Inhibition of p53 deSUMOylation Exacerbates Puromycin Aminonucleoside-Induced Apoptosis in Podocytes

Abstract
:1. Introduction
2. Results
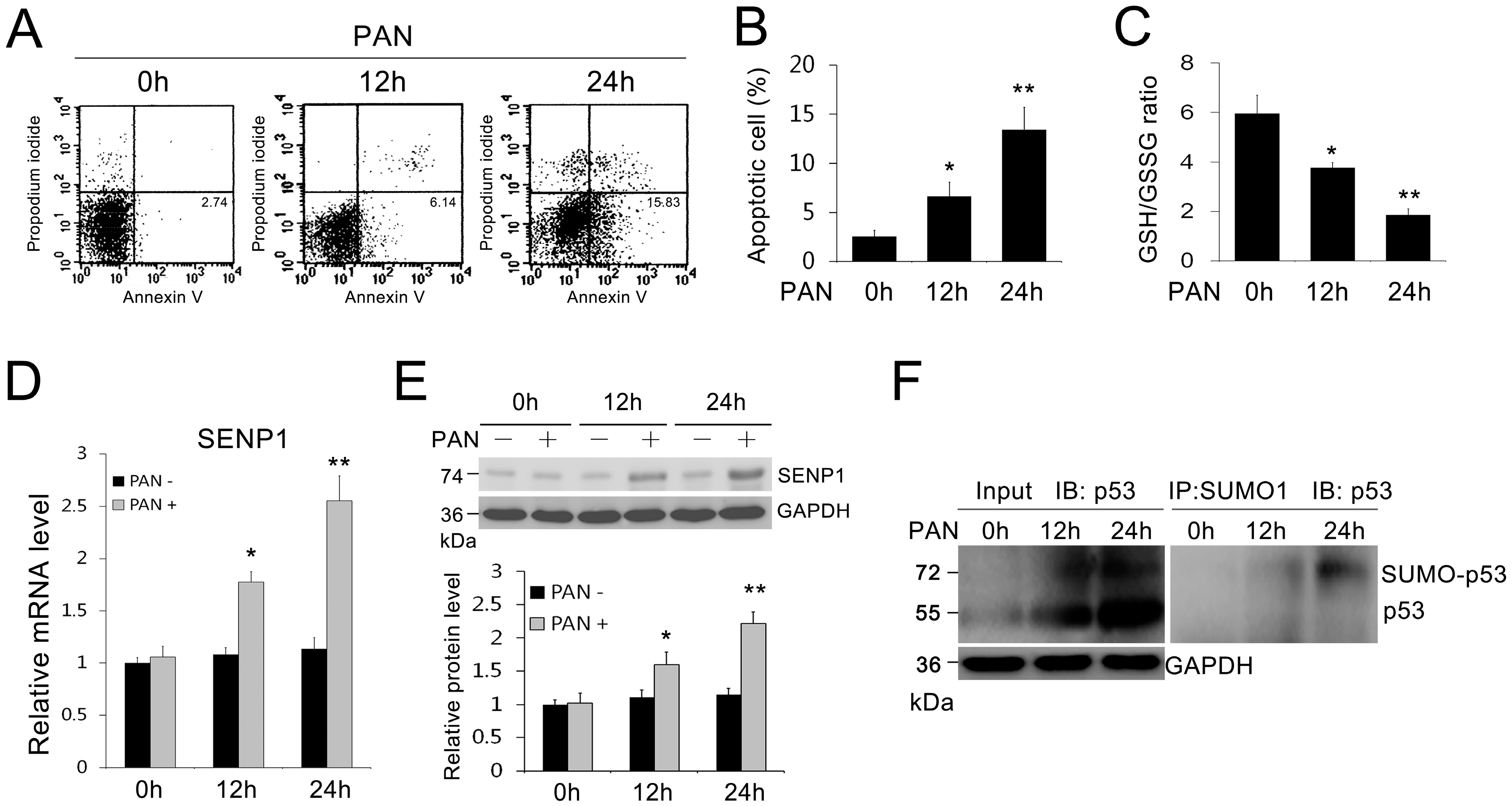
2.1. SENP1 and p53 Expression Increased and Accompanied Podocyte Apoptosis Induced by PAN
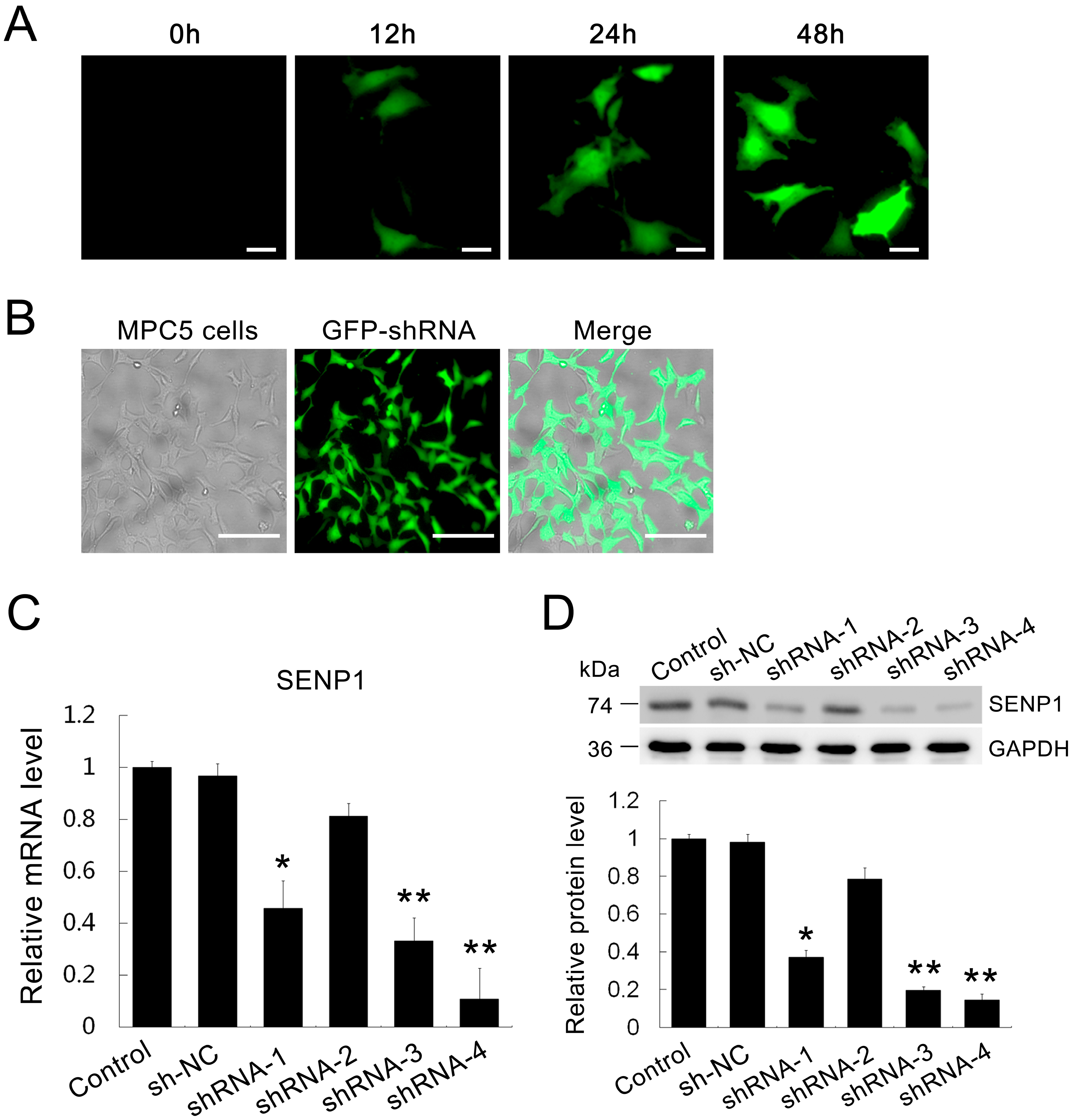
2.2. Establishment of SENP1-Knockdown in MPC5 Cells
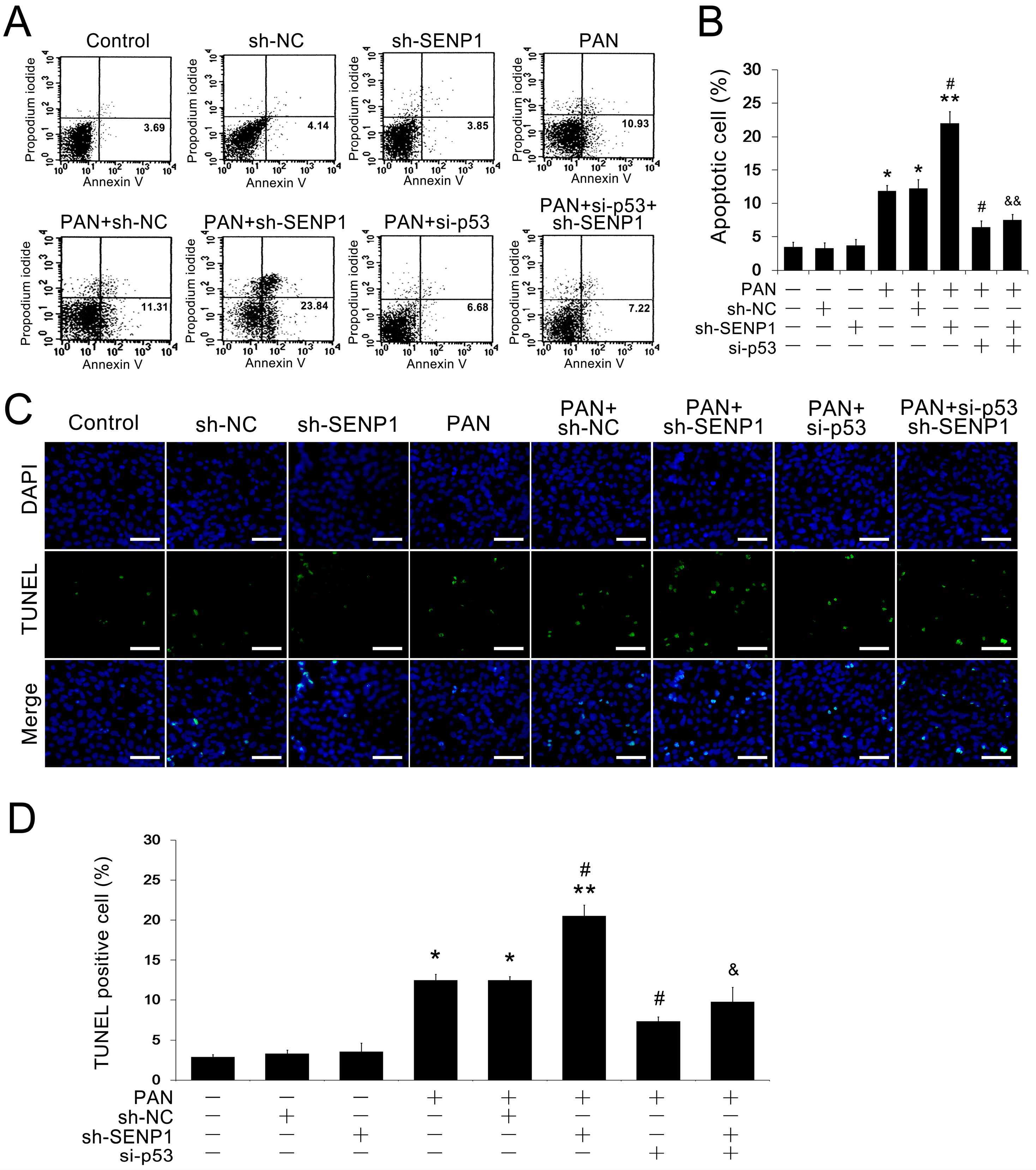
2.3. SENP1 Knockdown Increased PAN-Induced Apoptosis in Podocytes



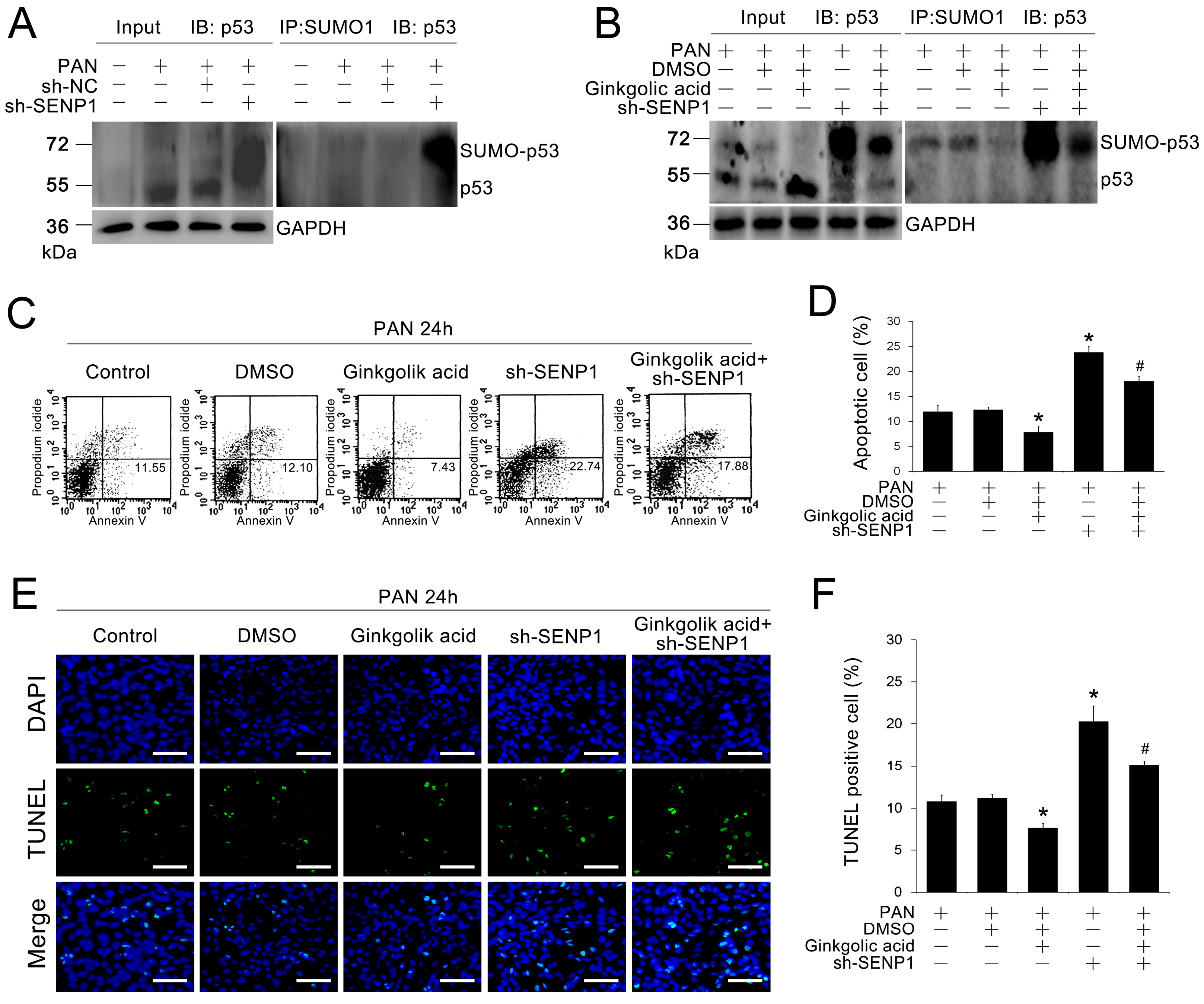
2.4. SENP1 Deficiency Induces Apoptosis in Podocytes through p53 SUMOylation

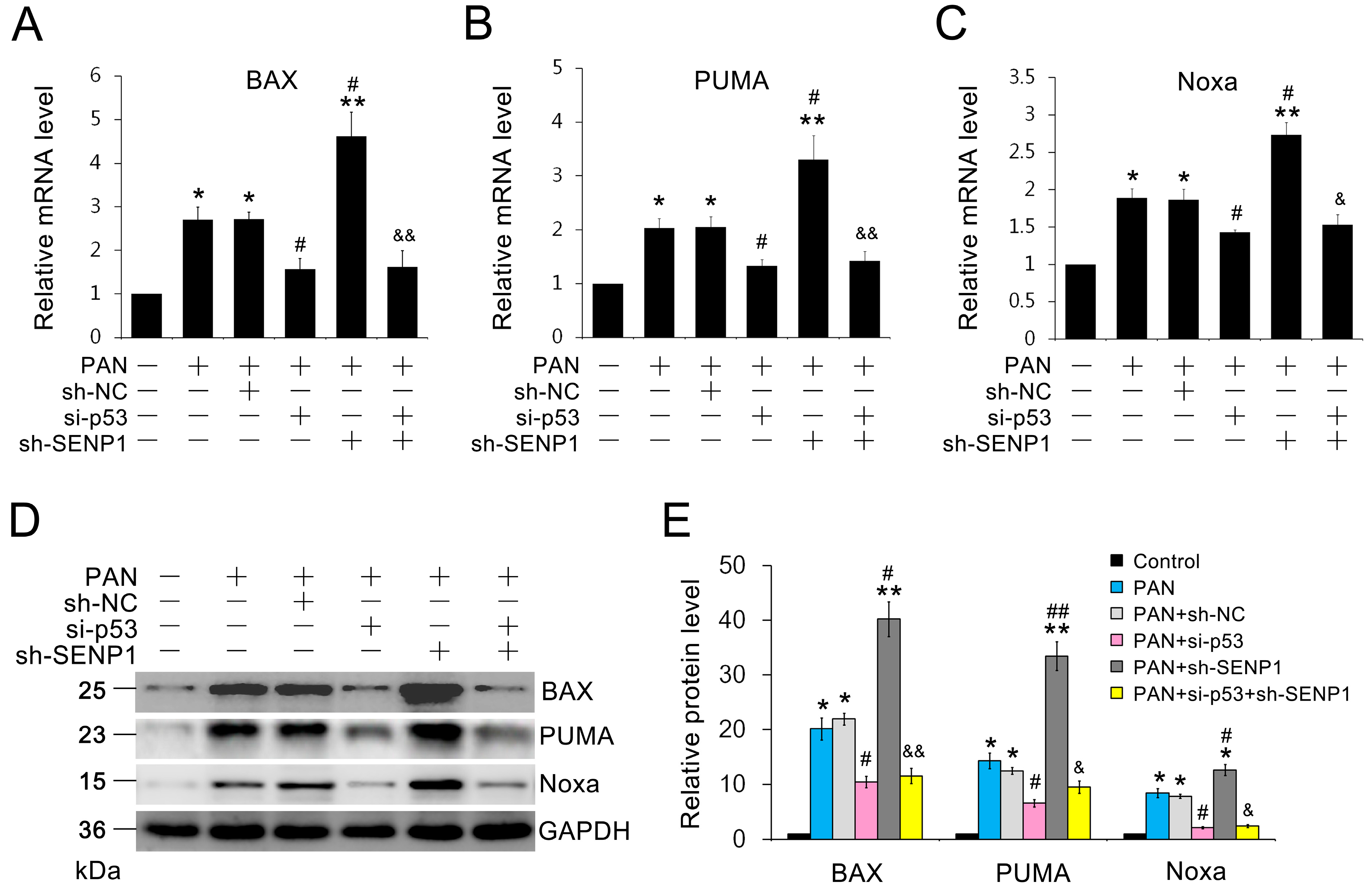
2.5. SENP1 Deficiency Induces the Upregulation of p53-Dependent Pro-Apoptotic Genes in Podocytes

3. Discussion
4. Experimental Section
4.1. Cell Culture
4.2. Small Hairpin RNA (shRNA) Plasmids, Small Interfering RNA(siRNA) and Transient Transfections
4.3. Protein Isolation and Immunoblotting
4.4. Immunoprecipitation
4.5. RNA Isolation and Real-Time RT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/Primer | Direction | Sequence (5' to 3') |
|---|---|---|
| SENP1 | Forward | CTACAAGAAGCCCAGCCTATCGTC |
| Reverse | GTCACCTGAGCCAAGGAAACTG | |
| BAX | Forward | ACCAAGAAGCTGAGCGAGTGT |
| Reverse | ACAAACATGGTCACGGTCTGC | |
| Noxa | Forward | GCAGAGCTGGAAGTCGAGTGT |
| Reverse | AAGTTTCTGCCGGAAGTTCAG | |
| PUMA | Forward | ACGACCTCAACGCACAGTACG |
| Reverse | TCCCATGATGAGATTGTACAGGAC | |
| GAPDH | Forward | GCACCGTCAAGGCTGAGAAC |
| Reverse | TGGTGAAGACGCCAGTGGA |
4.6. FACS Analysis
4.7. TUNEL Assay
4.8. Measurement of Oxidative Stress
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pagtalunan, M.E.; Miller, P.L.; Jumping-Eagle, S.; Nelson, R.G.; Myers, B.D.; Rennke, H.G.; Coplon, N.S.; Sun, L.; Meyer, T.W. Podocyte loss and progressive glomerular injury in type ii diabetes. J. Clin. Investig. 1997, 99, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Kriz, W.; Elger, M.; Nagata, M.; Kretzler, M.; Uiker, S.; Koeppen-Hageman, I.; Tenschert, S.; Lemley, K.V. The role of podocytes in the development of glomerular sclerosis. Kidney Int. Suppl. 1994, 45, S64–S72. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, M.; Najafian, B.; Kim, Y.; Caramori, M.L.; Mauer, M. Podocyte detachment and reduced glomerular capillary endothelial fenestration in human type 1 diabetic nephropathy. Diabetes 2007, 56, 2155–2160. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Goyal, M.; Kurnit, D.; Wharram, B.; Wiggins, J.; Holzman, L.; Kershaw, D.; Wiggins, R. Podocyte depletion and glomerulosclerosis have a direct relationship in the pan-treated rat. Kidney Int. 2001, 60, 957–968. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, M.; Bitzer, M.; Roberts, I.S.; Kopp, J.B.; ten Dijke, P.; Mundel, P.; Bottinger, E.P. Apoptosis in podocytes induced by tgf-beta and smad7. J. Clin. Investig. 2001, 108, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Mundel, P.; Shankland, S.J. Podocyte biology and response to injury. J. Am. Soc. Nephrol. 2002, 13, 3005–3015. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Reddy, K.; Kapasi, A.A.; Franki, N.; Gibbons, N.; Kasinath, B.S.; Singhal, P.C. Angiotensin ii induces apoptosis in rat glomerular epithelial cells. Am. J. Physiol. Renal Physiol. 2002, 283, F173–F180. [Google Scholar] [PubMed]
- Gareau, J.R.; Lima, C.D. The sumo pathway: Emerging mechanisms that shape specificity, conjugation and recognition. Nat. Rev. Mol. Cell Biol. 2010, 11, 861–871. [Google Scholar] [CrossRef] [PubMed]
- Geiss-Friedlander, R.; Melchior, F. Concepts in sumoylation: A decade on. Nat. Rev. Mol. Cell Biol. 2007, 8, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Li, S.J.; Hochstrasser, M. A new protease required for cell-cycle progression in yeast. Nature 1999, 398, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Andreou, A.M.; Tavernarakis, N. Sumoylation and cell signalling. Biotechnol. J. 2009, 4, 1740–1752. [Google Scholar] [CrossRef] [PubMed]
- Bawa-Khalfe, T.; Yeh, E.T. Sumo losing balance: Sumo proteases disrupt sumo homeostasis to facilitate cancer development and progression. Genes Cancer 2010, 1, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Choi, H.J.; Kim, B.; Kim, M.H.; Lee, J.M.; Kim, I.S.; Lee, M.H.; Choi, S.J.; Kim, K.I.; Kim, S.I.; et al. Roles of sumoylation of a reptin chromatin-remodelling complex in cancer metastasis. Nat. Cell Biol. 2006, 8, 631–639. [Google Scholar] [CrossRef]
- Brems-Eskildsen, A.S.; Zieger, K.; Toldbod, H.; Holcomb, C.; Higuchi, R.; Mansilla, F.; Munksgaard, P.P.; Borre, M.; Orntoft, T.F.; Dyrskjot, L. Prediction and diagnosis of bladder cancer recurrence based on urinary content of htert, senp1, ppp1ca, and mcm5 transcripts. BMC Cancer 2010, 10, 646. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Pippin, J.W.; Marshall, C.B.; Griffin, S.V.; Shankland, S.J. Dexamethasone prevents podocyte apoptosis induced by puromycin aminonucleoside: Role of p53 and bcl-2-related family proteins. J. Am. Soc. Nephrol. 2005, 16, 2615–2625. [Google Scholar] [CrossRef] [PubMed]
- Bode, A.M.; Dong, Z. Targeting signal transduction pathways by chemopreventive agents. Mutat. Res. 2004, 555, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Toledo, F.; Wahl, G.M. Regulating the p53 pathway: In vitro hypotheses, in vivo veritas. Nat. Rev. Cancer 2006, 6, 909–923. [Google Scholar] [CrossRef]
- Olsson, A.; Manzl, C.; Strasser, A.; Villunger, A. How important are post-translational modifications in p53 for selectivity in target-gene transcription and tumour suppression? Cell Death Differ. 2007, 14, 1561–1575. [Google Scholar] [CrossRef] [PubMed]
- Gostissa, M.; Hengstermann, A.; Fogal, V.; Sandy, P.; Schwarz, S.E.; Scheffner, M.; del Sal, G. Activation of p53 by conjugation to the ubiquitin-like protein sumo-1. EMBO J. 1999, 18, 6462–6471. [Google Scholar] [CrossRef] [PubMed]
- Stehmeier, P.; Muller, S. Regulation of p53 family members by the ubiquitin-like sumo system. DNA Repair (Amst) 2009, 8, 491–498. [Google Scholar] [CrossRef]
- Mauri, F.; McNamee, L.M.; Lunardi, A.; Chiacchiera, F.; del Sal, G.; Brodsky, M.H.; Collavin, L. Modification of drosophila p53 by sumo modulates its transactivation and pro-apoptotic functions. J. Biol. Chem. 2008, 283, 20848–20856. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.M.; Yang, F.F.; Yuan, Y.F.; Zhai, R.; Huo, L.J. Sumoylation of mouse p53b by sumo-1 promotes its pro-apoptotic function in ovarian granulosa cells. PLoS One 2013, 8, e63680. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Yamada, S.; Lualdi, M.; Dasso, M.; Kuehn, M.R. Senp1 is essential for desumoylating sumo1-modified proteins but dispensable for sumo2 and sumo3 deconjugation in the mouse embryo. Cell Rep. 2013, 3, 1640–1650. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, I.; Ito, A.; Hirai, G.; Nishimura, S.; Kawasaki, H.; Saitoh, H.; Kimura, K.; Sodeoka, M.; Yoshida, M. Ginkgolic acid inhibits protein sumoylation by blocking formation of the e1-sumo intermediate. Chem. Biol. 2009, 16, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, T.; Krajewski, S.; Krajewska, M.; Wang, H.G.; Lin, H.K.; Liebermann, D.A.; Hoffman, B.; Reed, J.C. Tumor suppressor p53 is a regulator of bcl-2 and bax gene expression in vitro and in vivo. Oncogene 1994, 9, 1799–1805. [Google Scholar]
- Nakajima, W.; Tanaka, N. Synergistic induction of apoptosis by p53-inducible bcl-2 family proteins noxa and puma. J. Nippon Med. Sch. 2007, 74, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Villunger, A.; Michalak, E.M.; Coultas, L.; Mullauer, F.; Bock, G.; Ausserlechner, M.J.; Adams, J.M.; Strasser, A. P53- and drug-induced apoptotic responses mediated by bh3-only proteins puma and noxa. Science 2003, 302, 1036–1038. [Google Scholar] [CrossRef] [PubMed]
- Oda, E.; Ohki, R.; Murasawa, H.; Nemoto, J.; Shibue, T.; Yamashita, T.; Tokino, T.; Taniguchi, T.; Tanaka, N. Noxa, a bh3-only member of the bcl-2 family and candidate mediator of p53-induced apoptosis. Science 2000, 288, 1053–1058. [Google Scholar] [CrossRef] [PubMed]
- Shibue, T.; Takeda, K.; Oda, E.; Tanaka, H.; Murasawa, H.; Takaoka, A.; Morishita, Y.; Akira, S.; Taniguchi, T.; Tanaka, N. Integral role of noxa in p53-mediated apoptotic response. Genes Dev. 2003, 17, 2233–2238. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Chiu, S.Y.; Hsu, W. Sumo-specific protease 2 in mdm2-mediated regulation of p53. Cell Death Differ. 2011, 18, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Heo, K.S.; Chang, E.; Le, N.T.; Cushman, H.; Yeh, E.T.; Fujiwara, K.; Abe, J. De-sumoylation enzyme of sentrin/sumo-specific protease 2 regulates disturbed flow-induced sumoylation of erk5 and p53 that leads to endothelial dysfunction and atherosclerosis. Circ. Res. 2013, 112, 911–923. [Google Scholar] [CrossRef] [PubMed]
- Kolli, N.; Mikolajczyk, J.; Drag, M.; Mukhopadhyay, D.; Moffatt, N.; Dasso, M.; Salvesen, G.; Wilkinson, K.D. Distribution and paralogue specificity of mammalian desumoylating enzymes. Biochem. J. 2010, 430, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Fan, Q.; Zhang, Z.; Zou, Y.; Cai, R.; Wang, Q.; Zuo, Y.; Cheng, J. Senp1 deficiency promotes er stress-induced apoptosis by increasing xbp1 sumoylation. Cell Cycle 2012, 11, 1118–1122. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.B.; Gao, Q.M.; Liang, W.; Xiu, B.; Zhang, W.J.; Liang, A.B. Down-regulation of senp1 expression increases apoptosis of burkitt lymphoma cells. Asian Pac. J. Cancer Prev. 2012, 13, 2045–2049. [Google Scholar] [CrossRef] [PubMed]
- Yates, K.E.; Korbel, G.A.; Shtutman, M.; Roninson, I.B.; DiMaio, D. Repression of the sumo-specific protease senp1 induces p53-dependent premature senescence in normal human fibroblasts. Aging Cell. 2008, 7, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Oren, M. Regulation of the p53 tumor suppressor protein. J. Biol. Chem. 1999, 274, 36031–36034. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.M.; Phillips, A.C.; Vousden, K.H. Regulation and function of the p53 tumor suppressor protein. Curr. Opin. Cell Biol. 2001, 13, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.S.; Desterro, J.M.; Lain, S.; Midgley, C.A.; Lane, D.P.; Hay, R.T. Sumo-1 modification activates the transcriptional response of p53. EMBO J. 1999, 18, 6455–6461. [Google Scholar] [CrossRef] [PubMed]
- Kwek, S.S.; Derry, J.; Tyner, A.L.; Shen, Z.; Gudkov, A.V. Functional analysis and intracellular localization of p53 modified by sumo-1. Oncogene 2001, 20, 2587–2599. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Bawa, T.; Lee, P.; Gong, L.; Yeh, E.T. Role of desumoylation in the development of prostate cancer. Neoplasia 2006, 8, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, J.; Zuo, Y.; Deng, J.; Wang, L.S.; Chen, G.Q. Sumo-specific protease 1 regulates the in vitro and in vivo growth of colon cancer cells with the upregulated expression of cdk inhibitors. Cancer Lett. 2011, 309, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xia, N.; Li, T.; Xu, Y.; Zou, Y.; Zuo, Y.; Fan, Q.; Bawa-Khalfe, T.; Yeh, E.T.; Cheng, J. Sumo-specific protease 1 promotes prostate cancer progression and metastasis. Oncogene 2013, 32, 2493–2498. [Google Scholar] [CrossRef] [PubMed]
- Feligioni, M.; Brambilla, E.; Camassa, A.; Sclip, A.; Arnaboldi, A.; Morelli, F.; Antoniou, X.; Borsello, T. Crosstalk between jnk and sumo signaling pathways: Desumoylation is protective against h2o2-induced cell injury. PLoS One 2011, 6, e28185. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Fan, Y.; Liu, X.; Zhou, L.; Cheng, J.; Cai, R.; Xue, S. Senp1 protects against myocardial ischemia/reperfusion injury via a hif1alpha-dependent pathway. Cardiovasc. Res. 2014, 104, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Meinecke, I.; Cinski, A.; Baier, A.; Peters, M.A.; Dankbar, B.; Wille, A.; Drynda, A.; Mendoza, H.; Gay, R.E.; Hay, R.T.; et al. Modification of nuclear pml protein by sumo-1 regulates fas-induced apoptosis in rheumatoid arthritis synovial fibroblasts. Proc. Natl. Acad. Sci. USA 2007, 104, 5073–5078. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Luo, Y.; Yu, L.; Lin, Y.; Luo, D.; Zhang, H.; He, Y.; Kim, Y.O.; Kim, Y.; Tang, S.; et al. Senp1 mediates tnf-induced desumoylation and cytoplasmic translocation of hipk1 to enhance ask1-dependent apoptosis. Cell Death Differ. 2008, 15, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.; Herr, D.; Chun, J.; Xu, Y. Ser18 and 23 phosphorylation is required for p53-dependent apoptosis and tumor suppression. EMBO J. 2006, 25, 2615–2622. [Google Scholar] [PubMed]
- Feng, L.; Hollstein, M.; Xu, Y. Ser46 phosphorylation regulates p53-dependent apoptosis and replicative senescence. Cell Cycle 2006, 5, 2812–2819. [Google Scholar] [CrossRef] [PubMed]
- Marchenko, N.D.; Wolff, S.; Erster, S.; Becker, K.; Moll, U.M. Monoubiquitylation promotes mitochondrial p53 translocation. EMBO J. 2007, 26, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Vucic, D.; Dixit, V.M.; Wertz, I.E. Ubiquitylation in apoptosis: A post-translational modification at the edge of life and death. Nat. Rev. Mol. Cell Biol. 2011, 12, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Mundel, P.; Reiser, J.; Zuniga Mejia Borja, A.; Pavenstadt, H.; Davidson, G.R.; Kriz, W.; Zeller, R. Rearrangements of the cytoskeleton and cell contacts induce process formation during differentiation of conditionally immortalized mouse podocyte cell lines. Exp. Cell Res. 1997, 236, 248–258. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Zhu, J.; Fang, M.; Zhang, T.; Xie, H.; Wang, N.; Shen, N.; Guo, H.; Fu, B.; Lin, H. Inhibition of p53 deSUMOylation Exacerbates Puromycin Aminonucleoside-Induced Apoptosis in Podocytes. Int. J. Mol. Sci. 2014, 15, 21314-21330. https://doi.org/10.3390/ijms151121314
Wang L, Zhu J, Fang M, Zhang T, Xie H, Wang N, Shen N, Guo H, Fu B, Lin H. Inhibition of p53 deSUMOylation Exacerbates Puromycin Aminonucleoside-Induced Apoptosis in Podocytes. International Journal of Molecular Sciences. 2014; 15(11):21314-21330. https://doi.org/10.3390/ijms151121314
Chicago/Turabian StyleWang, Lingyu, Jingwei Zhu, Ming Fang, Tuaner Zhang, Hua Xie, Nan Wang, Nan Shen, Hui Guo, Bo Fu, and Hongli Lin. 2014. "Inhibition of p53 deSUMOylation Exacerbates Puromycin Aminonucleoside-Induced Apoptosis in Podocytes" International Journal of Molecular Sciences 15, no. 11: 21314-21330. https://doi.org/10.3390/ijms151121314