 |
| LEU | 5.97 | 5.62 | 5.57 | 5.71 | 5.63 | 5.71 | 5.64 | 5.66 |
| TYR | 2.29 | 2.35 | 2.45 | 2.52 | 2.51 | 2.54 | 2.50 | 2.45 |
| PHE | 4.73 | 4.82 | 5.11 | 5.21 | 5.11 | 5.18 | 5.10 | 5.19 |
| HIS | 2.45 | 2.33 | 2.34 | 2.40 | 2.30 | 2.37 | 2.36 | 2.34 |
| LYS | 4.18 | 3.81 | 3.23 | 3.17 | 3.29 | 3.20 | 3.21 | 3.19 |
| ARG | 10.63 | 11.59 | 11.82 | 11.91 | 11.32 | 11.62 | 11.98 | 11.83 |
| TRP | 1.40 | 1.45 | 1.29 | 1.31 | 1.39 | 1.37 | 1.38 | 1.34 |
| AAA | 8.42 | 8.62 | 8.85 | 9.03 | 9.01 | 9.08 | 8.97 | 8.98 |
| BCAA | 15.29 | 14.27 | 14.79 | 15.23 | 14.71 | 14.97 | 14.81 | 15.80 |
| HAA | 35.75 | 34.55 | 35.22 | 35.72 | 35.15 | 35.55 | 35.11 | 36.13 |
The afore-stated inhibition values of
in vitro renin activity at the 2.5% and 3% enzyme concentrations are comparable to the values of about 49% and 40%, respectively, which were reported for renin inhibition at the same final assay concentration of 1 mg/mL by trypsin-digested canola proteins [
20] and rapeseed protein sequentially digested by 4% pepsin-pancreatin [
21]. However, while >2.5 mg/mL of 1% thermolysin-digested flaxseed proteins was reported to accomplish 50%
in vitro inhibition of human recombinant renin in a previous study [
6], the concentration of 1 mg/mL used in the present study resulted in ≈40% renin inhibitory activity at the 2.5% and 3% enzyme concentrations. Higher renin-inhibitory activity values of about 45%, 52% and 82%, respectively, have been reported for rapeseed protein hydrolysates produced from 4% thermolysin, 4% proteinase K, and 4% alcalase digestion [
21]. However, a 4% alcalase protein hydrolysate of African yam bean was reported to have a renin inhibition value of about 38% which is similar to the results for the 2.5% and 3% thermoase enzyme concentrations in the present study [
22]. Furthermore, significantly (
p < 0.05) lower values than those observed for the 2.5% and 3% thermoase concentrations in this study have been reported for 2% alcalase-hydrolysed chicken thigh skin protein (≈16% inhibition), 3% alcalase-hydrolysed chicken breast protein (≈24%) and 4% alcalase hydrolysed chicken breast protein (≈9%) as well as chicken breast skin protein sequentially hydrolysed with 4% pepsin-pancreatin (≈14%) [
12]. As was suggested by Udenigwe
et al. [
6], the considerably lower inhibition of renin by enzyme-derived food protein hydrolysates in comparison to their ACE inhibition could be as a result of the presence of more ACE-inhibitory peptides than similar renin inhibitors in enzymatically-digested food proteins thus resulting in a relatively easier inhibition of ACE than renin.
The FPH samples obtained at the 2.5% and 3% enzyme concentrations showed the highest inhibition of ACE and renin and were consequently subjected to membrane fractionation as well as used for in vivo systolic blood pressure measurements in spontaneously hypertensive rats.
2.2. ACE and Renin Inhibition by FPH Membrane Fractions
FPH samples obtained using 2.5% (2.5% FPH) or 3% (2.5% FPH) thermoase concentration were separated by membrane ultrafiltration and the defined peptide size fractions used for
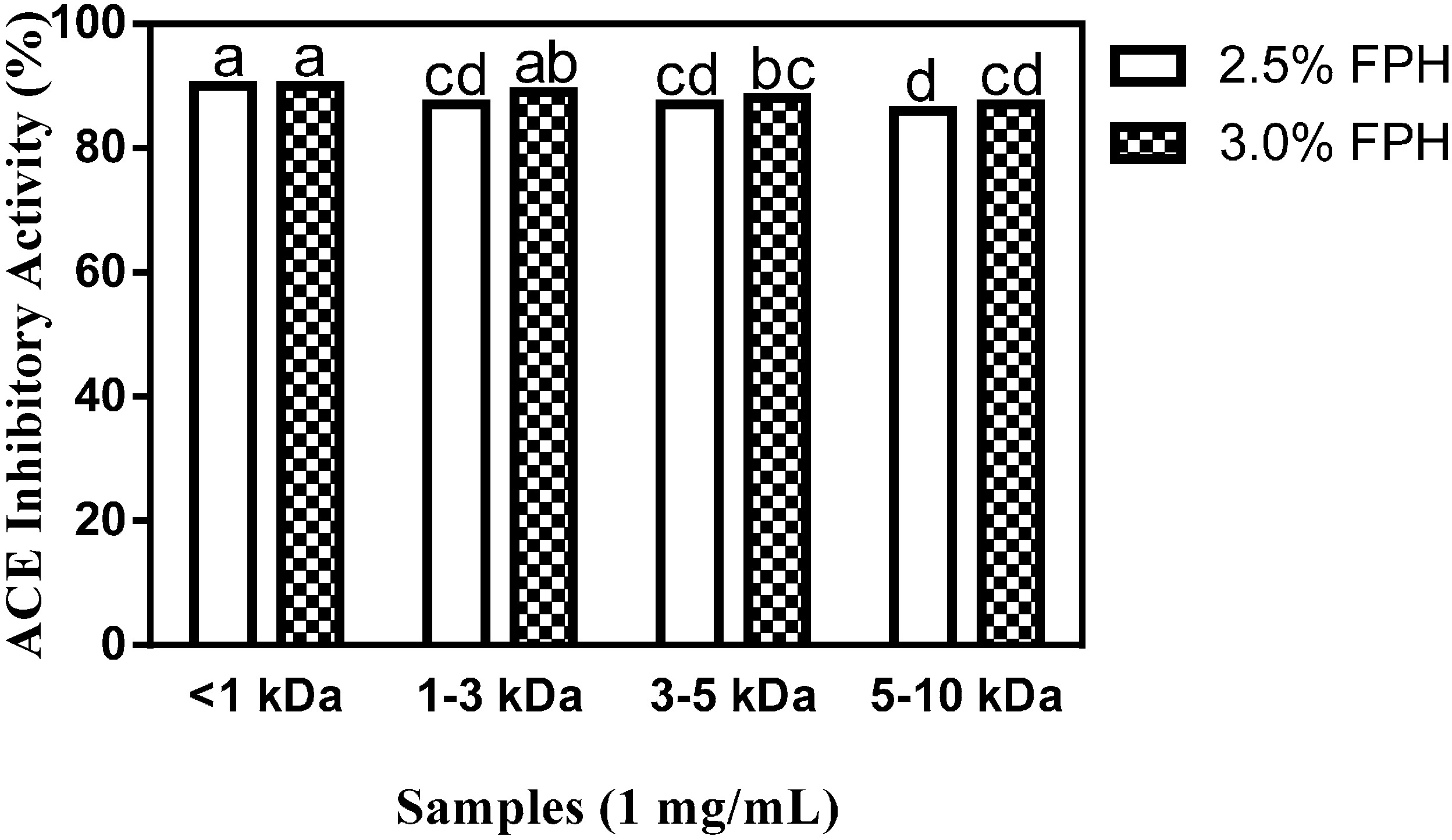
in vitro ACE and renin inhibition tests. As shown in
Figure 3, membrane ultrafiltration influenced the antihypertensive properties of the <1 kDa FPH fractions at both the 2.5% and 3% enzyme concentrations as evidenced by the considerable increase in ACE inhibition (up to 90%) relative to the inhibition of ACE activity by the unfractionated protein hydrolysate samples (
Section 2.1). This level of ACE inhibition by the <1 kDa peptide fraction is the highest recorded both for the unfractionated protein hydrolysate and the membrane fractions, and is significantly (
p < 0.05) higher than the ACE-inhibitory activity of the 1–3, the 3–5 and the 5–10 kDa fractions. No significant difference was observed in the ACE-inhibitory activity of the <1 kDa peptide fractions obtained from 2.5% FPH (2.5% FPH* < 1 kDa) or 3.0% FPH concentrations (3% FPH* < 1 kDa), which indicates that peptide size at the <1 kDa level played a greater role in ACE inhibition than the amount of enzyme used for proteolysis. For the 3 kDa molecular weight cut-off (MWCO) membrane, peptide fractions from the 3% enzyme-derived hydrolysate (3% FPH* 1–3 kDa) displayed a significantly (
p < 0.05) higher inhibition of ACE than the corresponding peptide fractions from the 2.5% enzyme hydrolysate (2.5% FPH* 1–3 kDa). As the MWCO increased from 3 to 10 kDa, the ability of the fractions to inhibit ACE activity markedly decreased although the amount of enzyme used for digestion remained the same. While increasing MWCO and the correspondingly larger peptide sizes clearly resulted in a decrease in peptide fraction ACE inhibitory potential, interestingly, there was no significant difference between the 5–10 kDa peptide fraction at the 3% enzyme concentration (3% FPH* 5–10 kDa) and those of 1–3 and 3–5 kDa at the 2.5% enzyme concentration (2.5% FPH* 1–3 kDa and 2.5% FPH* 3–5 kDa, respectively), which indicates a lack of influence at high enzyme concentrations.
The ACE inhibition of 90% shown by the two <1 kDa fractions in
Figure 3 is similar to previously reported results of ≈84% and 88%, respectively, for <1 kDa membrane fractions from 4% thermolysin- and 4% proteinase K-digested rapeseed proteins [
21], comparable to results obtained with peptide fractions of the same size from chicken thigh skin protein hydrolysate at 3% alcalase [
12] concentration (≈82% inhibition) but significantly (
p < 0.05) higher than the ≈78% inhibition of <1 kDa membrane fractions obtained following the hydrolysis of chicken breast skin protein with 1% pepsin-pancreatin [
12]. The 2.5% FPH* 1–3 KDa peptide fractions from the current work inhibited ACE activity to a degree similar to the 86% inhibition reported for peptide fractions of an identical size from 4% thermolysin-hydrolyzed rapeseed proteins but they were significantly (
p < 0.05) more potent ACE inhibitors than peptide fractions 1–3 kDa in size from both 3% alcalase-digested chicken thigh skin protein hydrolysate (with ACE inhibitory activity of ≈62%) and chicken breast skin protein hydrolysed using 1% pepsin-pancreatin (≈68% inhibition) [
12]. Similarly, at inhibition percentages ranging from 86% to 88%, the 3–5 and 5–10 kDa peptide fractions inhibited ACE activity at levels that are comparable to previously reported values of 82%–85% inhibition for 4% thermolysin-digested rapeseed protein hydrolysate fractions of similar sizes [
21] but showed significantly higher inhibitory activity than membrane fractions of identical sizes from chicken protein hydrolysate samples [
12]. Altogether, the most potent ACE-inhibitory fractions from this study (2.5% FPH* < 1 kDa and 3% FPH* < 1 kDa) showed a degree of inhibition higher than the maximum of 80% inhibition reported for arginine-rich flaxseed cationic fractions at a final assay concentration of 1 mg/mL [
23], which may be due to the presence of more hydrophobic and aromatic amino acid residues in the peptide fractions from the current study [
24] as shown in
Table 1.
Figure 3.
Percentage (mean ± standard error) of ACE-inhibitory activity of flaxseed protein hydrolysate (FPH) membrane ultrafiltration fractions after hydrolysis of isolated flaxseed proteins with 2.5% or 3.0% thermoase GL-30 and passing the hydrolysates through molecular weight cut-offs (MWCOs) of 1, 3, 5 and 10 kDa. Bars with different letters have mean values that are significantly different (p < 0.05).
Figure 3.
Percentage (mean ± standard error) of ACE-inhibitory activity of flaxseed protein hydrolysate (FPH) membrane ultrafiltration fractions after hydrolysis of isolated flaxseed proteins with 2.5% or 3.0% thermoase GL-30 and passing the hydrolysates through molecular weight cut-offs (MWCOs) of 1, 3, 5 and 10 kDa. Bars with different letters have mean values that are significantly different (p < 0.05).
As
Figure 4 shows, renin activity was moderately inhibited by virtually all the membrane fractions. While the renin-inhibitory activities of the 2.5% FPH* 3–5 kDa, 2.5% FPH* 5–10 kDa and 3% FPH* 5–10 kDa fractions ranged from 37% to 38%, the rest of the membrane fractions accomplished at least 40%
in vitro inhibition of renin activity. The results for membrane fractions that show <40% renin-inhibitory activity are similar to the 36% and 38% inhibition, respectively, reported at the same final assay concentration for the 3–5 kDa fractions from flavourzyme-hydrolyzed rapeseed proteins and 1–3 kDa membrane fractions from alcalase-hydrolyzed chicken thigh skin proteins. Similarly, the >40% renin-inhibitory values are comparable to previous results reported for <1 kDa rapeseed protein hydrolysates and 3–5 kDa chicken thigh skin protein hydrolysates [
12,
21]. Although no significant difference was observed in the inhibition of renin activity between the <1 and 1–3 kDa fractions at the two different enzyme concentrations used, at 40% renin inhibitory activity, the 3% FPH* 3–5 kDa fraction significantly (
p < 0.05) inhibited renin activity more than the 38% inhibition observed with the 2.5% FPH* 3–5 kDa fractions. This difference could be attributed to the release of a greater amount of peptides from the native protein structure at the higher enzyme concentration. The results generally suggest higher renin inhibitory activity by the smaller peptides than those larger in size—a trend which has been reported in previous studies [
9,
21].
Figure 4.
Percentage (mean ± standard error) renin-inhibitory activity of flaxseed protein hydrolysate (FPH) membrane ultrafiltration fractions after hydrolysis of isolated flaxseed proteins with 2.5% or 3.0% thermoase GL-30 and passing the hydrolysates through MWCOs of 1, 3, 5 and 10 kDa. Bars with different letters have mean values that are significantly different (p < 0.05).
Figure 4.
Percentage (mean ± standard error) renin-inhibitory activity of flaxseed protein hydrolysate (FPH) membrane ultrafiltration fractions after hydrolysis of isolated flaxseed proteins with 2.5% or 3.0% thermoase GL-30 and passing the hydrolysates through MWCOs of 1, 3, 5 and 10 kDa. Bars with different letters have mean values that are significantly different (p < 0.05).
Inhibition of renin activity by the 2.5% FPH* < 1 kDa and 3% FPH* < 1 kDa fractions is similar to the 39% and 40% inhibition, respectively, which were previously reported for membrane fractions that contain an identical peptide size obtained from chicken thigh skin proteins digested with 3% alcalase [
12] and rapeseed proteins digested with 4% alcalase [
21]. The 2.5% FPH* 1–3 kDa and 3% FPH* 1–3 kDa fractions also inhibited renin activity to a degree similar to the 39% renin inhibitory activity accomplished by peptide membrane fractions of an identical size from rapeseed proteins sequentially digested with 4% pepsin and pancreatin [
21]. However, the 49% and 57% reported inhibition of renin activity by 1–3 kDa membrane fractions from 4% alcalase- and 4% proteinase K-digested rapeseed proteins respectively are considerably higher than the inhibitory activity shown by fractions of identical size (2.5% FPH* 1–3 kDa and 3% FPH* 1–3 kDa) from the current study [
12]. Inhibition of renin activity by the 2.5% FPH* 3–5 kDa and 3% FPH* 3–5 kDa fractions are comparable to the 38% and 42% inhibition respectively reported for fractions of corresponding size obtained from 4% flavourzyme and 4% alcalase hydrolysates of rapeseed [
21]. However, the renin-inhibitory activity of 2.5% FPH* 5–10 kDa and 3% FPH* 5–10 kDa fractions was each considerably higher than the 24%, 26% and 23% inhibition respectively accomplished by peptide fractions of the same size from 4% alcalase, 4% pepsin-pancreatin and 4% flavourzyme hydrolysate fractions of rapeseed proteins but substantially lower than the inhibition of 55% and 57% correspondingly shown by 4% proteinase K- and 4% thermolysin-digested rapeseed protein hydrolysate fractions of the same size [
21]. A previous work that screened cationic membrane fractions from flaxseed proteins sequentially digested with trypsin and pronase reported a maximum renin inhibition of 44.5% at a peptide final assay concentration of 0.75 mg/mL. In the current work, although the FPH peptide fractions were used at a slightly higher final assay concentration of 1 mg/mL in a similarly-designed assay, renin-inhibitory values akin to that of the previous work were obtained [
23]; the differences may be attributed to variations in the type of amino acid residues present in both fractions [
23].
2.3. In Vivo Antihypertensive Activity of FPH and FPH Membrane Fractions
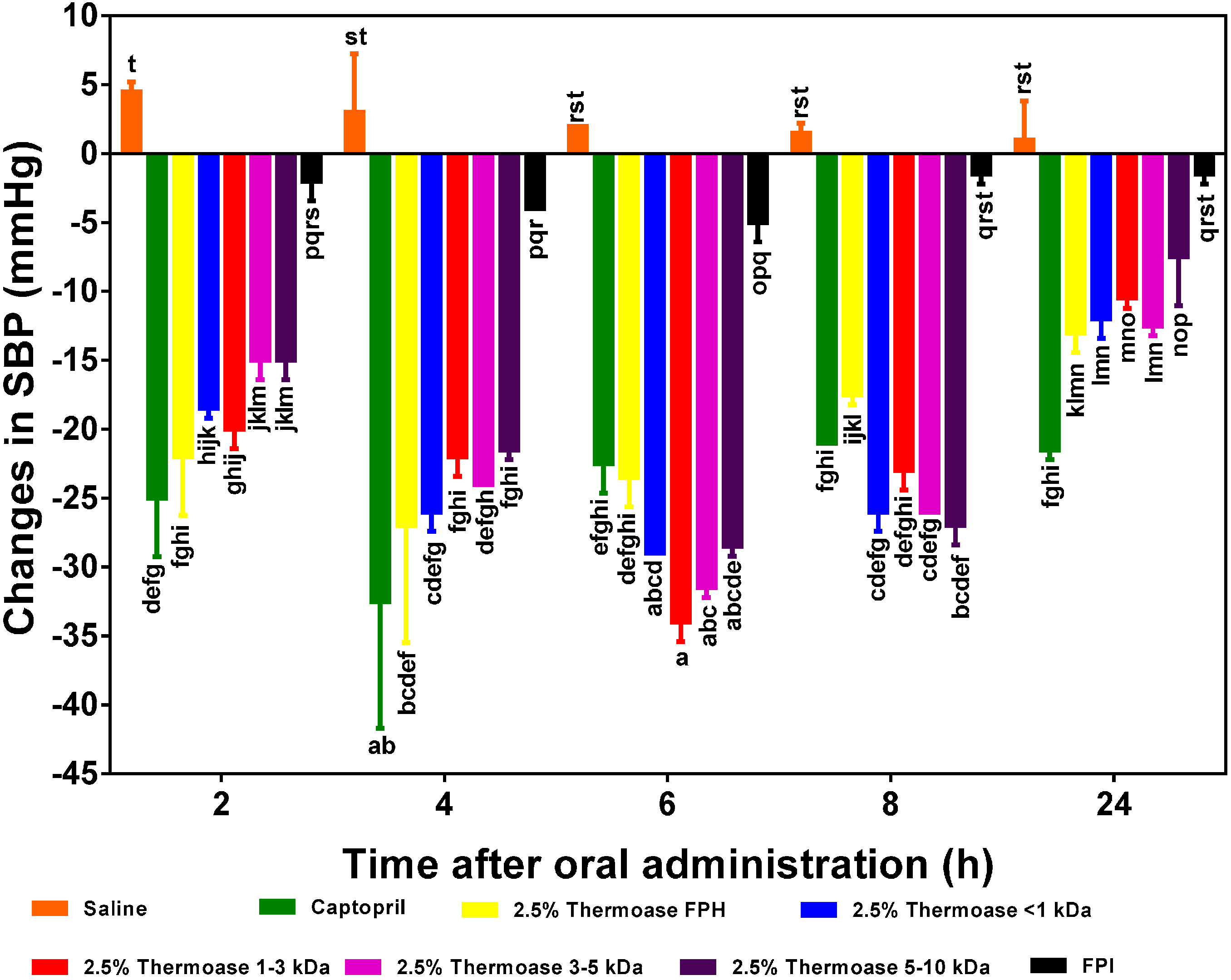
As illustrated in
Figure 5,
Figure 6 and
Figure 7, all the FPH and membrane fraction samples administered to the spontaneously hypertensive rats (SHR) significantly (
p < 0.05) lowered systolic blood pressure (SBP) at all measured time points better than phosphate buffered saline (PBS), thus providing
prima facie evidence and preliminary confirmation that the test samples do indeed possess
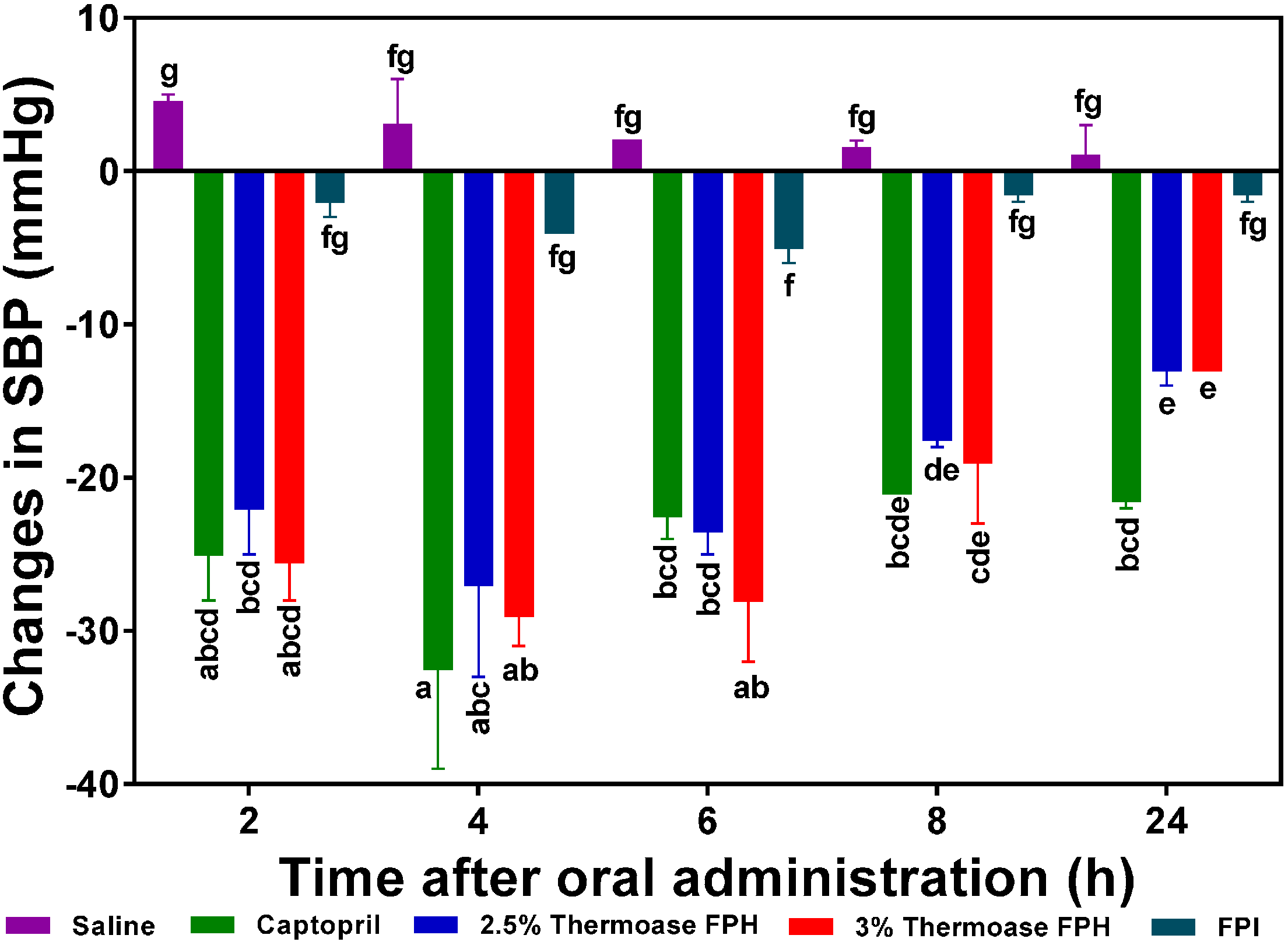
in vivo antihypertensive effects. The protein hydrolysate which showed the highest SBP reduction (−29 mm Hg after 4 h) was the 3% FPH sample while the unhydrolyzed flaxseed protein isolate (FPI) had the least BP-lowering activity (−4 to −5 mm Hg after 4–6 h) as shown in
Figure 5. The SBP-lowering activities of the 2.5% FPH and 3% FPH samples after 2–8 h were comparable to that of captopril (used as a positive control and at a dosage 20 times lower), thus suggesting the potential of the peptide samples to rapidly lower blood pressure on a short term basis. However, the blood pressure-lowering ability of captopril was significantly (
p < 0.05) better than those of the protein hydrolysates after 24 h of oral administration. Although the hypotensive effect of FPI slightly increased from −2 mm Hg after 2 h to ≈−4 to −5 mm Hg after 4–6 h, the two FPH samples showed significantly (
p < 0.05) higher SBP decreases than the FPI sample at every time point. The results support the fundamental principle that protein hydrolysates contain more rapidly absorbable peptides than the unhydrolysed protein isolate. Additionally, since proteolysis of native proteins must take place before absorption of any antihypertensive peptides, it is logical to expect weak FPI antihypertensive effect on a short-term basis when compared to FPH samples that contain predigested peptides.
The SBP-lowering activity of the two hydrolysate samples were similar but had diminished considerably after 24 h post-oral administration with a maximum value of −13 mmHg, which is still significantly (p < 0.05) higher than the maximum effect (−5 mmHg after 6 h) of the unhydrolysed protein (FPI).
In
Figure 6 and
Figure 7, the 2.5% FPH and 3% FPH samples both showed significantly (
p < 0.05) larger decreases in SBP than some of the corresponding membrane permeates (3–5 and 5–10 kDa) after 2 and 4 h. However, at both the 6 and 8 h marks, the SBP-reducing effect of some of the membrane permeates (1–3 and 3–5 kDa) was significantly (
p < 0.05) greater than that of their corresponding protein hydrolysates. This could be as a result of the fact that the membrane fractions had longer intrinsic synergistic effects or less antagonistic effects due to the higher level of peptide homogeneity when compared to the high peptide heterogeneity in the unfractionated protein hydrolysates. Therefore, the protein hydrolysates may be useful ingredients to formulate fast-acting antihypertensive products while the membrane fractions would be suitable for extended-release type of antihypertensive formulations.
Figure 5.
Effects of 2.5% and 3% enzymatic flaxseed protein hydrolysate (FPH), and flaxseed protein isolate (FPI) samples on systolic blood pressure (SBP) of spontaneously hypertensive rats (SHR) after oral gavage. Rats were administered with FPH and FPI using a dose of 200 mg protein/kg rat body weight (BW) while the positive control, captopril was given at 10 mg/kg body weight. Saline was used as negative control. Bars with different letters have mean SBP values that are significantly (p < 0.05) different.
Figure 5.
Effects of 2.5% and 3% enzymatic flaxseed protein hydrolysate (FPH), and flaxseed protein isolate (FPI) samples on systolic blood pressure (SBP) of spontaneously hypertensive rats (SHR) after oral gavage. Rats were administered with FPH and FPI using a dose of 200 mg protein/kg rat body weight (BW) while the positive control, captopril was given at 10 mg/kg body weight. Saline was used as negative control. Bars with different letters have mean SBP values that are significantly (p < 0.05) different.
Figure 6.
Effects of 2.5% enzymatic flaxseed protein hydrolysate (FPH), membrane ultrafiltration peptide fractions and flaxseed protein isolate (FPI) on systolic blood pressure (SBP) of spontaneously hypertensive rats (SHR) after oral gavage. Rats were administered with FPH, FPH fractions and FPI using a dose of 200 mg protein/kg rat body weight (BW) while the positive control, captopril was given at 10 mg/kg body weight. Saline was used as the negative control. Bars with different letters have mean SBP values that are significantly (p < 0.05) different.
Figure 6.
Effects of 2.5% enzymatic flaxseed protein hydrolysate (FPH), membrane ultrafiltration peptide fractions and flaxseed protein isolate (FPI) on systolic blood pressure (SBP) of spontaneously hypertensive rats (SHR) after oral gavage. Rats were administered with FPH, FPH fractions and FPI using a dose of 200 mg protein/kg rat body weight (BW) while the positive control, captopril was given at 10 mg/kg body weight. Saline was used as the negative control. Bars with different letters have mean SBP values that are significantly (p < 0.05) different.
Among the peptide fractions from the 2.5% FPH sample (
Figure 6), the hypotensive activities of 2.5% FPH* < 1 kDa and 2.5% FPH* 1–3 kDa tended to be the highest after 2 h indicating their possible ability to lower blood pressure on a short term basis but also suggesting that the larger peptides in the 5 and 10 kDa fractions might not have been as efficiently absorbed as the smaller <1 and 1–3 kDa peptides in such a short amount of time. Alternatively, it is possible that the longer peptides undergo initial hydrolysis to release active fragments that are then subsequently absorbed; this will delay the antihypertensive effect. The significant (
p < 0.05) increases in the BP-reducing effect of the larger peptides (3–5 and 5–10 kDa) after 4 and 6 h lends credence to this reasoning. Remarkably, all the membrane fractions from both protein hydrolysate samples maintained significant hypotensive effects after 24 h (approximately −8 mm Hg in the least).
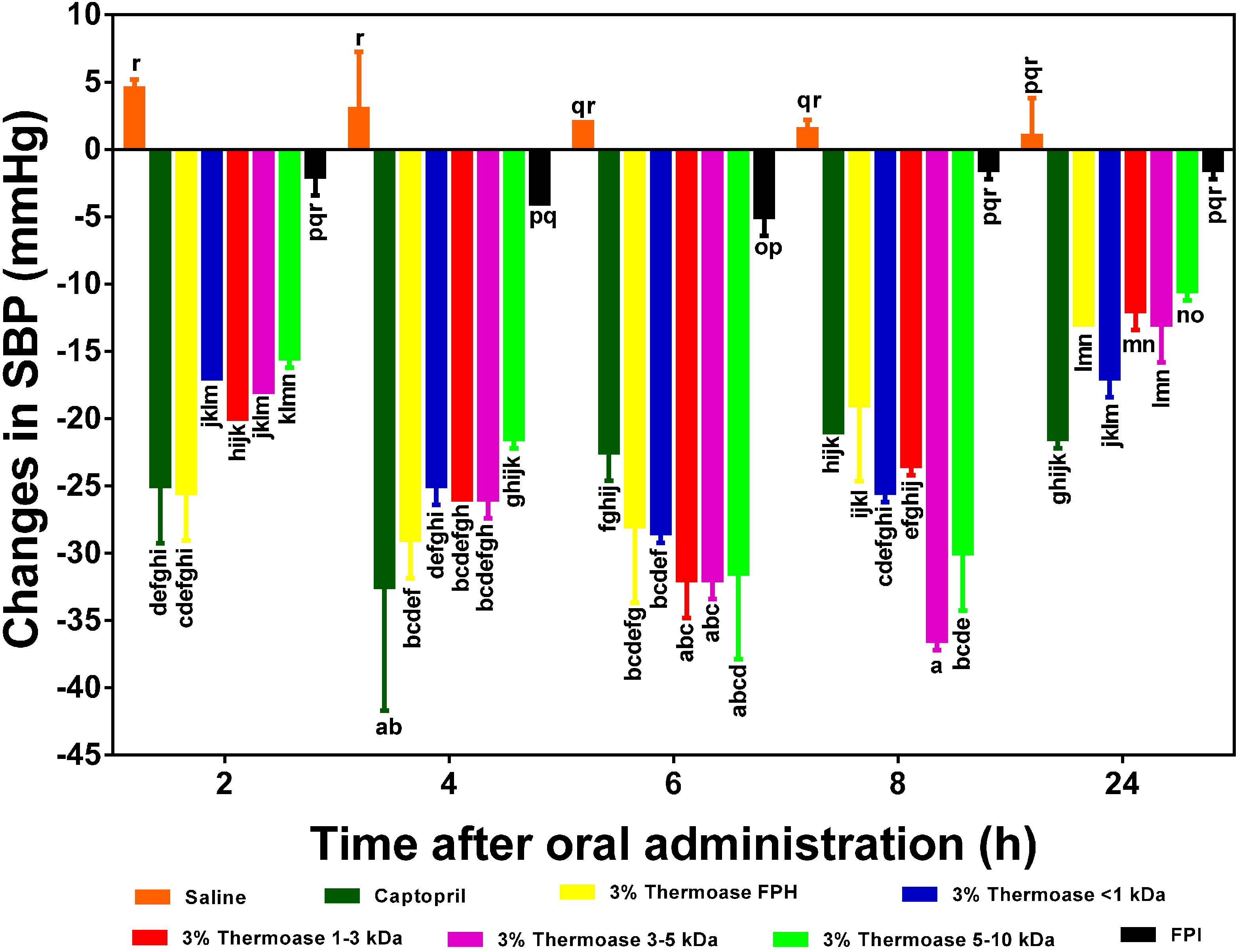
Figure 7.
Effects of 3% enzymatic flaxseed protein hydrolysate (FPH), membrane ultrafiltration peptide fractions and flaxseed protein isolate (FPI) samples on systolic blood pressure (SBP) of spontaneously hypertensive rats (SHR) after oral gavage. Rats were administered with FPH, FPH fractions and FPI using a dose of 200 mg protein/kg rat body weight (BW) while the positive control, captopril was given at 10 mg/kg body weight. Saline was used as a blank. Bars with different letters have mean SBP values that are significantly (p < 0.05) different.
Figure 7.
Effects of 3% enzymatic flaxseed protein hydrolysate (FPH), membrane ultrafiltration peptide fractions and flaxseed protein isolate (FPI) samples on systolic blood pressure (SBP) of spontaneously hypertensive rats (SHR) after oral gavage. Rats were administered with FPH, FPH fractions and FPI using a dose of 200 mg protein/kg rat body weight (BW) while the positive control, captopril was given at 10 mg/kg body weight. Saline was used as a blank. Bars with different letters have mean SBP values that are significantly (p < 0.05) different.
Contrary to what was observed with the 2.5% FPH fractions, the 3–5 and 5–10 kDa fractions of the 3.5% FPH sample (
Figure 7) were very effective in lowering elevated blood pressure in SHR. In fact, unlike in the fractions from the 2.5% FPH sample where 2.5% FPH* 1–3 kDa was responsible for the largest decrease in blood pressure (−34 mm Hg) after 6 h, 3% FPH* 3–5 kDa exerted the highest blood pressure-lowering effect of −37 mm Hg after 8 h among the fractions from the 3% FPH sample. This difference could be as a result of, among other things, the difference in the concentration of peptides liberated by the different enzyme concentrations and additional processing of the peptides in the gastrointestinal tract of the SHR.
Similar results have been obtained with the enzymatic hydrolysate samples of hemp seed protein hydrolysate [
9] (after 2 h in comparison to 2.5% FPH at the same time point) and pepsin-digested canola protein hydrolysate after 8 h [
20] (compared to 3% FPH after the same number of hours). Although a reduction in SBP of about −15 mm Hg, which is comparable to that of 2.5% FPH after 8 h, was reported with an oral dose of sweet potato protein hydrolysate [
25] lower than the 200 mg/kg BW used in this study at the same hour mark, the SBP-lowering effects of the flaxseed protein hydrolysate and membrane fractions used in this study lasted longer (an average of −12 mm Hg after 24 h) when compared to that of the enzyme-digested sweet potato proteins (nearly zero after 24 h).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




