Cartilage Turnover Reflected by Metabolic Processing of Type II Collagen: A Novel Marker of Anabolic Function in Chondrocytes

Abstract
:1. Introduction
2. Results
2.1. Specificity of the Monoclonal Antibody

2.2. Technical Performance of the Pro-C2 ELISA

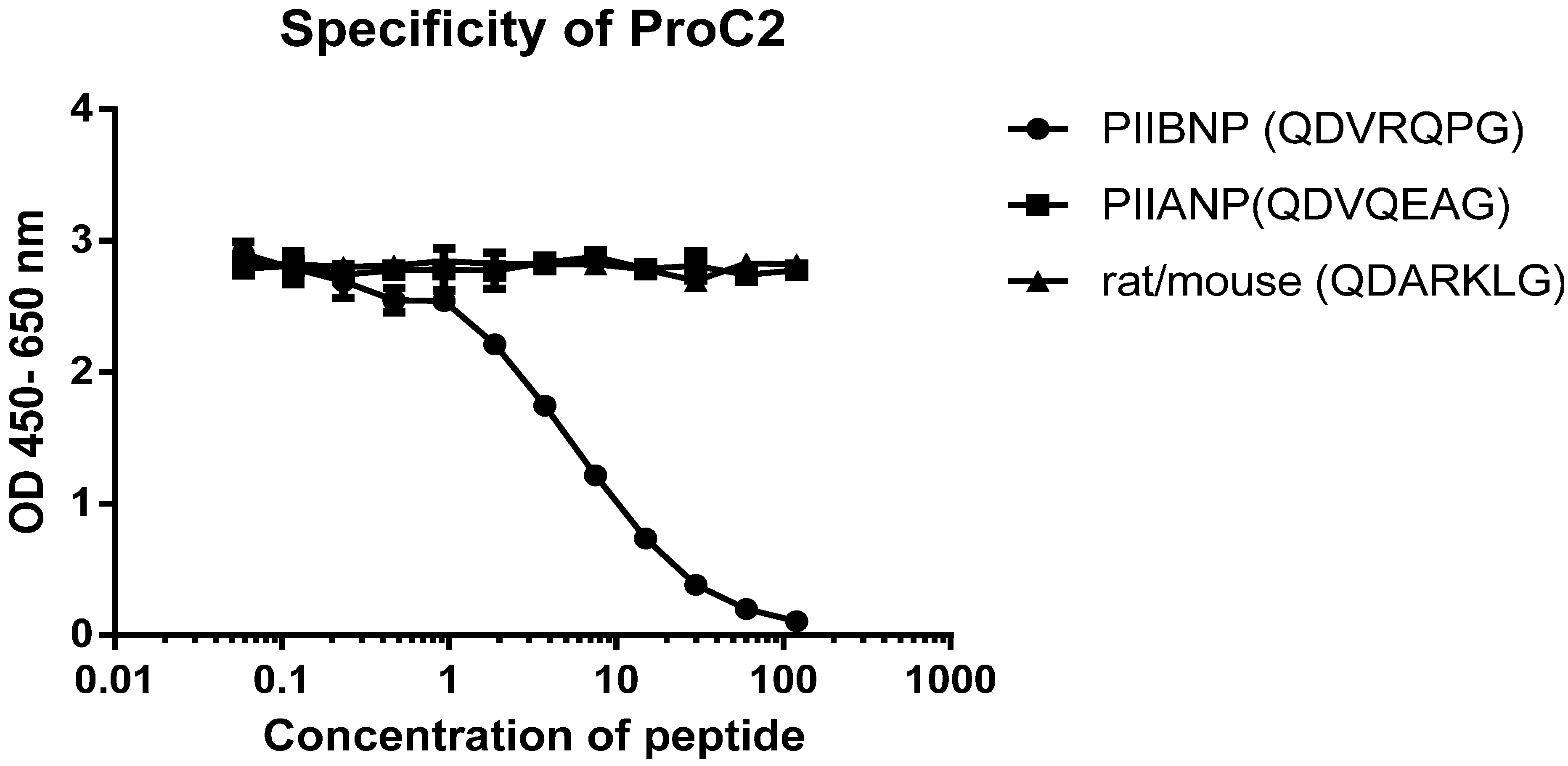{kind=link}
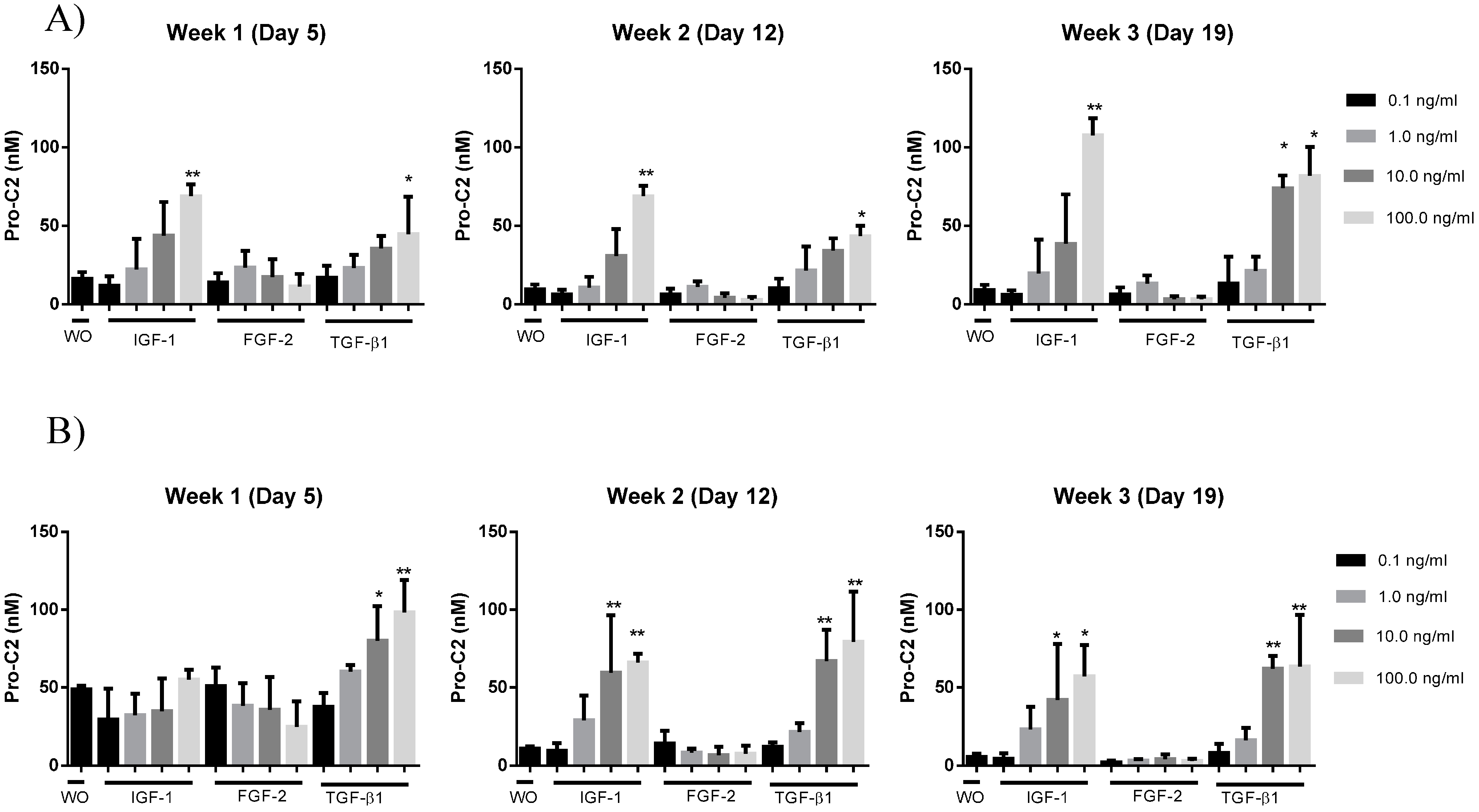{kind=link}
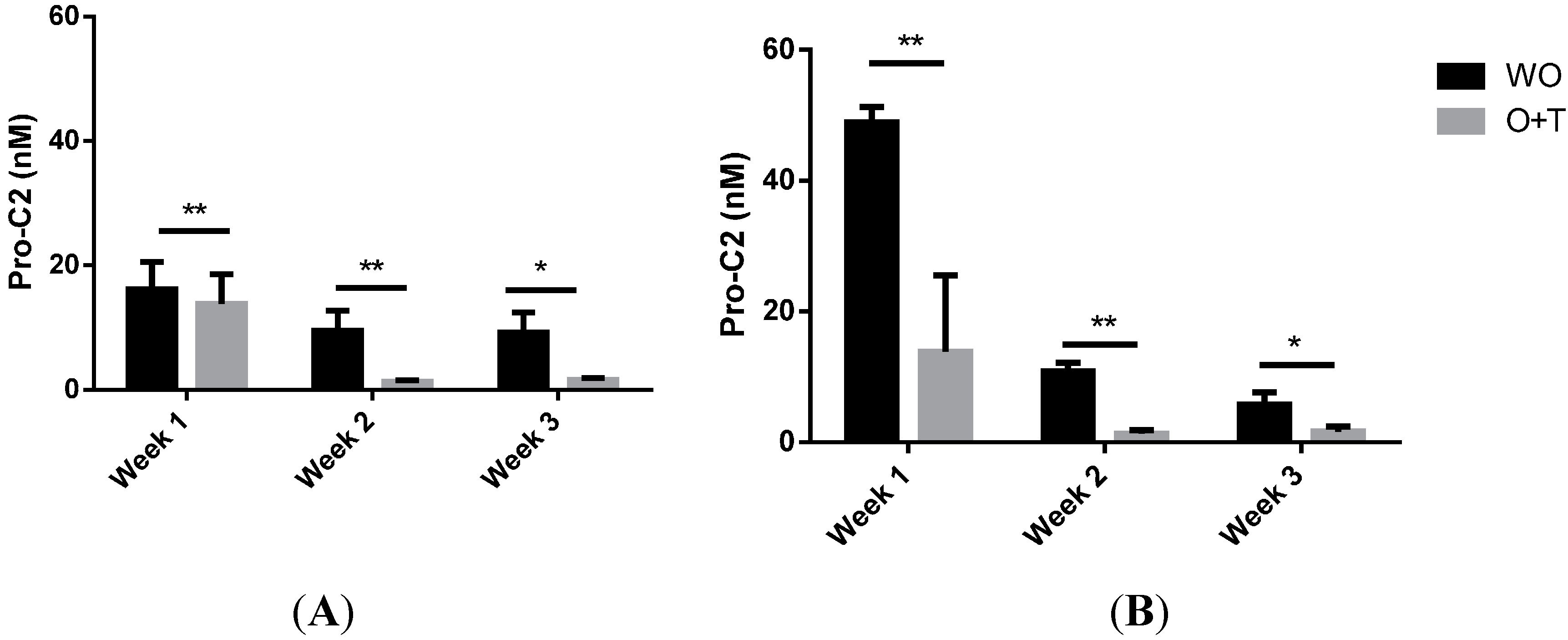{kind=link}
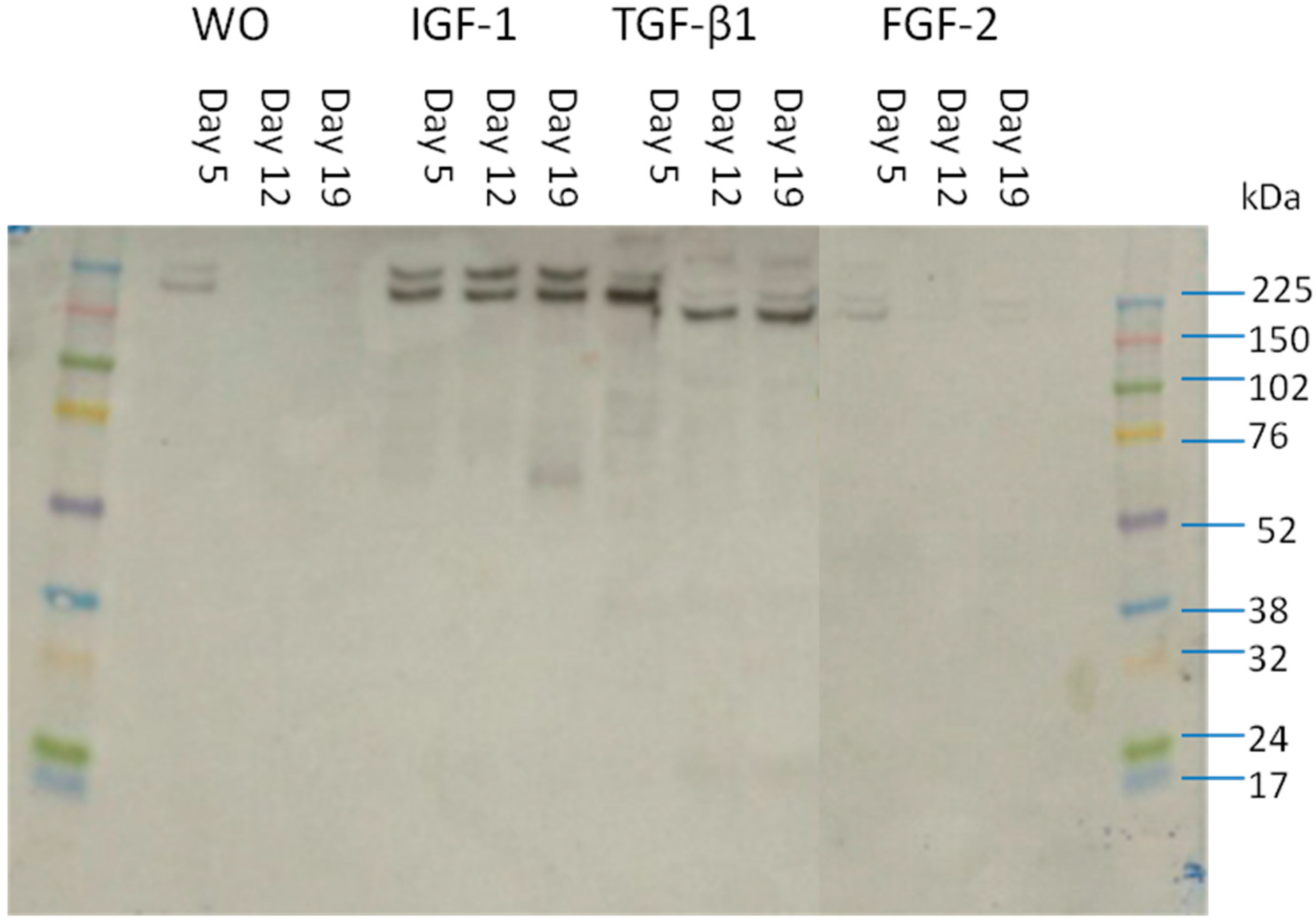{kind=link}
{kind=link}
{kind=link}
| Assay Specifications | Pro-C2 |
|---|---|
| Slope of standard curve | 1.03 |
| IC50, nM | 7.82 |
| Intra-assay CV% | 10.8 |
| Inter-assay CV% | 9.2 |
| Lower limit of detection, nM | 0.96 |
| Quantifiable range, nM | 2.5–19 |
2.3. PIIBNP Levels Measured in Serum of Humans and Other Species
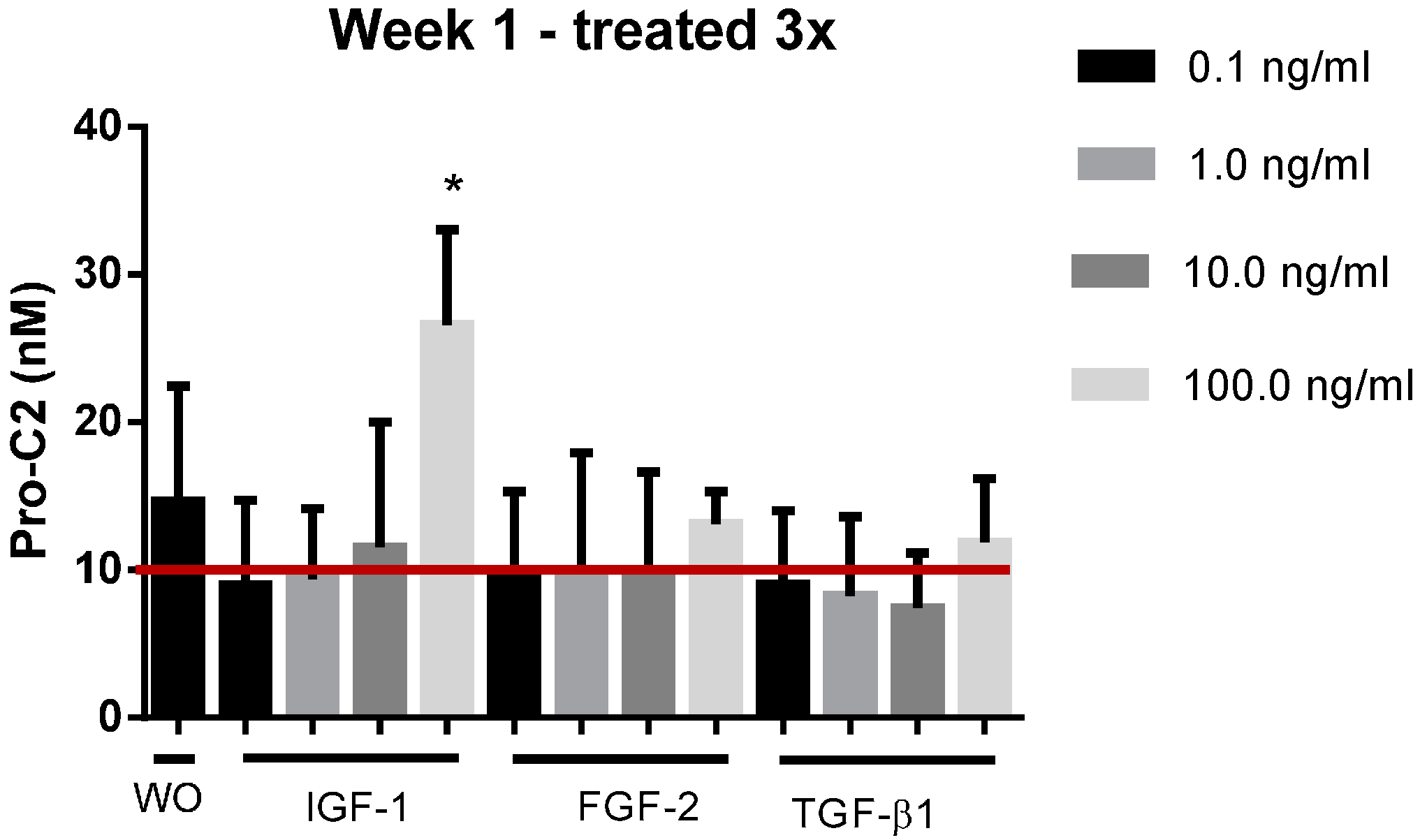
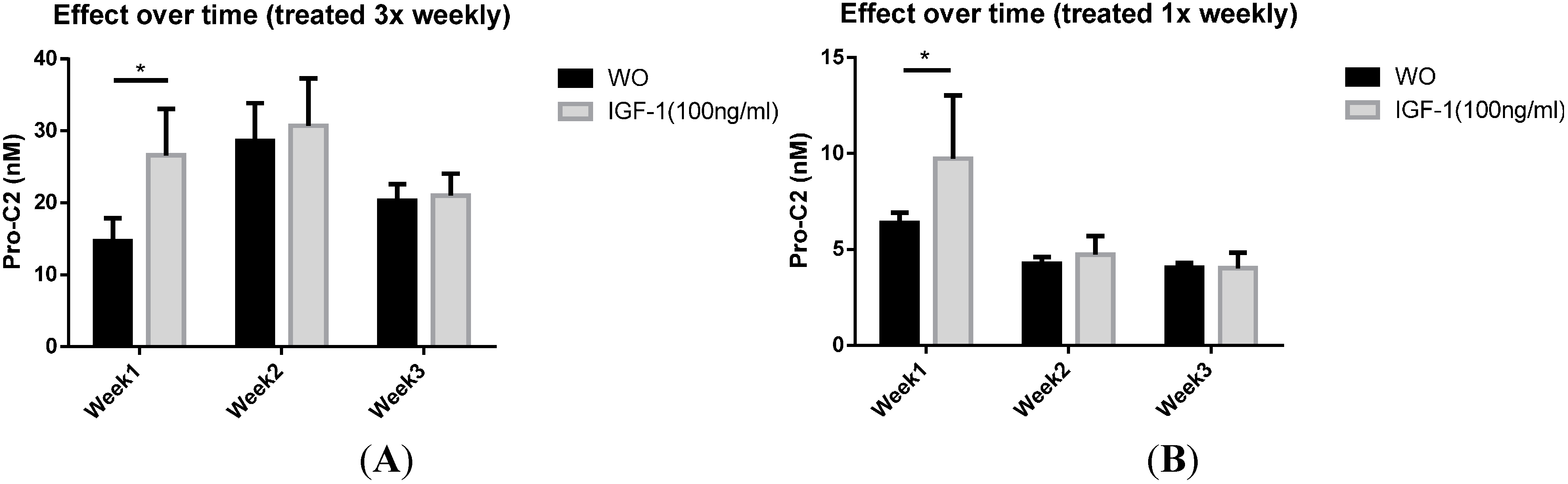
2.4. PIIBNP Release in Bovine Cartilage Explants


2.5. Size of the Detected Fragments in Supernatant

2.6. PIIBNP Measured in Human Cartilage Explants


3. Discussion
| Species | PIIBNP (nM) |
|---|---|
| Human (adult) serum | 1.5 |
| Amniotic fluid | 175 |
| Fetal bovine serum | 875 |
| Bovine serum | 9 |
| Fetal horse serum | 415 |
| Chicken serum | 0.3 |
| Rabbit serum | 0.2 |
4. Experimental Section
4.1. Immunogen Identification, Alignment and Selection
| Species | Sequence | NCBI Reference |
|---|---|---|
| Human Col2α1, isoform 2 (target) | QDVRQPG | NP_149162.2 |
| Human Col2α1, isoform 1 | QDV  G G | NP_001835.3 |
| Bovine Col2α1, isoform 1 | QDV  G G | NP_001001135.2 |
| Bovine Col2α1, isoform 2 | QDVRQPG | NP_001106695.1 |
| Mouse Col2α1, isoform 1 | QD  G G | NP_112440.2 |
| Mouse Col2α1, isoform 2 | QD  G G | NP_001106987.2 |
| Rat Col2α1 | QD G | NP_037061.1 |
| Rabbit Col2α1 | QDV G | NP_001182600 |
| Horse Col2α1 | QDVRQPG | NP_001075233.1 |
| Chicken Col2α1 |  RQPG RQPG | NP_989757.1 |
4.2. Generation of the Monoclonal Antibody
4.3. Assay Development
4.4. Bovine and Human Cartilage Explants Culture
4.5. Western Blotting of BEX Supernatant
4.6. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Abbreviations
| AGG-C1 | A biomarker of an aggrecan specific neoepitope [19] |
| AGNX-II | A biomarker of an aggrecan specific neoepitope [18] |
| BEX | Bovine cartilage explants |
| Bio | Biotinylated |
| C2C | A biomarker of a collagen type II specific neoepitope |
| CIIM | A biomarker of a collagen type II specific neoepitope [13] |
| CIINE | A biomarker of a collagen type II specific neoepitope [16] |
| COMP | A biomarker for cartilage oligomeric matrix protein [20] |
| CTX-II | A biomarker of a type II collagen C-terminal telopeptide [14] |
| CV | Coefficient of variation |
| DJD | Degenerative joint disease |
| DMARD | Disease-modifying anti-rheumatic drugs |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DMOARD | Disease modifying osteoarthritis drug |
| ECL | Enhanced chemiluminescence |
| ECM | Extra cellular matrix |
| FBS | Fetal bovine serum |
| FGF | Fibroblastic growth factor |
| HAT | Hyposanthine/aminoptrein/thymidine |
| HEX | Human cartilage explants |
| IGF | Insulin like growth factor |
| LLOD | Lower limit of detection |
| LLOQ | lower limit of quantification |
| MRI | Magnetic resonance imaging |
| OA | Osteoarthritis |
| PBS | Phosphate buffered saline |
| PIIANP | Isotype I of type II procollagen |
| PIIBNP | Isotype II of type II procollagen |
| PIICP | C-terminal propeptide of type II collagen |
| PIINP | N-terminal propeptide of type II collagen |
| RA | Rheumatoid arthritis |
| SD | Standard deviation |
| TBS | Tris-buffered Saline |
| TGF | Transforming growth factor |
| TIINE | A biomarker of a type II collagen inter-helical fragment [15] |
| TMB | Tetramethyl benzinidine |
| ULOQ | Upper limit of quantification |
| WO | without treatment |
Conflicts of Interest
References
- Dieppe, P.A.; Lohmander, L.S. Pathogenesis and management of pain in osteoarthritis. Lancet 2005, 365, 965–973. [Google Scholar] [CrossRef]
- Tchetina, E.V. Developmental mechanisms in articular cartilage degradation in osteoarthritis. Arthritis 2011, 2011, 683970. [Google Scholar] [CrossRef]
- Tchetina, E.V.; Squires, G.; Poole, A.R. Increased type II collagen degradation and very early focal cartilage degeneration is associated with upregulation of chondrocyte differentiation related genes in early human articular cartilage lesions. J. Rheumatol. 2005, 32, 876–886. [Google Scholar]
- Davies, P.S.; Graham, S.M.; MacFarlane, R.J.; Leonidou, A.; Mantalaris, A.; Tsiridis, E. Disease-modifying osteoarthritis drugs: In vitro and in vivo data on the development of DMOADs under investigation. Expert Opin. Investig. Drugs 2013, 22, 423–441. [Google Scholar] [CrossRef]
- Pelletier, J.P.; Cooper, C.; Peterfy, C.; Reginster, J.Y.; Brandi, M.L.; Bruyere, O.; Chapurlat, R.; Cicuttini, F.; Conaghan, P.G.; Doherty, M.; et al. What is the predictive value of MRI for the occurrence of knee replacement surgery in knee osteoarthritis? Ann. Rheum. Dis. 2013, 72, 1594–1604. [Google Scholar] [CrossRef]
- Abadie, E.; Ethgen, D.; Avouac, B.; Bouvenot, G.; Branco, J.; Bruyere, O.; Calvo, G.; Devogelaer, J.P.; Dreiser, R.L.; Herrero-Beaumont, G.; et al. Recommendations for the use of new methods to assess the efficacy of disease-modifying drugs in the treatment of osteoarthritis. Osteoarthr. Cartil. 2004, 12, 263–268. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, S.; Huang, J.; Guo, W.; Chen, J.; Zhang, L.; Zhao, B.; Peng, J.; Wang, A.; Wang, Y.; et al. The ECM-Cell interaction of cartilage extracellular matrix on chondrocytes. Biomed. Res. Int. 2014, 2014, 648459. [Google Scholar]
- Martel-Pelletier, J.; Boileau, C.; Pelletier, J.P.; Roughley, P.J. Cartilage in normal and osteoarthritis conditions. Best Pract. Res. Clin. Rheumatol. 2008, 22, 351–384. [Google Scholar] [CrossRef]
- Van der, R.M.; Garrone, R. Collagen family of proteins. FASEB J. 1991, 5, 2814–2823. [Google Scholar]
- Aigner, T.; Zhu, Y.; Chansky, H.H.; Matsen, F.A., III; Maloney, W.J.; Sandell, L.J. Reexpression of type IIA procollagen by adult articular chondrocytes in osteoarthritic cartilage. Arthritis Rheumatol. 1999, 42, 1443–1450. [Google Scholar] [CrossRef]
- Rousseau, J.C.; Sandell, L.J.; Delmas, P.D.; Garnero, P. Development and clinical application in arthritis of a new immunoassay for serum type IIA procollagen NH2 propeptide. Methods Mol. Med. 2004, 101, 25–37. [Google Scholar]
- Sandell, L.J.; Morris, N.; Robbins, J.R.; Goldring, M.B. Alternatively spliced type II procollagen mRNAs define distinct populations of cells during vertebral development: Differential expression of the amino-propeptide. J. Cell Biol. 1991, 114, 1307–1319. [Google Scholar] [CrossRef]
- Bay-Jensen, A.C.; Liu, Q.; Byrjalsen, I.; Li, Y.; Wang, J.; Pedersen, C.; Leeming, D.J.; Dam, E.B.; Zheng, Q.; Qvist, P.; et al. Enzyme-linked immunosorbent assay (ELISAs) for metalloproteinase derived type II collagen neoepitope, CIIM—Increased serum CIIM in subjects with severe radiographic osteoarthritis. Clin. Biochem. 2011, 44, 423–429. [Google Scholar] [CrossRef]
- Christgau, S.; Garnero, P.; Fledelius, C.; Moniz, C.; Ensig, M.; Gineyts, E.; Rosenquist, C.; Qvist, P. Collagen type II C-telopeptide fragments as an index of cartilage degradation. Bone 2001, 29, 209–215. [Google Scholar] [CrossRef]
- Downs, J.T.; Lane, C.L.; Nestor, N.B.; McLellan, T.J.; Kelly, M.A.; Karam, G.A.; Mezes, P.S.; Pelletier, J.P.; Otterness, I.G. Analysis of collagenase-cleavage of type II collagen using a neoepitope ELISA. J. Immunol. Methods 2001, 247, 25–34. [Google Scholar] [CrossRef]
- Takahashi, T.; Naito, S.; Onoda, J.; Yamauchi, A.; Nakamura, E.; Kishino, J.; Kawai, T.; Matsukawa, S.; Toyosaki-Maeda, T.; Tanimura, M.; et al. Development of a novel immunoassay for the measurement of type II collagen neoepitope generated by collagenase cleavage. Clin. Chim. Acta 2012, 413, 1591–1599. [Google Scholar] [CrossRef]
- Poole, A.R.; Ionescu, M.; Fitzcharles, M.A.; Billinghurst, R.C. The assessment of cartilage degradation in vivo: Development of an immunoassay for the measurement in body fluids of type II collagen cleaved by collagenases. J. Immunol. Methods 2004, 294, 145–153. [Google Scholar] [CrossRef]
- Wang, B.; Chen, P.; Jensen, A.C.; Karsdal, M.A.; Madsen, S.H.; Sondergaard, B.C.; Zheng, Q.; Qvist, P. Suppression of MMP activity in bovine cartilage explants cultures has little if any effect on the release of aggrecanase-derived aggrecan fragments. BMC Res. Notes 2009, 2, 259. [Google Scholar] [CrossRef] [Green Version]
- Chockalingam, P.S.; Zeng, W.; Morris, E.A.; Flannery, C.R. Release of hyaluronan and hyaladherins (aggrecan G1 domain and link proteins) from articular cartilage exposed to ADAMTS-4 (aggrecanase 1) or ADAMTS-5 (aggrecanase 2). Arthritis Rheumatol. 2004, 50, 2839–2848. [Google Scholar] [CrossRef]
- Saxne, T.; Heinegard, D. Cartilage oligomeric matrix protein: A novel marker of cartilage turnover detectable in synovial fluid and blood. Br. J. Rheumatol. 1992, 31, 583–591. [Google Scholar] [CrossRef]
- Olsen, A.K.; Sondergaard, B.C.; Byrjalsen, I.; Tanko, L.B.; Christiansen, C.; Muller, A.; Hein, G.E.; Karsdal, M.A.; Qvist, P. Anabolic and catabolic function of chondrocyte ex vivo is reflected by the metabolic processing of type II collagen. Osteoarthr. Cartil. 2007, 15, 335–342. [Google Scholar] [CrossRef]
- Madsen, S.H.; Sondergaard, B.C.; Bay-Jensen, A.C.; Karsdal, M.A. Cartilage formation measured by a novel PIINP assay suggests that IGF-I does not stimulate but maintains cartilage formation ex vivo. Scand. J. Rheumatol. 2009, 38, 222–226. [Google Scholar] [CrossRef]
- Nemirovskiy, O.V.; Sunyer, T.; Aggarwal, P.; Abrams, M.; Hellio Le Graverand, M.P.; Mathews, W.R. Discovery and development of the N-terminal procollagen type II (NPII) biomarker: A tool for measuring collagen type II synthesis. Osteoarthr. Cartil. 2008, 16, 1494–1500. [Google Scholar] [CrossRef]
- Hinek, A.; Reiner, A.; Poole, A.R. The calcification of cartilage matrix in chondrocyte culture: Studies of the C-propeptide of type II collagen (chondrocalcin). J. Cell Biol. 1987, 104, 1435–1441. [Google Scholar] [CrossRef]
- Sharif, M.; Kirwan, J.; Charni, N.; Sandell, L.J.; Whittles, C.; Garnero, P. A 5-yr longitudinal study of type IIA collagen synthesis and total type II collagen degradation in patients with knee osteoarthritis—Association with disease progression. Rheumatology 2007, 46, 938–943. [Google Scholar] [CrossRef]
- Gelse, K.; Poschl, E.; Aigner, T. Collagens—Structure, function, and biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef] [Green Version]
- Garvican, E.R.; Vaughan-Thomas, A.; Innes, J.F.; Clegg, P.D. Biomarkers of cartilage turnover. Part 1: Markers of collagen degradation and synthesis. Vet. J. 2010, 185, 36–42. [Google Scholar] [CrossRef]
- Munk, H.L.; Svendsen, A.J.; Hjelmborg, J.V.; Sorensen, G.L.; Kyvik, K.O.; Junker, P. Heritability assessment of cartilage metabolism. A twin study on circulating procollagen II A N-terminal propeptide (PIIANP). Osteoarthr. Cartil. 2014, 22, 1142–1147. [Google Scholar] [CrossRef]
- Hall, B.K.; Miyake, T. Divide, accumulate, differentiate: Cell condensation in skeletal development revisited. Int. J. Dev. Biol. 1995, 39, 881–893. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gudmann, N.S.; Wang, J.; Hoielt, S.; Chen, P.; Siebuhr, A.S.; He, Y.; Christiansen, T.G.; Karsdal, M.A.; Bay-Jensen, A.C. Cartilage Turnover Reflected by Metabolic Processing of Type II Collagen: A Novel Marker of Anabolic Function in Chondrocytes. Int. J. Mol. Sci. 2014, 15, 18789-18803. https://doi.org/10.3390/ijms151018789
Gudmann NS, Wang J, Hoielt S, Chen P, Siebuhr AS, He Y, Christiansen TG, Karsdal MA, Bay-Jensen AC. Cartilage Turnover Reflected by Metabolic Processing of Type II Collagen: A Novel Marker of Anabolic Function in Chondrocytes. International Journal of Molecular Sciences. 2014; 15(10):18789-18803. https://doi.org/10.3390/ijms151018789
Chicago/Turabian StyleGudmann, Natasja Stæhr, Jianxia Wang, Sabine Hoielt, Pingping Chen, Anne Sofie Siebuhr, Yi He, Thorbjørn G. Christiansen, Morten Asser Karsdal, and Anne Christine Bay-Jensen. 2014. "Cartilage Turnover Reflected by Metabolic Processing of Type II Collagen: A Novel Marker of Anabolic Function in Chondrocytes" International Journal of Molecular Sciences 15, no. 10: 18789-18803. https://doi.org/10.3390/ijms151018789