Squamosamide Derivative FLZ Protects Retinal Pigment Epithelium Cells from Oxidative Stress through Activation of Epidermal Growth Factor Receptor (EGFR)-AKT Signaling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. FLZ Protects Retinal Pigment Epithelium (RPE) Cells from Hydrogen Peroxide (H2O2)

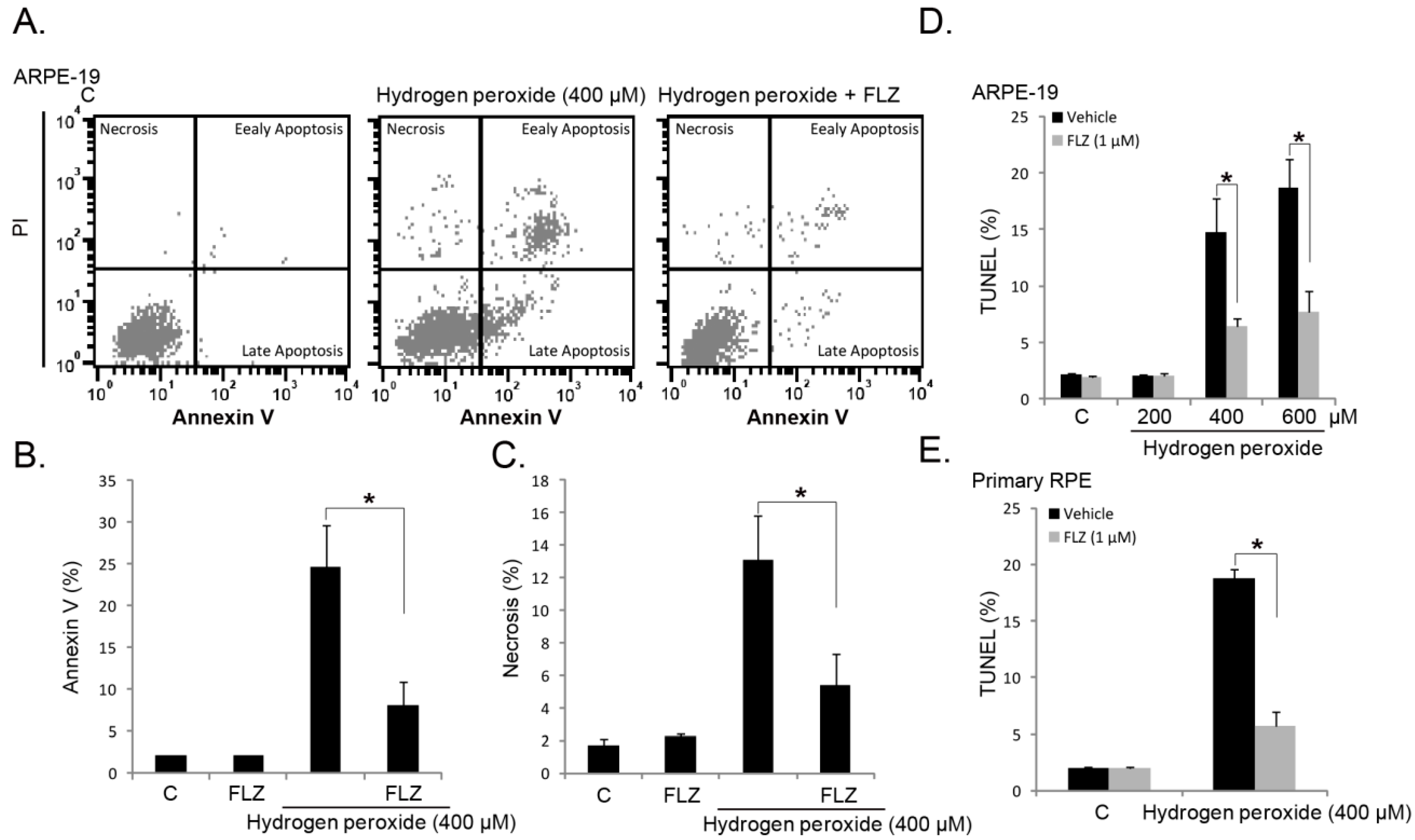
2.2. FLZ Attenuates H2O2-Induced RPE Cell Apoptosis

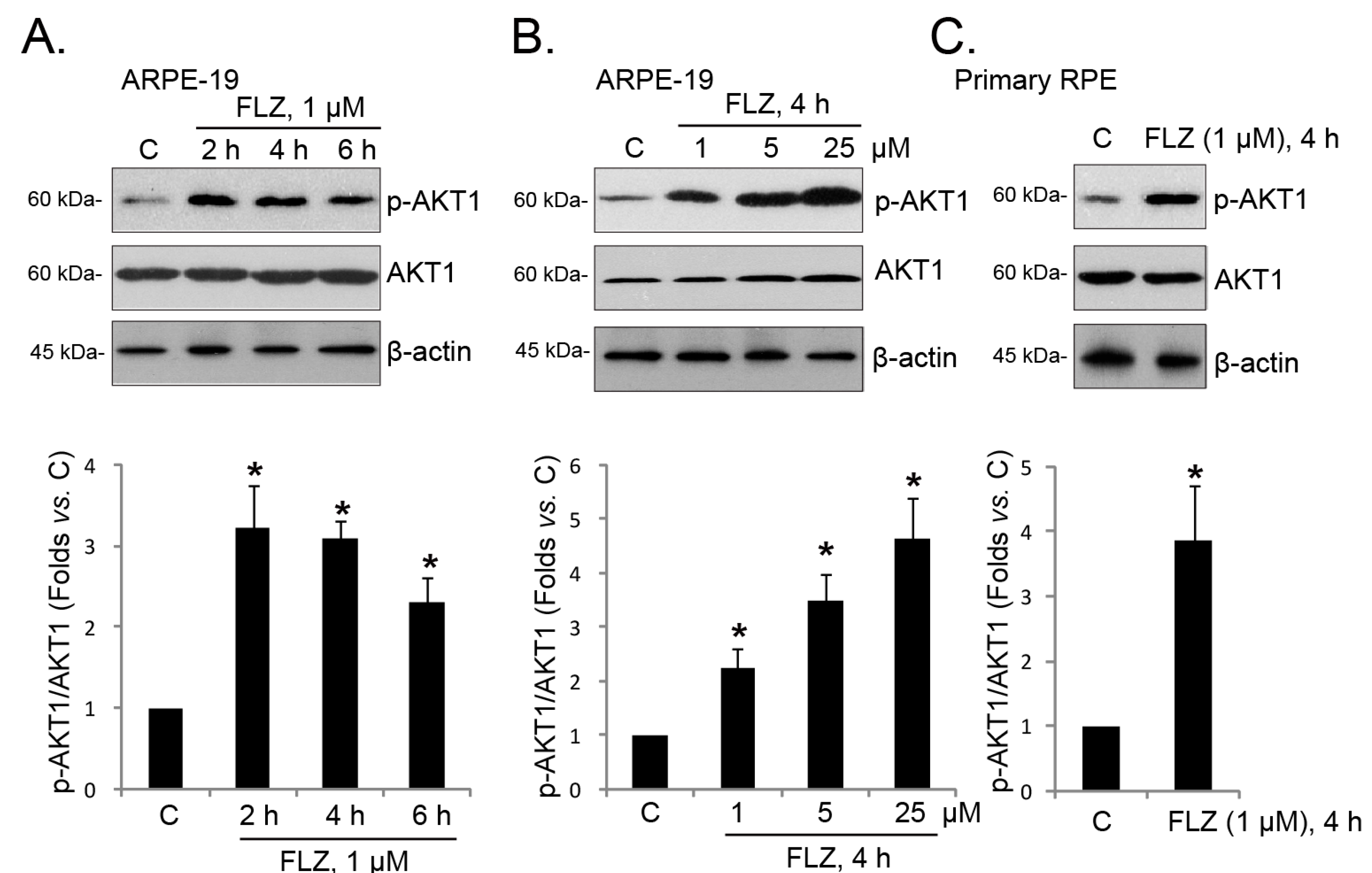
2.3. FLZ Activates AKT in RPE Cells

2.4. AKT Activation Mediates the FLZ-Induced Pro-Survival Effect against H2O2

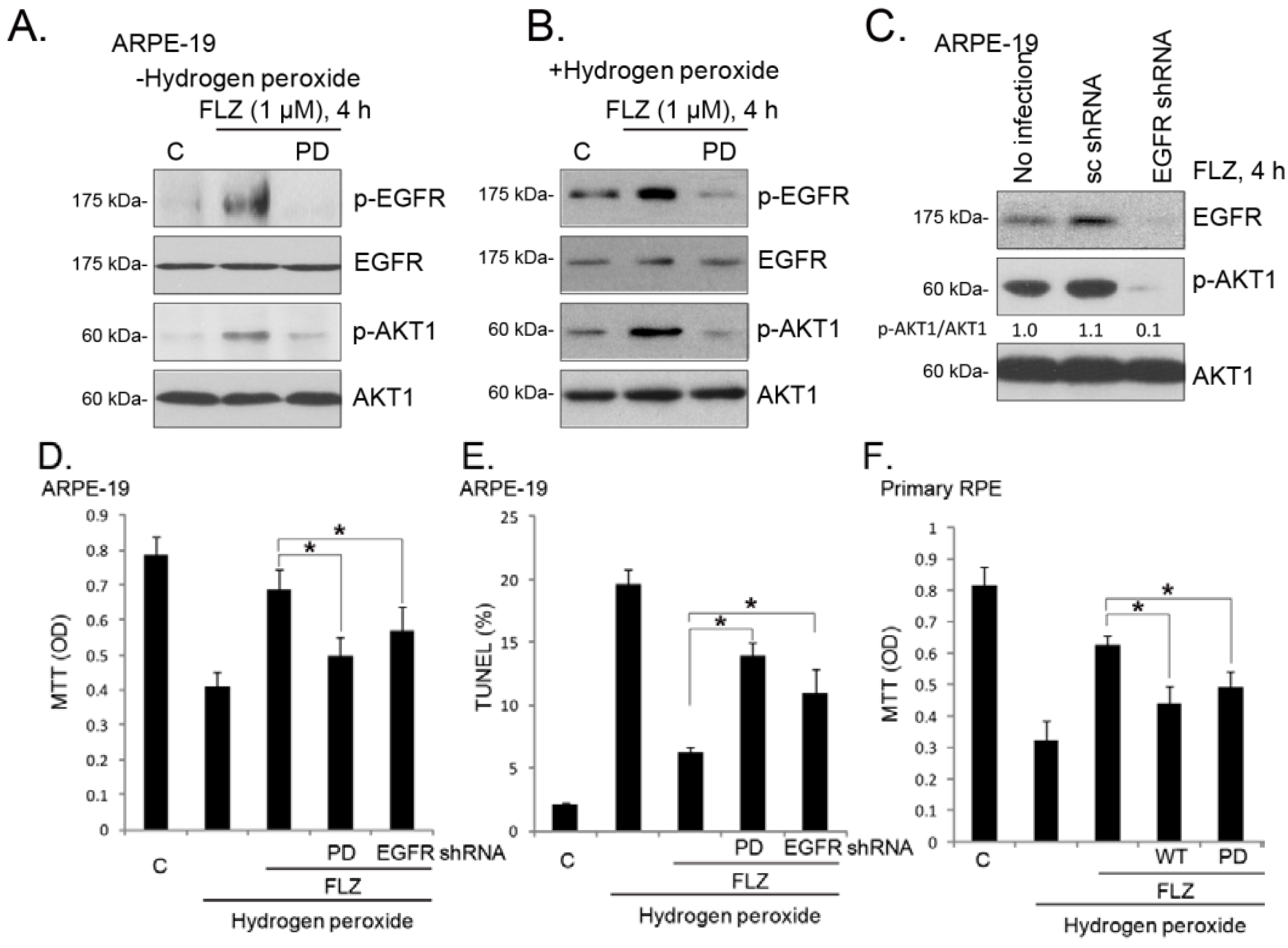
2.5. EGFR (Epidermal Growth Factor Receptor) Trans-Activation Mediates FLZ-Induced AKT Activation and Pro-Survival Effect in RPE Cells


2.6. Discussion
3. Experimental Section
3.1. Chemical and Reagents
3.2. APRE-19 Cell Culture
3.3. Primary Mouse RPE Cell Isolation and Culture
3.4. Cell Viability Assay
3.5. Western Blotting
3.6. Annexin-V/PI FACS (Fluorescence-Activated Cell Sorting) Assay
3.7. TUNEL (Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling) Staining
3.8. Stable AKT1 Knockdown by Short Hairpin RNA (shRNA)
3.9. Transit Knockdown of Epidermal Growth Factor Receptor (EGFR) by shRNA
3.10. Statistical Analysis
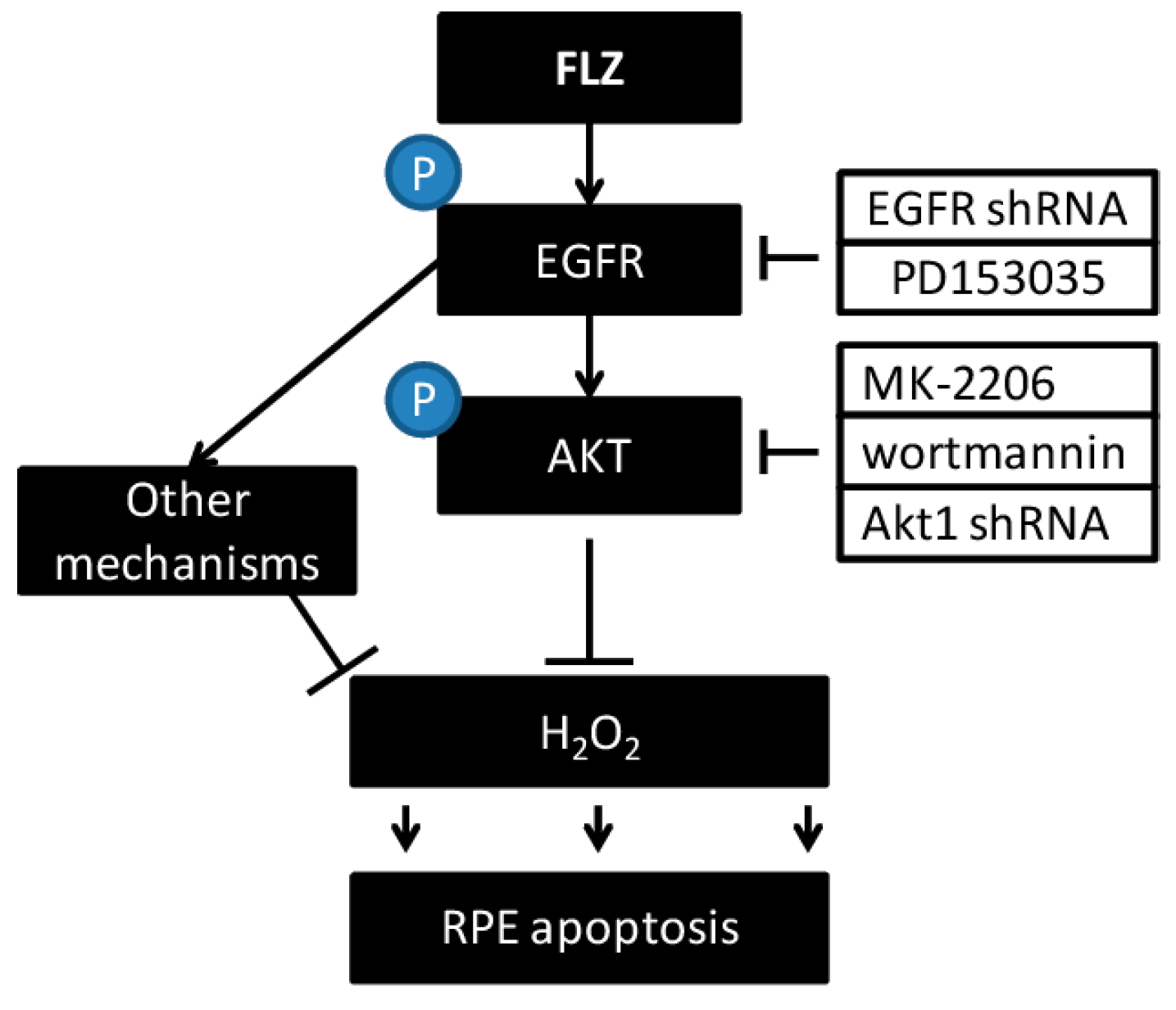
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Friedman, D.S.; O’Colmain, B.J.; Munoz, B.; Tomany, S.C.; McCarty, C.; de Jong, P.T.; Nemesure, B.; Mitchell, P.; Kempen, J. Prevalence of age-related macular degeneration in the United States. Arch. Ophthalmol. 2004, 122, 564–572. [Google Scholar]
- Chen, Y.; Gibson, S.B. Is mitochondrial generation of reactive oxygen species a trigger for autophagy? Autophagy 2008, 4, 246–248. [Google Scholar]
- Pacifici, R.E.; Davies, K.J. Protein, lipid and DNA repair systems in oxidative stress: The free-radical theory of aging revisited. Gerontology 1991, 37, 166–180. [Google Scholar]
- Bartlett, H.; Eperjesi, F. A randomised controlled trial investigating the effect of nutritional supplementation on visual function in normal, and age-related macular disease affected eyes: Design and methodology [ISRCTN78467674]. Nutr. J. 2003, 2. [Google Scholar] [CrossRef] [Green Version]
- Richer, S.; Stiles, W.; Statkute, L.; Pulido, J.; Frankowski, J.; Rudy, D.; Pei, K.; Tsipursky, M.; Nyland, J. Double-masked, placebo-controlled, randomized trial of lutein and antioxidant supplementation in the intervention of atrophic age-related macular degeneration: The Veterans LAST study (Lutein Antioxidant Supplementation Trial). Optometry 2004, 75, 216–230. [Google Scholar]
- Cao, G.F.; Liu, Y.; Yang, W.; Wan, J.; Yao, J.; Wan, Y.; Jiang, Q. Rapamycin sensitive mTOR activation mediates nerve growth factor (NGF) induced cell migration and pro-survival effects against hydrogen peroxide in retinal pigment epithelial cells. Biochem. Biophys. Res. Commun. 2011, 414, 499–505. [Google Scholar]
- Yao, J.; Bi, H.E.; Sheng, Y.; Cheng, L.B.; Wendu, R.L.; Wang, C.H.; Cao, G.F.; Jiang, Q. Ultraviolet (UV) and hydrogen peroxide activate ceramide-ER stress-AMPK signaling axis to promote retinal pigment epithelium (RPE) cell apoptosis. Int. J. Mol. Sci. 2013, 14, 10355–10368. [Google Scholar]
- Zhang, H.; Liu, Y.Y.; Jiang, Q.; Li, K.R.; Zhao, Y.X.; Cao, C.; Yao, J. Salvianolic acid A protects RPE cells against oxidative stress through activation of Nrf2/HO-1 signaling. Free Radic. Biol. Med. 2014, 69, 219–228. [Google Scholar]
- Cheng, L.B.; Cheng, L.; Bi, H.E.; Zhang, Z.Q.; Yao, J.; Zhou, X.Z.; Jiang, Q. Alpha-melanocyte stimulating hormone protects retinal pigment epithelium cells from oxidative stress through activation of melanocortin 1 receptor-Akt-mTOR signaling. Biochem. Biophys. Res. Commun. 2014, 443, 447–452. [Google Scholar]
- Chen, S.H.; Li, D.L.; Yang, F.; Wu, Z.; Zhao, Y.Y.; Jiang, Y. Gemcitabine-induced pancreatic cancer cell death is associated with MST1/Cyclophilin D mitochondrial complexation. Biochimie 2014, 103, 71–79. [Google Scholar]
- Bao, X.Q.; Kong, X.C.; Kong, L.B.; Wu, L.Y.; Sun, H.; Zhang, D. Squamosamide derivative FLZ protected dopaminergic neuron by activating Akt signaling pathway in 6-OHDA-induced in vivo and in vitro Parkinson’s disease models. Brain Res. 1547, 49–57. [Google Scholar]
- Bao, X.Q.; Li, N.; Wang, T.; Kong, X.C.; Tai, W.J.; Sun, H.; Zhang, D. FLZ alleviates the memory deficits in transgenic mouse model of Alzheimer’s disease via decreasing beta-amyloid production and tau hyperphosphorylation. PLoS One 2013, 8, e78033. [Google Scholar]
- Bao, X.Q.; Kong, X.C.; Qian, C.; Zhang, D. FLZ protects dopaminergic neuron through activating protein kinase B/mammalian target of rapamycin pathway and inhibiting RTP801 expression in Parkinson’s disease models. Neuroscience 2012, 202, 396–404. [Google Scholar]
- Fang, F.; Liu, G.T. Novel squamosamide derivative (compound FLZ) attenuates Aβ25–35-induced toxicity in SH-SY5Y cells. Acta Pharmacol. Sin. 2008, 29, 152–160. [Google Scholar]
- Kong, X.C.; Zhang, D.; Qian, C.; Liu, G.T.; Bao, X.Q. FLZ, a novel HSP27 and HSP70 inducer, protects SH-SY5Y cells from apoptosis caused by MPP(+). Brain Res. 2011, 1383, 99–107. [Google Scholar]
- Zhang, D.; Zhang, J.J.; Liu, G.T. The novel squamosamide derivative FLZ protects against 6-hydroxydopamine-induced apoptosis through inhibition of related signal transduction in SH-SY5Y cells. Eur. J. Pharmacol. 2007, 561, 1–6. [Google Scholar]
- Kang, R.X.; Zhang, J.J. A natural squamosamide derivative FLZ inhibits homocysteine-induced rat brain microvascular endothelial cells dysfunction. Biochem. Biophys. Res. Commun. 2012, 417, 1176–1181. [Google Scholar]
- Song, G.; Ouyang, G.; Bao, S. The activation of Akt/PKB signaling pathway and cell survival. J. Cell Mol. Med. 2005, 9, 59–71. [Google Scholar]
- Vivanco, I.; Sawyers, C.L. The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat. Rev. Cancer 2002, 2, 489–501. [Google Scholar]
- Hennessy, B.T.; Smith, D.L.; Ram, P.T.; Lu, Y.; Mills, G.B. Exploiting the PI3K/AKT pathway for cancer drug discovery. Nat. Rev. Drug Discov. 2005, 4, 988–1004. [Google Scholar]
- Li, K.R.; Zhang, Z.Q.; Yao, J.; Zhao, Y.X.; Duan, J.; Cao, C.; Jiang, Q. Ginsenoside Rg-1 protects retinal pigment epithelium (RPE) cells from cobalt chloride (CoCl2) and hypoxia assaults. PLoS One 2013, 8, e84171. [Google Scholar]
- Ji, D.; Zhang, Z.; Cheng, L.; Chang, J.; Wang, S.; Zheng, B.; Zheng, R.; Sun, Z.; Wang, C.; Liu, R.; et al. The combination of RAD001 and MK-2206 exerts synergistic cytotoxic effects against PTEN mutant gastric cancer cells: Involvement of MAPK-dependent autophagic, but not apoptotic cell death pathway. PLoS One 2014, 9, e85116. [Google Scholar]
- Yap, T.A.; Yan, L.; Patnaik, A.; Fearen, I.; Olmos, D.; Papadopoulos, K.; Baird, R.D.; Delgado, L.; Taylor, A.; Lupinacci, L.; et al. First-in-man clinical trial of the oral pan-AKT inhibitor MK-2206 in patients with advanced solid tumors. J. Clin. Oncol. 2011, 29, 4688–4695. [Google Scholar]
- Brunn, G.J.; Williams, J.; Sabers, C.; Wiederrecht, G.; Lawrence, J.C., Jr.; Abraham, R.T. Direct inhibition of the signaling functions of the mammalian target of rapamycin by the phosphoinositide 3-kinase inhibitors, wortmannin and LY294002. EMBO J. 1996, 15, 5256–5267. [Google Scholar]
- Tomas, A.; Futter, C.E.; Eden, E.R. EGF receptor trafficking: Consequences for signaling and cancer. Trends Cell Biol. 2014, 24, 26–34. [Google Scholar]
- Cao, C.; Lu, S.; Jiang, Q.; Wang, W.J.; Song, X.; Kivlin, R.; Wallin, B.; Bagdasarian, A.; Tamakloe, T.; Chu, W.M.; et al. EGFR activation confers protections against UV-induced apoptosis in cultured mouse skin dendritic cells. Cell Signal. 2008, 20, 1830–1838. [Google Scholar]
- Maretzky, T.; Evers, A.; Zhou, W.; Swendeman, S.L.; Wong, P.M.; Rafii, S.; Reiss, K.; Blobel, C.P. Migration of growth factor-stimulated epithelial and endothelial cells depends on EGFR transactivation by ADAM17. Nat. Commun. 2011, 2. [Google Scholar] [CrossRef]
- Oganesian, A.; Yarov-Yarovoy, V.; Parks, W.C.; Schwinn, D.A. Constitutive coupling of a naturally occurring human alpha1a-adrenergic receptor genetic variant to EGFR transactivation pathway. Proc. Natl. Acad. Sci. USA 2011, 108, 19796–19801. [Google Scholar]
- Sur, S.; Agrawal, D.K. Transactivation of EGFR by G protein-coupled Receptor in the Pathophysiology of Intimal Hyperplasia. Curr. Vasc. Pharmacol. 2014, 12, 190–201. [Google Scholar]
- McElroy, S.J.; Hobbs, S.; Kallen, M.; Tejera, N.; Rosen, M.J.; Grishin, A.; Matta, P.; Schneider, C.; Upperman, J.; Ford, H.; et al. Transactivation of EGFR by LPS induces COX-2 expression in enterocytes. PLoS One 2012, 7, e38373. [Google Scholar]
- Cao, C.; Sun, Y.; Healey, S.; Bi, Z.; Hu, G.; Wan, S.; Kouttab, N.; Chu, W.; Wan, Y. EGFR-mediated expression of aquaporin-3 is involved in human skin fibroblast migration. Biochem. J. 2006, 400, 225–234. [Google Scholar]
- Qiu, L.; Zhou, C.; Sun, Y.; Di, W.; Scheffler, E.; Healey, S.; Wanebo, H.; Kouttab, N.; Chu, W.; Wan, Y. Paclitaxel and ceramide synergistically induce cell death with transient activation of EGFR and ERK pathway in pancreatic cancer cells. Oncol. Rep. 2006, 16, 907–913. [Google Scholar]
- Niwa, K.; Inanami, O.; Yamamori, T.; Ohta, T.; Hamasu, T.; Kuwabara, M. Redox regulation of PI3K/Akt and p53 in bovine aortic endothelial cells exposed to hydrogen peroxide. Antioxid. Redox Signal. 2003, 5, 713–722. [Google Scholar]
- Chen, K.; Thomas, S. R.; Albano, A.; Murphy, M. P.; Keaney, J. F., Jr. Mitochondrial function is required for hydrogen peroxide-induced growth factor receptor transactivation and downstream signaling. J. Biol. Chem. 2004, 279, 35079–35086. [Google Scholar]
- Radisavljevic, Z.M.; Gonzalez-Flecha, B. TOR kinase and Ran are downstream from PI3K/Akt in H2O2-induced mitosis. J. Cell Biochem. 2004, 91, 1293–1300. [Google Scholar]
- Young, R.W. Pathophysiology of age-related macular degeneration. Surv. Ophthalmol. 1987, 31, 291–306. [Google Scholar]
- Zarbin, M. Age-related macular degeneration: Review of pathogenesis. Eur. J. Ophthalmol. 1998, 8, 199–206. [Google Scholar]
- Roduit, R.; Schorderet, D.F. MAP kinase pathways in UV-induced apoptosis of retinal pigment epithelium ARPE19 cells. Apoptosis 2008, 13, 343–353. [Google Scholar]
- Liang, Y.G.; Jorgensen, A.G.; Kaestel, C.G.; Wiencke, A.K.; Lui, G.M.; la Cour, M.H.; Ropke, C.H.; Nissen, M.H. Bcl-2, Bax, and c-Fos expression correlates to RPE cell apoptosis induced by UV-light and daunorubicin. Curr. Eye Res. 2000, 20, 25–34. [Google Scholar]
- Nilsson, S.E.; Sundelin, S.P.; Wihlmark, U.; Brunk, U.T. Aging of cultured retinal pigment epithelial cells: Oxidative reactions, lipofuscin formation and blue light damage. Doc. Ophthal. Mol. 2003, 106, 13–16. [Google Scholar]
- Drobek-Slowik, M.; Karczewicz, D.; Safranow, K. The potential role of oxidative stress in the pathogenesis of the age-related macular degeneration (AMD). (in Polish). Postepy Hig. Med. Dosw. Online 2007, 61, 28–37. [Google Scholar]
- Nowak, M.; Gnitecki, W.; Jurowski, P. The role of retinal oxygen metabolism in origin of age-related macular degeneration (AMD). (in Polish). Klin. Oczna 2005, 107, 715–718. [Google Scholar]
- King, A.; Gottlieb, E.; Brooks, D.G.; Murphy, M.P.; Dunaief, J.L. Mitochondria-derived reactive oxygen species mediate blue light-induced death of retinal pigment epithelial cells. Photochem. Photobiol. 2004, 79, 470–475. [Google Scholar]
- Xu, K.P.; Yu, F.S. Cross talk between c-Met and epidermal growth factor receptor during retinal pigment epithelial wound healing. Investig. Ophthal. Mol. Vis. Sci. 2007, 48, 2242–2248. [Google Scholar]
- Yan, F.; Hui, Y.N.; Li, Y.J.; Guo, C.M.; Meng, H. Epidermal growth factor receptor in cultured human retinal pigment epithelial cells. Ophthalmologica 2007, 221, 244–250. [Google Scholar]
- Zhang, L.; Wang, F.; Jiang, Y.; Xu, S.; Lu, F.; Wang, W.; Sun, X. Migration of retinal pigment epithelial cells is EGFR/PI3K/AKT dependent. Front. BioSci. Schol. Ed. 2013, 5, 661–671. [Google Scholar]
- Herbst, R.S. Review of epidermal growth factor receptor biology. Int. J. Radiat. Oncol. Biol. Phys. 2004, 59, 21–26. [Google Scholar]
- Cao, C.; Huang, X.; Han, Y.; Wan, Y.; Birnbaumer, L.; Feng, G.S.; Marshall, J.; Jiang, M.; Chu, W.M. Galpha(i1) and Galpha(i3) are required for epidermal growth factor-mediated activation of the Akt-mTORC1 pathway. Sci. Signal 2009, 2. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, L.-B.; Chen, C.-M.; Zhong, H.; Zhu, L.-J. Squamosamide Derivative FLZ Protects Retinal Pigment Epithelium Cells from Oxidative Stress through Activation of Epidermal Growth Factor Receptor (EGFR)-AKT Signaling. Int. J. Mol. Sci. 2014, 15, 18762-18775. https://doi.org/10.3390/ijms151018762
Cheng L-B, Chen C-M, Zhong H, Zhu L-J. Squamosamide Derivative FLZ Protects Retinal Pigment Epithelium Cells from Oxidative Stress through Activation of Epidermal Growth Factor Receptor (EGFR)-AKT Signaling. International Journal of Molecular Sciences. 2014; 15(10):18762-18775. https://doi.org/10.3390/ijms151018762
Chicago/Turabian StyleCheng, Li-Bo, Chun-Ming Chen, Hong Zhong, and Li-Juan Zhu. 2014. "Squamosamide Derivative FLZ Protects Retinal Pigment Epithelium Cells from Oxidative Stress through Activation of Epidermal Growth Factor Receptor (EGFR)-AKT Signaling" International Journal of Molecular Sciences 15, no. 10: 18762-18775. https://doi.org/10.3390/ijms151018762