MicroRNA-130b Promotes Cell Aggressiveness by Inhibiting Peroxisome Proliferator-Activated Receptor Gamma in Human Hepatocellular Carcinoma

Abstract
:1. Introduction
2. Results
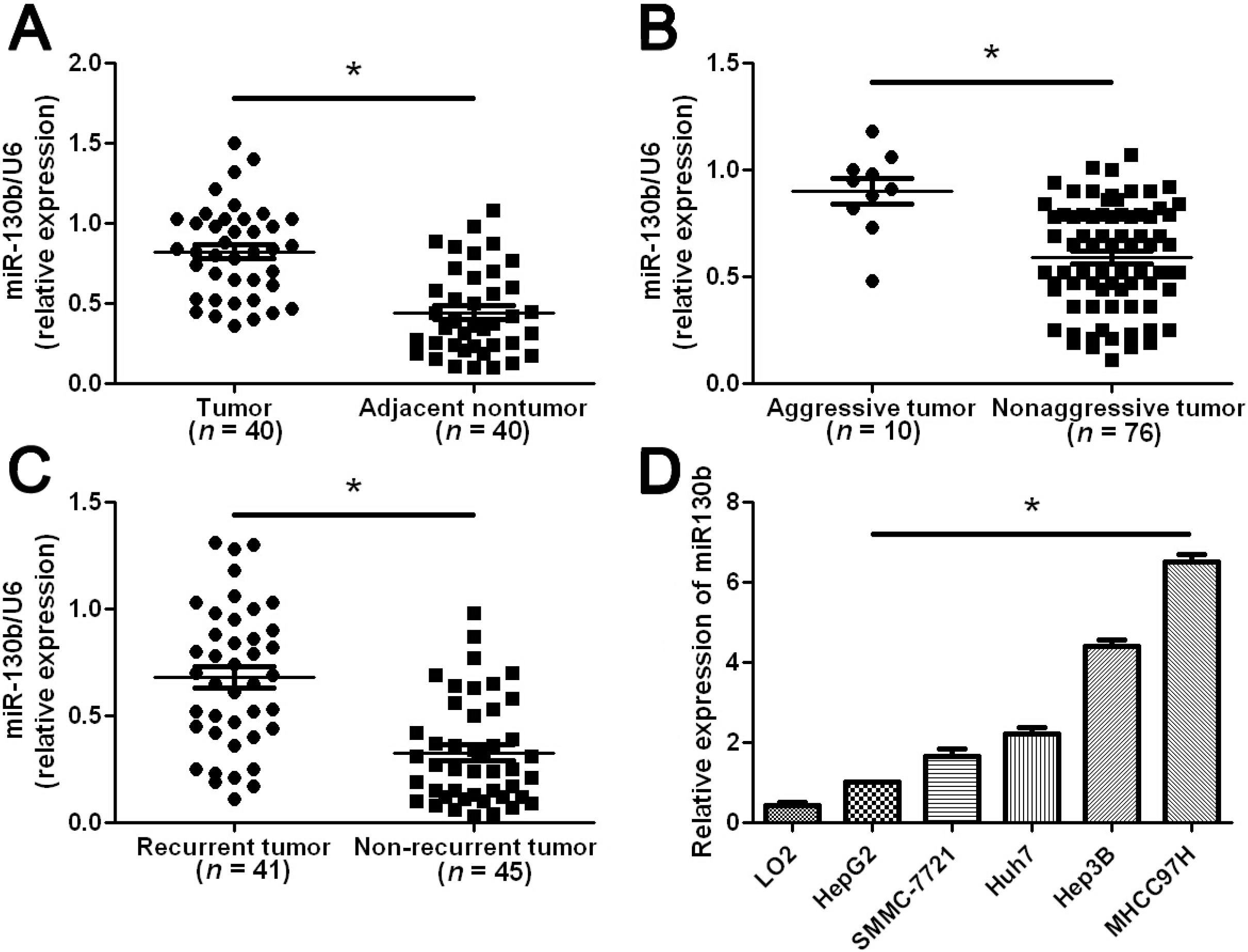
2.1. Elevated miR-130b Expression Confers Metastasis and Recurrence of HCC (Hepatocellular Carcinoma)

2.2. Clinical Significance of miR-130b Expression in HCC Specimens

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Total No. of Patients, n = 86 | No. of Patients | p | ||
|---|---|---|---|---|---|
| miR-130b high | miR-130b low | ||||
| Age (year) | <50 | 36 | 19 | 17 | 0.323 |
| ≥50 | 50 | 21 | 29 | ||
| Sex | Male | 65 | 28 | 37 | 0.261 |
| Female | 21 | 12 | 9 | ||
| HBV | Absent | 28 | 9 | 19 | 0.063 |
| Present | 58 | 31 | 27 | ||
| Serum AFP level (ng/mL) | <400 | 32 | 14 | 18 | 0.693 |
| ≥400 | 54 | 26 | 28 | ||
| Tumor size (cm) | <5 | 32 | 16 | 16 | 0.618 |
| ≥5 | 54 | 24 | 30 | ||
| No. of tumor nodules | 1 | 68 | 28 | 40 | 0.054 |
| ≥2 | 18 | 12 | 6 | ||
| Cirrhosis | Absent | 36 | 13 | 23 | 0.101 |
| Present | 50 | 27 | 23 | ||
| Venous infiltration | Absent | 76 | 31 | 45 | 0.009 * |
| Present | 10 | 9 | 1 | ||
| Edmondson-Steiner grading | I + II | 65 | 25 | 40 | 0.008 * |
| III + IV | 21 | 15 | 6 | ||
| TNM tumor stage | I + II | 66 | 22 | 44 | <0.001 * |
| III + IV | 20 | 18 | 2 | ||
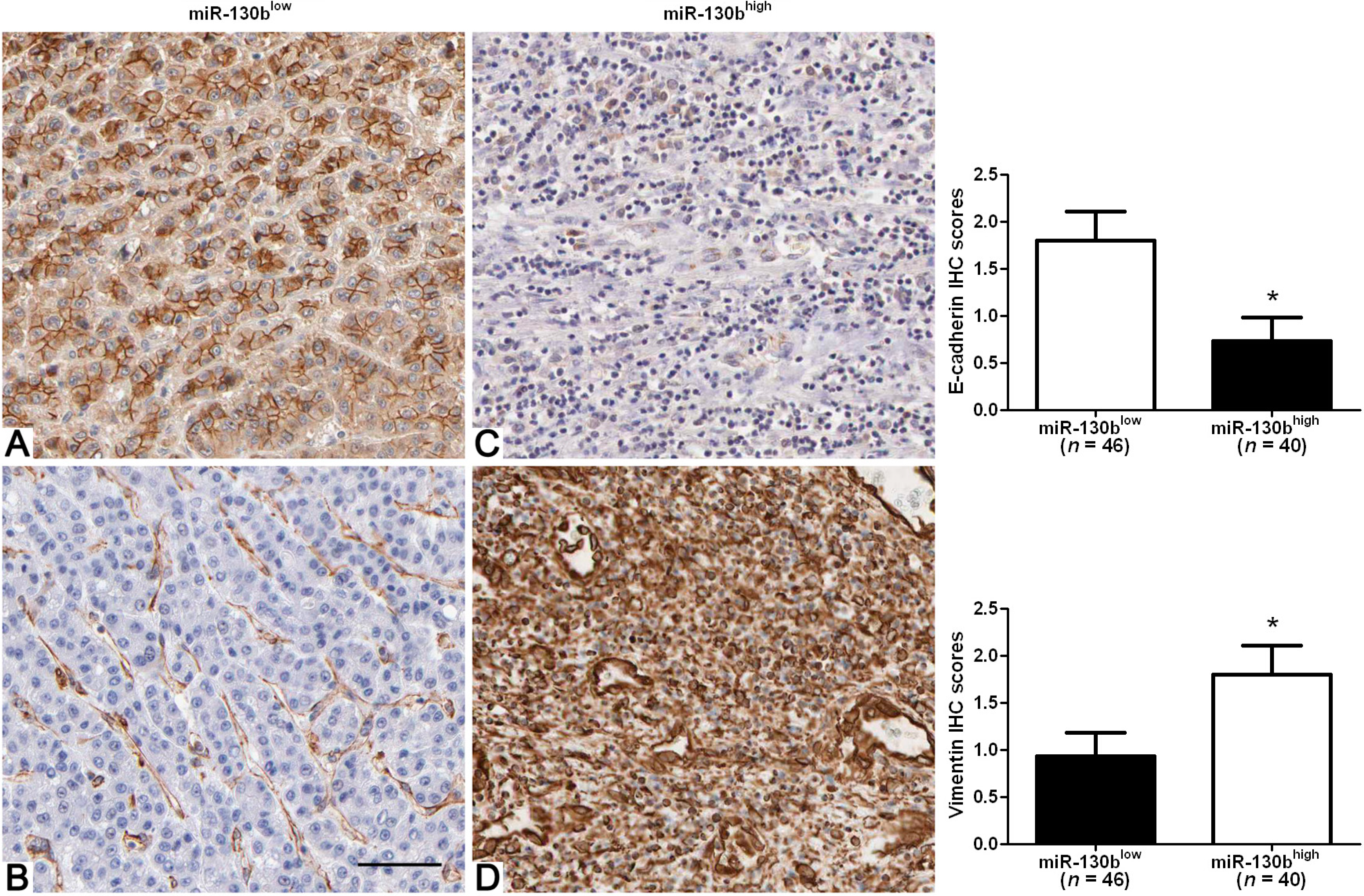
2.3. High-Expression of miR-130b Correlates with Mesenchymal Phenotype of HCC

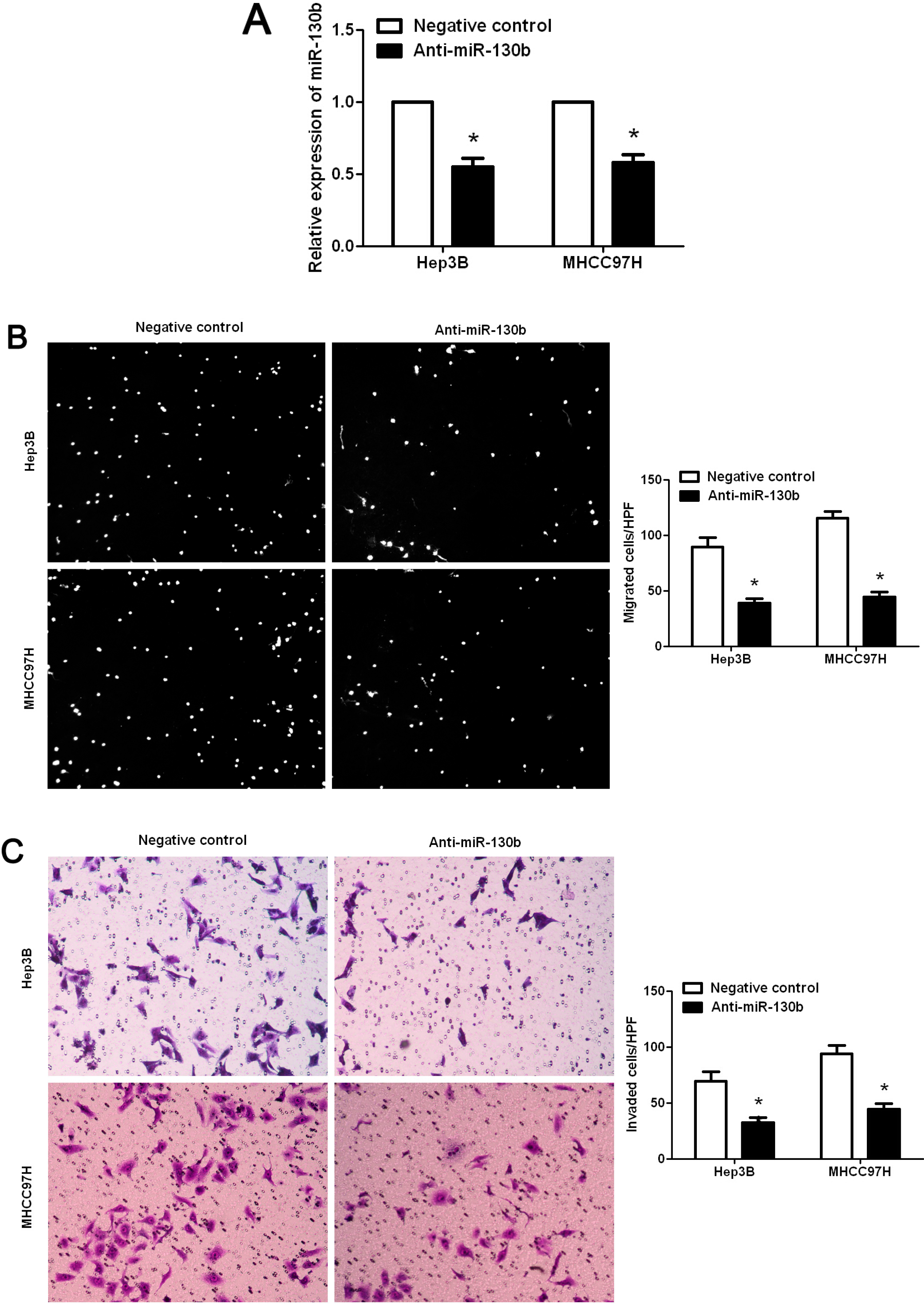
2.4. Promoting Effect of miR-130b on HCC Cell Migration and Invasion

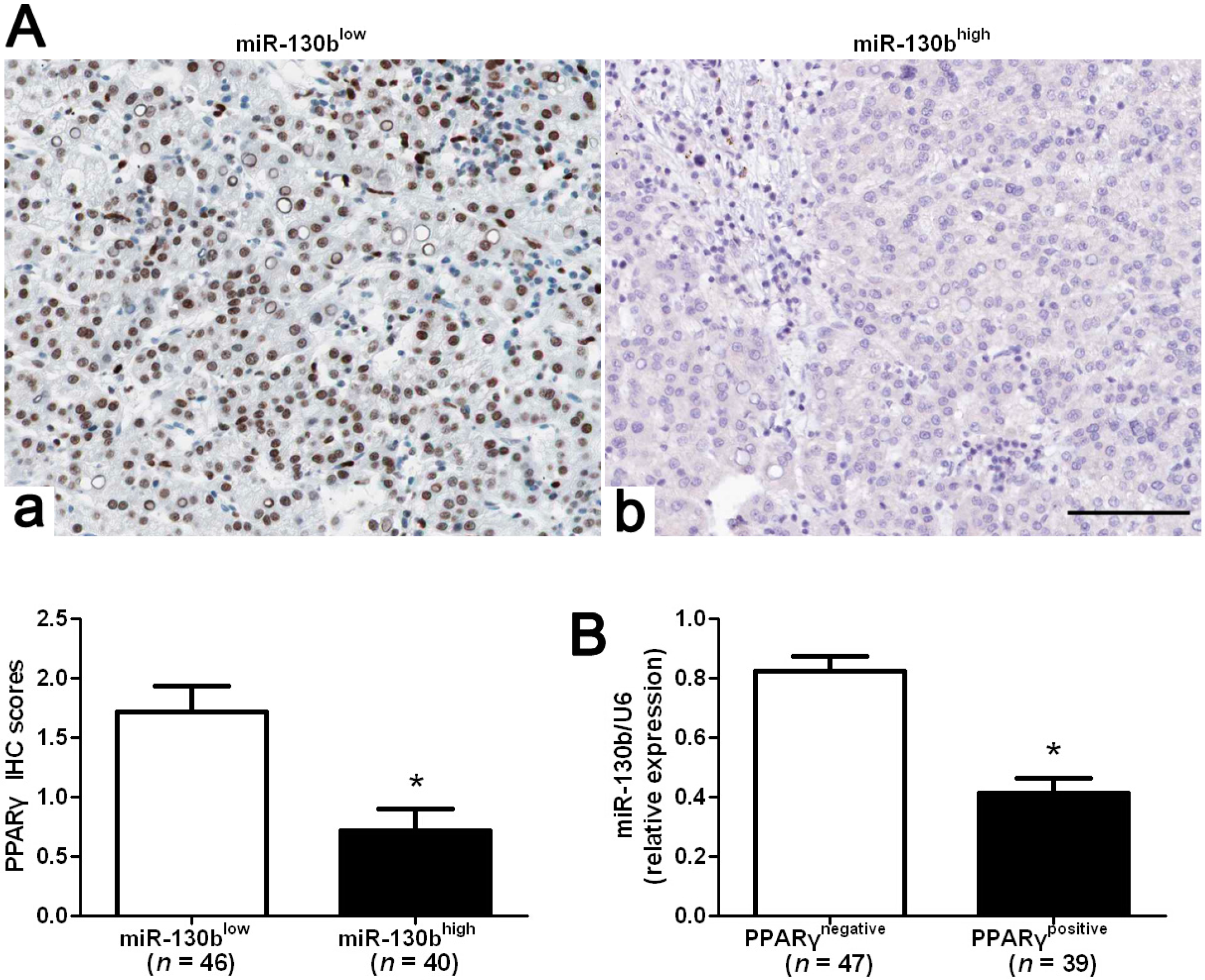
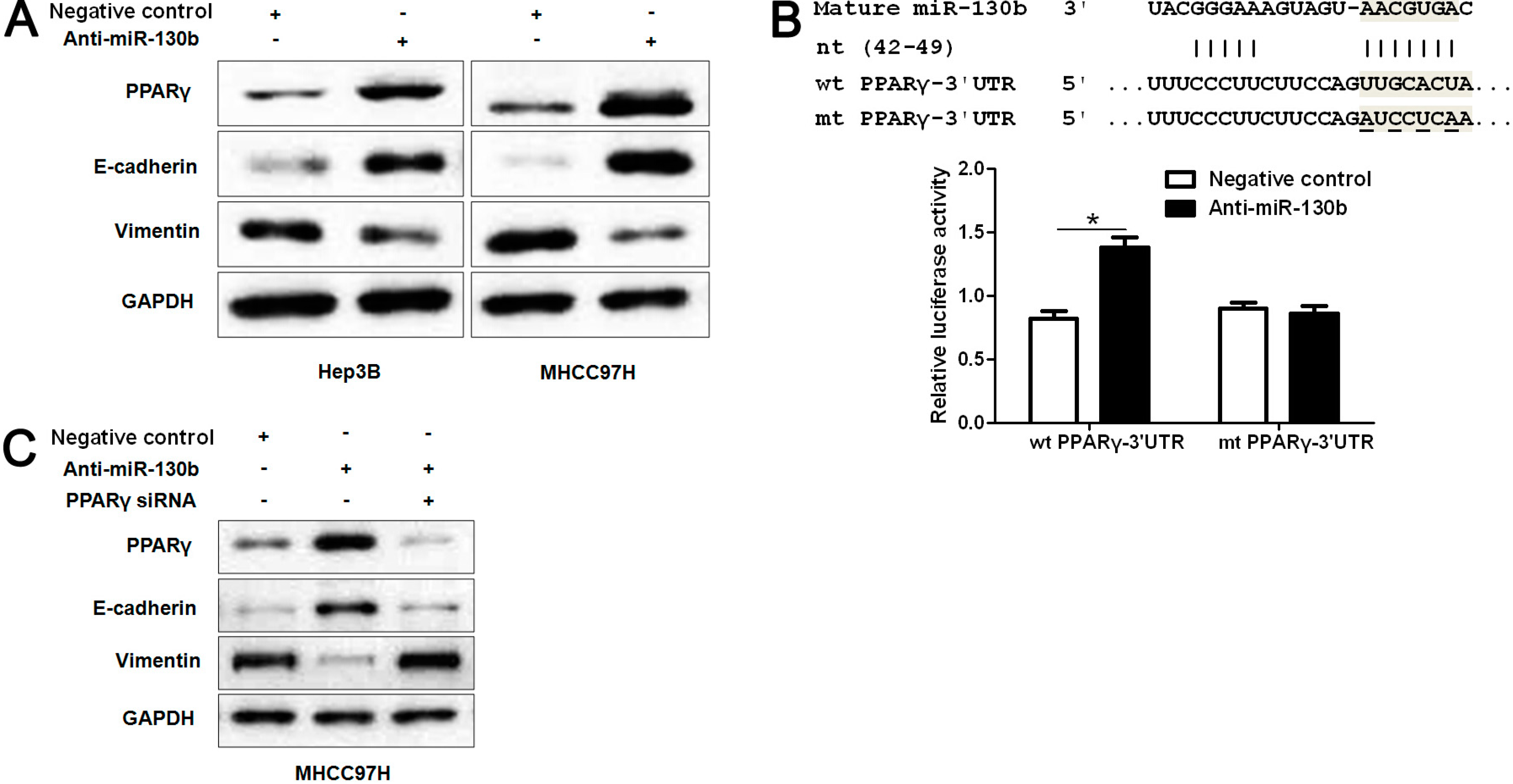
2.5. MiR-130b Promotes EMT (Epithelial-Mesenchymal Transition) by Inhibiting PPARγ in HCC


3. Discussion
4. Experimental Section
4.1. Ethical Review
4.2. Clinical Samples
4.3. Immunohistochemical Staining
4.4. Cell Lines and Transfection
4.5. Western Blot
4.6. Boyden Chamber and Transwell Assays
4.7. Real Time Quantitative Reverse Transcription-PCR (qRT-PCR)
4.8. Luciferase Reporter Assay
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Garzon, R.; Calin, G.A.; Croce, C.M. MicroRNAs in Cancer. Annu. Rev. Med. 2009, 60, 167–179. [Google Scholar]
- Rosa, A.; Brivanlou, A.H. MicroRNAs in early vertebrate development. Cell Cycle 2009, 8, 3513–3520. [Google Scholar]
- Harfe, B.D. MicroRNAs in vertebrate development. Curr. Opin. Genet. Dev. 2005, 15, 410–415. [Google Scholar]
- Vasudevan, S.; Tong, Y.; Steitz, J.A. Switching from repression to activation: microRNAs can up-regulate translation. Science 2007, 318, 1931–1934. [Google Scholar]
- Croce, C.M.; Calin, G.A. miRNAs, cancer, and stem cell division. Cell 2005, 122, 6–7. [Google Scholar]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar]
- Jansson, M.D.; Lund, A.H. MicroRNA and cancer. Mol. Oncol. 2012, 6, 590–610. [Google Scholar]
- Wang, W.; Peng, B.; Wang, D.; Ma, X.; Jiang, D.; Zhao, J.; Yu, L. Human tumor microRNA signatures derived from large-scale oligonucleotide microarray datasets. Int. J. Cancer 2011, 129, 1624–1634. [Google Scholar]
- Yip, L.; Kelly, L.; Shuai, Y.; Armstrong, M.J.; Nikiforov, Y.E.; Carty, S.E.; Nikiforova, M.N. MicroRNA signature distinguishes the degree of aggressiveness of papillary thyroid carcinoma. Ann. Surg. Oncol. 2011, 18, 2035–2041. [Google Scholar]
- Dong, P.; Karaayvaz, M.; Jia, N.; Kaneuchi, M.; Hamada, J.; Watari, H.; Sudo, S.; Ju, J.; Sakuragi, N. Mutant p53 gain-of-function induces epithelial-mesenchymal transition through modulation of the miR-130b-ZEB1 axis. Oncogene 2013, 32, 3286–3295. [Google Scholar]
- Leone, V.; Langella, C.; D’Angelo, D.; Mussnich, P.; Wierinckx, A.; Terracciano, L.; Raverot, G.; Lachuer, J.; Rotondi, S.; Jaffrain-Rea, M.L.; et al. Mir-23b and miR-130b expression is downregulated in pituitary adenomas. Mol. Cell Endocrinol. 2014, 390, 1–7. [Google Scholar]
- Zhao, G.; Zhang, J.G.; Shi, Y.; Qin, Q.; Liu, Y.; Wang, B.; Tian, K.; Deng, S.C.; Li, X.; Zhu, S.; et al. MiR-130b is a prognostic marker and inhibits cell proliferation and invasion in pancreatic cancer through targeting STAT3. PLoS One 2013, 8, e73803. [Google Scholar]
- Sand, M.; Skrygan, M.; Sand, D.; Georgas, D.; Gambichler, T.; Hahn, S.A.; Altmeyer, P.; Bechara, F.G. Comparative microarray analysis of microRNA expression profiles in primary cutaneous malignant melanoma, cutaneous malignant melanoma metastases, and benign melanocytic nevi. Cell Tissue Res. 2013, 351, 85–98. [Google Scholar]
- Lai, K.W.; Koh, K.X.; Loh, M.; Tada, K.; Subramaniam, M.M.; Lim, X.Y.; Vaithilingam, A.; Salto-Tellez, M.; Iacopetta, B.; Ito, Y.; et al. MicroRNA-130b regulates the tumour suppressor RUNX3 in gastric cancer. Eur. J. Cancer 2010, 46, 1456–1463. [Google Scholar]
- Kim, B.H.; Hong, S.W.; Kim, A.; Choi, S.H.; Yoon, S.O. Prognostic implications for high expression of oncogenic microRNAs in advanced gastric carcinoma. J. Surg. Oncol. 2013, 107, 505–510. [Google Scholar]
- Scheffer, A.R.; Holdenrieder, S.; Kristiansen, G.; von Ruecker, A.; Muller, S.C.; Ellinger, J. Circulating microRNAs in serum: Novel biomarkers for patients with bladder cancer? World J. Urol. 2014, 32, 353–358. [Google Scholar]
- Colangelo, T.; Fucci, A.; Votino, C.; Sabatino, L.; Pancione, M.; Laudanna, C.; Binaschi, M.; Bigioni, M.; Maggi, C.A.; Parente, D.; et al. MicroRNA-130b promotes tumor development and is associated with poor prognosis in colorectal cancer. Neoplasia 2013, 15, 1218–1231. [Google Scholar]
- Wu, X.; Weng, L.; Li, X.; Guo, C.; Pal, S.K.; Jin, J.M.; Li, Y.; Nelson, R.A.; Mu, B.; Onami, S.H.; et al. Identification of a 4-microRNA signature for clear cell renal cell carcinoma metastasis and prognosis. PLoS One 2012, 7, e35661. [Google Scholar]
- Kjersem, J.B.; Ikdahl, T.; Lingjaerde, O.C.; Guren, T.; Tveit, K.M.; Kure, E.H. Plasma microRNAs predicting clinical outcome in metastatic colorectal cancer patients receiving first-line oxaliplatin-based treatment. Mol. Oncol. 2014, 8, 59–67. [Google Scholar]
- Liu, A.M.; Yao, T.J.; Wang, W.; Wong, K.F.; Lee, N.P.; Fan, S.T.; Poon, R.T.; Gao, C.; Luk, J.M. Circulating miR-15b and miR-130b in serum as potential markers for detecting hepatocellular carcinoma: A retrospective cohort study. BMJ Open 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Kutay, H.; Bai, S.; Datta, J.; Motiwala, T.; Pogribny, I.; Frankel, W.; Jacob, S.T.; Ghoshal, K. Downregulation of miR-122 in the rodent and human hepatocellular carcinomas. J. Cell Biochem. 2006, 99, 671–678. [Google Scholar]
- Ma, S.; Tang, K.H.; Chan, Y.P.; Lee, T.K.; Kwan, P.S.; Castilho, A.; Ng, I.; Man, K.; Wong, N.; To, K.F.; et al. miR-130b Promotes CD133+ liver tumor-initiating cell growth and self-renewal via tumor protein 53-induced nuclear protein 1. Cell Stem Cell 2010, 7, 694–707. [Google Scholar]
- Yang, M.-H.; Chen, C.-L.; Chau, G.-Y.; Chiou, S.-H.; Su, C.-W.; Chou, T.-Y.; Peng, W.-L.; Wu, J.-C. Comprehensive analysis of the independent effect of twist and snail in promoting metastasis of hepatocellular carcinoma. Hepatology 2009, 50, 1464–1474. [Google Scholar]
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar]
- Petrelli, A.; Perra, A.; Cora, D.; Sulas, P.; Menegon, S.; Manca, C.; Migliore, C.; Kowalik, M.A.; Ledda-Columbano, G.M.; Giordano, S.; et al. MicroRNA/gene profiling unveils early molecular changes and nuclear factor erythroid related factor 2 (NRF2) activation in a rat model recapitulating human hepatocellular carcinoma (HCC). Hepatology 2014, 59, 228–241. [Google Scholar]
- Wang, W.Y.; Zhang, H.F.; Wang, L.; Ma, Y.P.; Gao, F.; Zhang, S.J.; Wang, L.C. High expression of microRNA-130b correlates with poor prognosis of patients with hepatocellular carcinoma. Diagn. Pathol. 2014, 9. [Google Scholar] [CrossRef]
- Koeffler, H.P. Peroxisome proliferator-activated receptor gamma and cancers. Clin. Cancer Res. 2003, 9, 1–9. [Google Scholar]
- Yu, J.; Qiao, L.; Zimmermann, L.; Ebert, M.P.; Zhang, H.; Lin, W.; Rocken, C.; Malfertheiner, P.; Farrell, G.C. Troglitazone inhibits tumor growth in hepatocellular carcinoma in vitro and in vivo. Hepatology 2006, 43, 134–143. [Google Scholar]
- Shen, B.; Chu, E.S.; Zhao, G.; Man, K.; Wu, C.W.; Cheng, J.T.; Li, G.; Nie, Y.; Lo, C.M.; Teoh, N.; et al. PPARγ inhibits hepatocellular carcinoma metastases in vitro and in mice. Br. J. Cancer 2012, 106, 1486–1494. [Google Scholar]
- Lee, E.K.; Lee, M.J.; Abdelmohsen, K.; Kim, W.; Kim, M.M.; Srikantan, S.; Martindale, J.L.; Hutchison, E.R.; Kim, H.H.; Marasa, B.S.; et al. miR-130 suppresses adipogenesis by inhibiting peroxisome proliferator-activated receptor gamma expression. Mol. Cell. Biol. 2011, 31, 626–638. [Google Scholar]
- Tu, K.; Zheng, X.; Zan, X.; Han, S.; Yao, Y.; Liu, Q. Evaluation of Fbxw7 expression and its correlation with the expression of c-Myc, cyclin E and p53 in human hepatocellular carcinoma. Hepatol. Res. 2012, 42, 904–910. [Google Scholar]
- Tu, K.; Yang, W.; Li, C.; Zheng, X.; Lu, Z.; Guo, C.; Yao, Y.; Liu, Q. Fbxw7 is an independent prognostic marker and induces apoptosis and growth arrest by regulating YAP abundance in hepatocellular carcinoma. Mol. Cancer 2014, 13. [Google Scholar] [CrossRef]
- Friedman, S.L. Mechanisms of disease: Mechanisms of hepatic fibrosis and therapeutic implications. Nat. Clin. Pract Gastroenterol. Hepatol. 2004, 1, 98–105. [Google Scholar]
- Li, C.; Yang, W.; Zhang, J.; Zheng, X.; Yao, Y.; Tu, K.; Liu, Q. SREBP-1 has a prognostic role and contributes to invasion and metastasis in human hepatocellular carcinoma. Int. J. Mol. Sci. 2014, 15, 7124–7138. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, K.; Zheng, X.; Dou, C.; Li, C.; Yang, W.; Yao, Y.; Liu, Q. MicroRNA-130b Promotes Cell Aggressiveness by Inhibiting Peroxisome Proliferator-Activated Receptor Gamma in Human Hepatocellular Carcinoma. Int. J. Mol. Sci. 2014, 15, 20486-20499. https://doi.org/10.3390/ijms151120486
Tu K, Zheng X, Dou C, Li C, Yang W, Yao Y, Liu Q. MicroRNA-130b Promotes Cell Aggressiveness by Inhibiting Peroxisome Proliferator-Activated Receptor Gamma in Human Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2014; 15(11):20486-20499. https://doi.org/10.3390/ijms151120486
Chicago/Turabian StyleTu, Kangsheng, Xin Zheng, Changwei Dou, Chao Li, Wei Yang, Yingmin Yao, and Qingguang Liu. 2014. "MicroRNA-130b Promotes Cell Aggressiveness by Inhibiting Peroxisome Proliferator-Activated Receptor Gamma in Human Hepatocellular Carcinoma" International Journal of Molecular Sciences 15, no. 11: 20486-20499. https://doi.org/10.3390/ijms151120486