The Cyclic Antibacterial Peptide Enterocin AS-48: Isolation, Mode of Action, and Possible Food Applications

Abstract
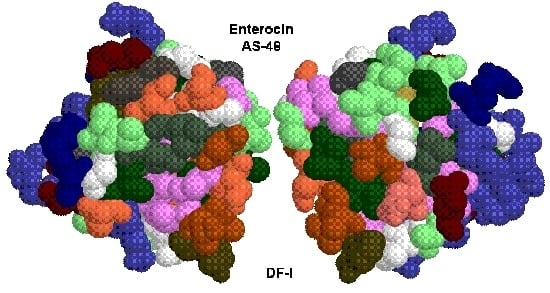
:
1. Introduction
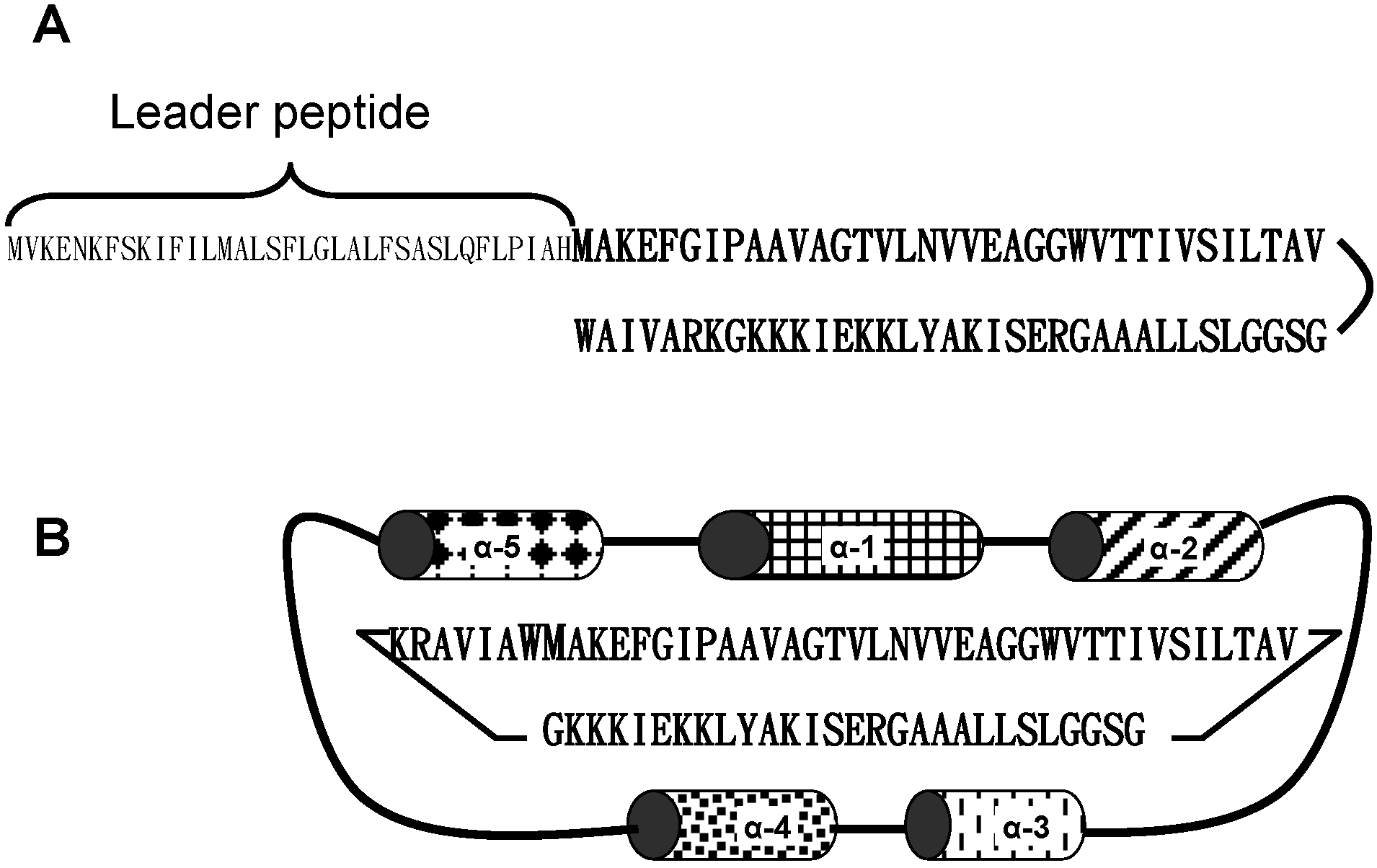
2. The Bacteriocin Enterocin AS-48

3. Inhibitory Spectrum of Enterocin AS-48
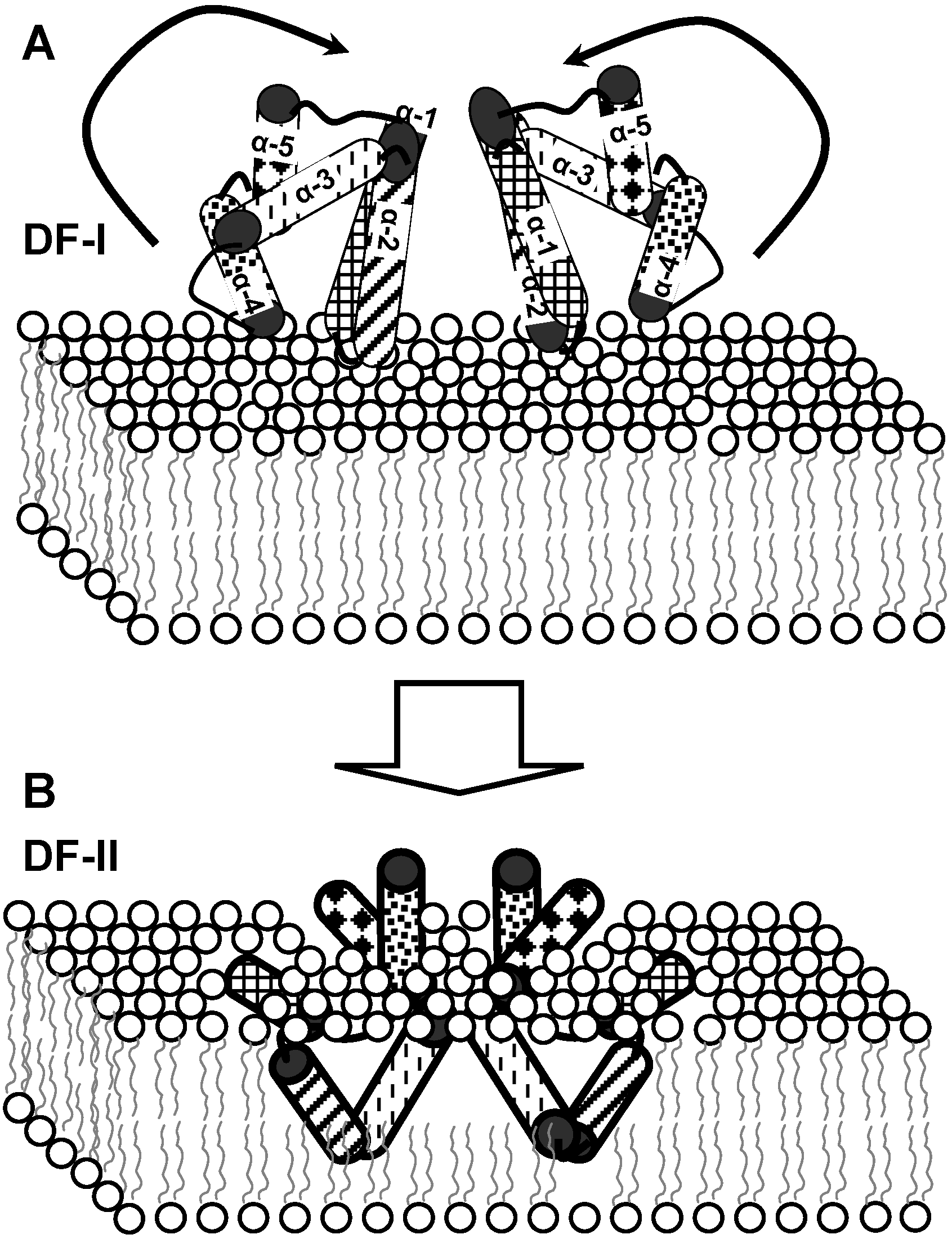
4. Bacteriocin Mode of Action

5. Antibacterial Effects of Enterocin AS-48 in Food Systems

{kind=link}
{kind=link}
{kind=link}
| Target Bacterium | Food Substrate | Reference(s) |
|---|---|---|
| Bacillus cereus | Cheese, rice gruel, cooked rice, sprouts, green asparagus, whole fruit pieces and sliced fruits, vegetable soups, wheat dough, desserts | [55,76,82,92,94,96] |
| Bacillus weihenstephanensis | Sprouts, green asparagus | [82] |
| Bacillus licheniformis | Apple cider, energy drinks, wheat dough | [56,91,94] |
| Bacillus coagulans | Canned foods | [93] |
| Bacillus subtilis | Wheat dough | [94] |
| Paenibacillus spp. | Vegetable soups | [92] |
| Geobacillus stearothermophilus | Canned foods and coconut juice | [58] |
| Alicyclobacillus acidoterrestris | Fruit juices | [57] |
| Listeria monocytogenes | Meat sausages, cooked ham, fermented sausage, skim milk, sprouts, green asparagus, whole fruit pieces and sliced fruits, ready-to-eat salads, energy drinks, desserts | [42,71,73,74,80,81,85,91,96] |
| Staphylococcus aureus | Meat sausages, cooked ham, fermented sausage, skim milk, cheese, sauces, energy drinks, bakery ingredients, desserts | [42,72,74,77,78,87,91,95,96] |
| Staphylococcus carnosus | Cooked ham | [75] |
| Brochothrix thermosphacta | Cooked ham | [75] |
| Lactobacillus sakei | Cooked ham | [75] |
| Lactobacillus collinoides | Apple cider | [88] |
| Lactobacillus diolivorans | Apple cider | [88] |
| Pediococcus parvulus | Apple cider | [88] |
| Escherichia coli | Apple juice, soybean sprouts | [50,83] |
| Salmonella enterica | Fermented sausage, soybean sprouts, ready-to-eat salads, apple juice | [52,74,83,86] |
| Shigella flexneri | Soybean sprouts | [83] |
| Enterobacter aerogenes | Soybean sprouts | [83] |
| Yersinia enterocolitica | Soybean sprouts | [83] |
| Aeromonas hydrophila | Soybean sprouts | [83] |
| Pseudomonas fluorescens | Soybean sprouts | [83] |
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jack, R.W.; Tagg, J.R.; Ray, B. Bacteriocins of Gram-positive bacteria. Microbiol. Mol. Biol. Rev. 1995, 59, 171–200. [Google Scholar]
- Cotter, P.D.; Hill, C.; Ross, R.P. Bacteriocins: Developing innate immunity for food. Nat. Rev. Microbiol. 2005, 3, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, A.; Giménez-Gallego, G.; Maqueda, M.; Valdivia, E. Purification and amino acid composition of peptide antibiotic AS-48 produced by Streptococcus (Enterococcus) faecalis subsp. liquefaciens S-48. Antimicrob. Agents Chemother. 1989, 33, 437–441. [Google Scholar] [CrossRef]
- Samyn, B.; Martínez-Bueno, M.; Devreese, B.; Maqueda, M.; Gálvez, A.; Valdivia, E.; Coyette, J.; van Beeumen, J. The cyclic structure of the enterococcal peptide antibiotic AS-48. FEBS Lett. 1994, 352, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Bueno, M.; Maqueda, M.; Gálvez, A.; Samyn, B.; van Beeumen, J.; Coyette, J.; Valdivia, E. Determination of the gene sequence and the molecular structure of the enterococcal peptide antibiotic AS-48. J. Bacteriol. 1994, 176, 6334–6339. [Google Scholar] [PubMed]
- Martínez-Bueno, M.; Valdivia, E.; Gálvez, A.; Coyette, J.; Maqueda, M. Analysis of the gene cluster involved in production and immunity of the peptide antibiotic AS-48 in Enterococcus faecalis. Mol. Microbiol. 1998, 27, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Maqueda, M.; Gálvez, A.; Bueno, M.M.; Sanchez-Barrena, M.J.; González, C.; Albert, A.; Rico, M.; Valdivia, E. Peptide AS-48: Prototype of a new class of cyclic bacteriocins. Curr. Protein Pept. Sci. 2004, 5, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Gabrielsen, C.; Brede, D.A.; Nes, I.F.; Diep, D.B. Circular bacteriocins: Biosynthesis and mode of action. Appl. Environ. Microbiol. 2014, 80, 6854–6862. [Google Scholar] [CrossRef] [PubMed]
- Maqueda, M.; Sánchez-Hidalgo, M.; Fernández, M.; Montalbán-López, M.; Valdivia, E.; Martínez-Bueno, M. Genetic features of circular bacteriocins produced by Gram-positive bacteria. FEMS Microbiol. Rev. 2008, 32, 2–22. [Google Scholar] [CrossRef] [PubMed]
- Montalbán-López, M.; Sánchez-Hidalgo, M.; Cebrián, R.; Maqueda, M. Discovering the bacterial circular proteins: Bacteriocins, cyanobactins, and pilins. J. Biol. Chem. 2012, 287, 27007–27013. [Google Scholar] [CrossRef] [PubMed]
- Van Belkum, M.J.; Martin-Visscher, L.A.; Vederas, J.C. Structure and genetics of circular bacteriocins. Trends Microbiol. 2011, 19, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Borrero, J.; Brede, D.A.; Skaugen, M.; Diep, D.B.; Herranz, C.; Nes, I.F.; Cintas, L.M.; Hernández, P.E. Characterization of garvicin ML, a novel circular bacteriocin produced by Lactococcus garvieae DCC43, isolated from mallard ducks (Anas platyrhynchos). Appl. Environ. Microbiol. 2011, 77, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Wirawan, R.E.; Swanson, K.M.; Kleffmann, T.; Jack, R.W.; Tagg, J.R. Uberolysin: A novel cyclic bacteriocin produced by Streptococcus uberis. Microbiology 2007, 153, 1619–1630. [Google Scholar] [CrossRef] [PubMed]
- Martin-Visscher, L.A.; van Belkum, M.J.; Garneau-Tsodikova, S.; Whittal, R.M.; Zheng, J.; McMullen, L.M.; Vederas, J.C. Isolation and characterization of carnocyclin A, a novel circular bacteriocin produced by Carnobacterium maltaromaticum UAL307. Appl. Environ. Microbiol. 2008, 74, 4756–4763. [Google Scholar] [CrossRef] [PubMed]
- Sawa, N.; Zendo, T.; Kiyofuji, J.; Fujita, K.; Himeno, K.; Nakayama, J.; Sonomoto, K. Identification and characterization of lactocyclicin Q, a novel cyclic bacteriocin produced by Lactococcus sp. strain QU 12. Appl. Environ. Microbiol. 2009, 75, 1552–1558. [Google Scholar] [CrossRef] [PubMed]
- Masuda, Y.; Ono, H.; Kitagawa, H.; Ito, H.; Mu, F.; Sawa, N.; Zendo, T.; Sonomoto, K. Identification and characterization of leucocyclicin Q, a novel cyclic bacteriocin produced by Leuconostoc mesenteroides TK41401. Appl. Environ. Microbiol. 2011, 77, 8164–8170. [Google Scholar] [CrossRef] [PubMed]
- Scholz, R.; Vater, J.; Budiharjo, A.; Wang, Z.; He, Y.; Dietel, K.; Schwecke, T.; Herfort, S.; Lasch, P.; Borriss, R. Amylocyclicin, a novel circular bacteriocin produced by Bacillus amyloliquefaciens FZB42. J. Bacteriol. 2014, 196, 1842–1852. [Google Scholar] [CrossRef] [PubMed]
- Kemperman, R.; Kuipers, A.; Karsens, H.; Nauta, A.; Kuipers, O.; Kok, J. Identification and characterization of two novel clostridial bacteriocins, circularin A and closticin 574. Appl. Environ. Microbiol. 2003, 69, 1589–1597. [Google Scholar] [CrossRef] [PubMed]
- Kawai, Y.; Saito, T.; Kitazawa, H.; Itoh, T.; Gassericin, A. An uncommon cyclic bacteriocin produced by Lactobacillus gasseri LA39 linked at N- and C-terminal ends. Biosci. Biotechnol. Biochem. 1998, 62, 2438–2440. [Google Scholar] [CrossRef] [PubMed]
- Kawai, Y.; Ishii, Y.; Uemura, K.; Kitazawa, H.; Saito, T.; Itoh, T. Lactobacillus reuteri LA6 and Lactobacillus gasseri LA39 isolated from faeces of the same human infant produce identical cyclic bacteriocin. Food Microbiol. 2001, 18, 407–415. [Google Scholar] [CrossRef]
- Arakawa, K.; Kawai, Y.; Ito, Y.; Nakamura, K.; Chujo, T.; Nishimura, J.; Kitazawa, H.; Saito, T. HPLC purification and re-evaluation of chemical identity of two circular bacteriocins, gassericin A and reutericin 6. Lett. Appl. Microbiol. 2010, 50, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Kalmokoff, M.L.; Cyr, T.D.; Hefford, M.A.; Whitford, M.F.; Teather, R.M. Butyrivibriocin AR10, a new cyclic bacteriocin produced by the ruminal anaerobe Butyrivibrio fibrisolvens AR10: Characterization of the gene and peptide. Can. J. Microbiol. 2003, 49, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, J.; Montville, T.J.; Nes, I.F.; Chikindas, M.L. Bacteriocins: Safe, natural antimicrobials for food preservation. Int. J. Food Microbiol. 2001, 71, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Zhang, C.; Wang, Y.; Shi, J.; Zhang, L.; Ding, Z.; Qu, X.; Cui, H. Class IIa bacteriocins: Diversity and new developments. Int. J. Mol. Sci. 2012, 13, 16668–16707. [Google Scholar] [CrossRef] [PubMed]
- Diep, D.B.; Nes, I.F. Ribosomally synthesized antibacterial peptides in Gram Positive bacteria. Curr. Drug Targets 2002, 3, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Franz, C.M.A.P.; van Belkum, M.J.; Holzapfel, W.H.; Abriouel, H.; Gálvez, A. Diversity of enterococcal bacteriocins and their grouping into a new classification scheme (review). FEMS Microbiol. Rev. 2007, 31, 293–310. [Google Scholar] [CrossRef] [PubMed]
- Juneja, V.K.; Dwivedi, H.P.; Yan, X. Novel natural food antimicrobials. Annu. Rev. Food Sci. Technol. 2012, 3, 381–403. [Google Scholar] [CrossRef] [PubMed]
- Nissen-Meyer, J.; Rogne, P.; Oppergard, C.; Haugen, H.S.; Kristiansen, P.E. Structure-function relationships of the non-lanthionine-containing peptide (class II) bacteriocins produced by Gram Positive bacteria. Curr. Pharm. Biotechnol. 2009, 10, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Papagianni, M.; Anastasiadou, S. Pediocins: The bacteriocins of pediococci. Sources, production, properties and applications. Microb. Cell Fact. 2009, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Rea, M.C.; Ross, P.; Cotter, P.D.; Hill, C. Classifications of bacteriocins from Gram Positive bacteria. In Prokaryotic Antimicrobial Peptides: From Genes to Applications; Drider, D., Rebuffat, S., Eds.; Springer: New York, NY, USA, 2011; pp. 29–53. [Google Scholar]
- Gálvez, A.; Maqueda, M.; Valdivia, E.; Quesada, A.; Montoya, E. Characterization and partial purification of a broad spectrum antibiotic AS-48 produced by Streptococcus faecalis. Can. J. Microbiol. 1986, 32, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Tomita, H.; Fujimoto, S.; Tanimoto, K.; Ike, Y. Cloning and genetic and sequence analyses of the bacteriocin 21 determinant encoded on the Enterococcus faecalis pheromone-responsive conjugative plasmid pPD1. J. Bacteriol. 1997, 179, 7843–7855. [Google Scholar] [PubMed]
- Folli, C.; Ramazzina, I.; Arcidiaco, P.; Stoppini, M.; Berni, R. Purification of bacteriocin AS-48 from an Enterococcus faecium strain and analysis of the gene cluster involved in its production. FEMS Microbiol. Lett. 2003, 221, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Joosten, H.M.L.J.; Nuñez, M.; Devreese, B.; van Beeumen, J.; Marugg, J.D. Purification and characterization of enterocin 4, a bacteriocin produced by Enterococcus faecalis INIA4. Appl. Environ. Microbiol. 1996, 62, 4220–4223. [Google Scholar] [PubMed]
- Maisnier-Patin, S.; Forni, E.; Richard, J. Purification, partial characterisation and mode of action of enteroccocin EFS2, an antilisterial bacteriocin produced by a strain of Enterococcus faecalis isolated from a cheese. Int. J. Food Microbiol. 1996, 30, 255–270. [Google Scholar] [CrossRef] [PubMed]
- Maqueda, M.; Gálvez, A.; Martínez-Bueno, M.; Valdivia, E. Widespread production of AS-48-like bacteriocins in strains of Enterococcus faecalis? Mol. Microbiol. 1998, 29, 1318–1319. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.; Zhang, L.; Chung, Y.K.; Zheng, Z.; Yousef, A.E. Characterization and application of enterocin RM6, a bacteriocin from Enterococcus faecalis. BioMed Res. Int. 2013, 2013, 206917. [Google Scholar] [PubMed]
- Cebrián, R.; Baños, A.; Valdivia, E.; Pérez-Pulido, R.; Martínez-Bueno, M.; Maqueda, M. Characterization of functional, safety, and probiotic properties of Enterococcus faecalis UGRA10, a new AS-48-producer strain. Food Microbiol. 2012, 30, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Abriouel, H.; Lucas, R.; Ben Omar, N.; Valdivia, E.; Maqueda, M.; Martínez-Cañamero, M.; Gálvez, A. Enterocin AS-48RJ: A variant of enterocin AS-48 chromosomally encoded by Enterococcus faecium RJ16 isolated from food. Syst. Appl. Microbiol. 2005, 28, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Abriouel, H.; Valdivia, E.; Martínez-Bueno, M.; Maqueda, M.; Gálvez, A. A simple method for semi-preparative-scale production and recovery of enterocin AS-48 derived from Enterococcus faecalis subsp. liquefaciens A-48–32. J. Microbiol. Methods 2003, 55, 599–605. [Google Scholar] [CrossRef]
- Ananou, S.; Muñoz, A.; Gálvez, A.; Martínez-Bueno, M.; Maqueda, M.; Valdivia, E. Optimization of the production of enterocin AS-48 on a whey-based substrate. Int. Dairy J. 2008, 18, 923–927. [Google Scholar] [CrossRef]
- Ananou, S.; Muñoz, A.; Martínez-Bueno, M.; González-Tello, P.; Gálvez, A.; Maqueda, M.; Valdivia, E. Evaluation of an enterocin AS-48 containing bioactive powder obtained by spray-drying. Food Microbiol. 2010, 27, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Langdon, G.M.; Bruix, M.; Gálvez, A.; Valdivia, E.; Maqueda, M.; Rico, M. Sequence-specific 1H assignment and secondary structure of the bacteriocin AS-48 cyclic peptide. J. Biomol. NMR 1998, 12, 173–175. [Google Scholar] [CrossRef] [PubMed]
- González, C.; Langdon, G.M.; Bruix, M.; Gálvez, A.; Valdivia, E.; Maqueda, M.; Rico, M. Bacteriocin AS-48, a cyclic polypeptide structurally and functionally related to mammalian NK-lysin. Proc. Natl. Acad. Sci. USA 2000, 97, 11221–11226. [Google Scholar] [CrossRef] [PubMed]
- Montalbán-López, M.; Spolaore, B.; Pinato, O.; Martínez-Bueno, M.; Valdivia, E.; Maqueda, M.; Fontana, A. Characterization of linear forms of the circular enterocin AS-48 obtained by limited proteolysis. FEBS Lett. 2008, 582, 3237–3242. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.; Martínez-Bueno, M.; Martín, M.C.; Valdivia, E.; Maqueda, M. Heterologous expression of enterocin AS-48 in several strains of lactic acid bacteria. J. Appl. Microbiol. 2007, 102, 1350–1361. [Google Scholar] [CrossRef] [PubMed]
- Montalbán-López, M.; Martínez-Bueno, M.; Valdivia, E.; Maqueda, M. Expression of linear permutated variants from circular enterocin AS-48. Biochimie 2011, 93, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, A.; Maqueda, M.; Martínez-Bueno, M.; Valdivia, E. Bactericidal and bacteriolytic action of peptide antibiotic AS-48 against Gram-positive and Gram-negative bacteria and other organisms. Res. Microbiol. 1989, 140, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Abriouel, H.; Valdivia, E.; Gálvez, A.; Maqueda, M. Response of Salmonella choleraesuis LT2 spheroplasts and permeabilized cells to the bacteriocin AS-48. Appl. Environ. Microbiol. 1998, 64, 4623–4626. [Google Scholar] [PubMed]
- Ananou, S.; Gálvez, A.; Martínez-Bueno, M.; Maqueda, M.; Valdivia, E. Synergistic effect of enterocin AS-48 in combination with outer membrane permeabilizing treatments against Escherichia coli O157:H7. J. Appl. Microbiol. 2005, 99, 1364–1372. [Google Scholar] [CrossRef] [PubMed]
- Grande Burgos, M.J.; Lucas López, R.; López Aguayo, M.C.; Pérez Pulido, R.; Gálvez, A. Inhibition of planktonic and sessile Salmonella enterica cells by combinations of enterocin AS-48, polymyxin B and biocides. Food Control 2012, 30, 214–221. [Google Scholar] [CrossRef]
- Martínez-Viedma, P.; Sobrino, A.; Ben Omar, N.; Abriouel, H.; Lucas López, R.; Valdivia, E.; Martín Belloso, O.; Gálvez, A. Enhanced bactericidal effect of high-intensity pulsed-electric field treatment in combination with enterocin AS-48 against Salmonella enterica in apple juice. Int. J. Food Microbiol. 2008, 128, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Abriouel, H.; Maqueda, M.; Gálvez, A.; Martínez-Bueno, M.; Valdivia, E. Inhibition of bacterial growth, enterotoxin production and spore outgrowth on strains of Bacillus cereus by bacteriocin AS-48. Appl. Environ. Microbiol. 2002, 68, 1473–1477. [Google Scholar] [CrossRef] [PubMed]
- Gould, G.W. Germination and the problem of dormancy. J. Appl. Bacteriol. 1970, 33, 34–49. [Google Scholar] [CrossRef] [PubMed]
- Grande, M.J.; Lucas, R.; Abriouel, H.; Valdivia, E.; Ben Omar, N.; Maqueda, M.; Martínez-Bueno, M.; Martínez-Cañamero, M.; Gálvez, A. Inhibition of toxicogenic Bacillus cereus in rice-based foods by enterocin AS-48. Int. J. Food Microbiol. 2006, 106, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Grande, M.J.; Lucas, R.; Abriouel, H.; Valdivia, E.; Ben Omar, N.; Maqueda, M.; Martínez-Cañamero, M.; Gálvez, A. Inhibition of Bacillus licheniformis LMG 19409 from ropy cider by enterocin AS-48. J. Appl. Microbiol. 2006, 101, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Grande, M.J.; Lucas, R.; Abriouel, H.; Ben Omar, N.; Maqueda, M.; Martínez-Bueno, M.; Martínez-Cañamero, M.; Valdivia, E.; Gálvez, A. Control of Alicyclobacillus acidoterrestris in fruit juices by enterocin AS-48. Int. J. Food Microbiol. 2005, 104, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Viedma, P.; Abriouel, H.; Ben Omar, N.; Lucas, R.; Valdivia, E.; Gálvez, A. Inactivation of Geobacillus stearothermophilus in canned foods and drinks by addition of enterocin AS-48. Food Microbiol. 2009, 26, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, A.; Valdivia, E.; Martínez, M.; Maqueda, M. Effect of peptide AS-48 on Enterococcus faecalis subsp. liquefaciens S-47. Antimicrob. Agents Chemother. 1989, 33, 641–645. [Google Scholar] [CrossRef]
- Gálvez, A.; Valdivia, E.; Martínez, M.; Maqueda, M. Bactericidal action of peptide antibiotic AS-48 against Escherichia coli K-12. Can. J. Microbiol. 1989, 35, 318–321. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, A.; Maqueda, M.; Martínez-Bueno, M.; Valdivia, E. Permeation of bacterial cells, permeation of cytoplasmic and artificial membrane vesicles, and channel formation on lipid bilayers by peptide antibiotic AS-48. J. Bacteriol. 1991, 173, 886–892. [Google Scholar] [PubMed]
- Gong, X.; Martin-Visscher, L.A.; Nahirney, D.; Vederas, J.C.; Duszyk, M. The circular bacteriocin, carnocyclin A, forms anion-selective channels in lipid bilayers. Biochim. Biophys. Acta 2009, 1788, 1797–1803. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Barrena, M.; Martínez-Ripoll, G.; Gálvez, A.; Valdivia, E.; Maqueda, M.; Cruz, V.; Albert, A. Structure of bacteriocin AS-48: From soluble state to membrane bound state. J. Mol. Biol. 2003, 334, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D. An “Upp”-turn in bacteriocin receptor identification. Mol. Microbiol. 2014, 92, 1159–1163. [Google Scholar] [PubMed]
- Bierbaum, G.; Sahl, H.G. Induction of autolysis of staphylococci by the basic peptide antibiotics Pep 5 and nisin and their influence on the activity of autolytic enzymes. Arch. Microbiol. 1985, 141, 249–254. [Google Scholar] [PubMed]
- Mora, D.; Musacchio, F.; Fortina, M.G.; Senini, L.; Manachini, P.L. Autolytic activity and pediocin-induced lysis in Pediococcus acidilactici and Pediococcus pentosaceus strains. J. Appl. Microbiol. 2003, 94, 561–570. [Google Scholar] [PubMed]
- Vollmer, W.; Joris, B.; Charlier, P.; Foster, S. Bacterial peptidoglycan (murein) hydrolases. FEMS Microbiol. Rev. 2008, 32, 259–286. [Google Scholar] [CrossRef] [PubMed]
- Grande Burgos, M.J.; Kovács, A.T.; Mirończuk, A.M.; Abriouel, H.; Gálvez, A.; Kuipers, O.P. Response of Bacillus cereus ATCC 14579 to challenges with sublethal concentrations of enterocin AS-48. BMC Microbiol. 2009, 28, 227. [Google Scholar] [CrossRef]
- Abriouel, H.; Lucas, R.; Ben Omar, N.; Valdivia, E.; Gálvez, A. Potential applications of the cyclic peptide enterocin AS-48 in the preservation of vegetable foods and beverages. Probiot. Antimicrob. Prot. 2010, 2, 77–89. [Google Scholar] [CrossRef]
- Khan, H.; Flint, S.; Yu, P.-L. Enterocins in food preservation. Int. J. Food Microbiol. 2010, 141, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ananou, S.; Garriga, M.; Hugas, M.; Maqueda, M.; Martínez-Bueno, M.; Gálvez, A.; Valdivia, E. Control of Listeria monocytogenes in model sausages by enterocin AS-48. Int. J. Food Microbiol. 2005, 103, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Ananou, S.; Maqueda, M.; Martínez-Bueno, M.; Gálvez, A.; Valdivia, E. Control of Staphylococcus aureus in sausages by enterocin AS-48. Meat Sci. 2005, 71, 549–576. [Google Scholar] [CrossRef] [PubMed]
- Ananou, S.; Baños, A.; Maqueda, M.; Martínez-Bueno, M.; Gálvez, A.; Valdivia, E. Effect of combined physico-chemical treatments based on enterocin AS-48 on the control of Listeria monocytogenes and Staphylococcus aureus in a model cooked ham. Food Control 2009, 21, 478–486. [Google Scholar] [CrossRef]
- Ananou, S.; Garriga, M.; Jofré, A.; Aymerich, T.; Gálvez, A.; Maqueda, M.; Martínez-Bueno, M.; Valdivia, E. Combined effect of enterocin AS-48 and high hydrostatic pressure to control food-borne pathogens inoculated in low acid fermented sausages. Meat Sci. 2010, 84, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Baños, A.; Ananou, S.; Martínez-Bueno, M.; Gálvez, A.; Maqueda, M.; Valdivia, E. Prevention of spoilage by enterocin AS-48 combined with chemical preservatives, under vacuum, or modified atmosphere in a cooked ham model. Food Control 2012, 24, 15–22. [Google Scholar] [CrossRef]
- Muñoz, A.; Maqueda, M.; Gálvez, A.; Martínez-Bueno, M.; Rodriguez, A.; Valdivia, E. Biocontrol of psychrotrophic enterotoxigenic Bacillus cereus in a non fat hard type cheese by an enterococcal strain-producing enterocin AS-48. J. Food Prot. 2004, 67, 1517–1521. [Google Scholar] [PubMed]
- Muñoz, A.; Ananou, S.; Gálvez, A.; Martínez-Bueno, M.; Rodríguez, A.; Maqueda, M.; Valdivia, E. Inhibition of Staphylococcus aureus in dairy products by enterocin AS-48 produced in situ and ex situ: Bactericidal synergism with heat. Int. Dairy J. 2007, 17, 760–769. [Google Scholar] [CrossRef]
- Sobrino, A.; Martínez Viedma, P.; Abriouel, H.; Valdivia, E.; Gálvez, A.; Martín Belloso, O. The impact of adding antimicrobial peptides to milk inoculated with Staphylococcus aureus and processed by high-intensity pulsed electric field. J. Dairy Sci. 2009, 92, 2514–2523. [Google Scholar] [CrossRef] [PubMed]
- Ananou, S.; Zentar, H.; Martínez-Bueno, M.; Gálvez, A.; Maqueda, M.; Valdivia, E. The impact of enterocin AS-48 on the shelf-life and safety of sardines (Sardina pilchardus) under different storage conditions. Food Microbiol. 2014, 44, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Cobo Molinos, A.; Abriouel, H.; Ben Omar, N.; Valdivia, E.; Lucas, R.; Maqueda, M.; Martínez Cañamero, M.; Gálvez, A. Effect of immersion solutions containing enterocin AS-48 on Listeria monocytogenes in vegetable foods. Appl. Environ. Microbiol. 2005, 71, 7781–7787. [Google Scholar] [CrossRef] [PubMed]
- Cobo Molinos, A.; Abriouel, H.; Ben Omar, N.; Lucas, R.; Valdivia, E.; Gálvez, A. Inactivation of Listeria monocytogenes in raw fruits by enterocin AS-48. J. Food Prot. 2008, 71, 2460–2467. [Google Scholar]
- Cobo Molinos, A.; Abriouel, H.; Lucas, R.; Ben Omar, N.; Valdivia, E.; Gálvez, A. Inhibition of Bacillus cereus and B. weihenstephanensis in raw vegetables by application of washing solutions containing enterocin AS-48 alone and in combination with other antimicrobials. Food Microbiol. 2008, 25, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Cobo Molinos, A.; Abriouel, H.; Lucas, R.; Valdivia, E.; Ben Omar, N.; Gálvez, A. Combined physico-chemical treatments based on enterocin AS-48 for inactivation of gram-negative bacteria in soybean sprouts. Food Chem. Toxicol. 2008, 46, 2912–2921. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.P.; Erickson, M.C. Summer meeting 2007—The problems with fresh produce: An overview. J. Appl. Microbiol. 2008, 105, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Cobo Molinos, A.; Abriouel, H.; Ben Omar, N.; Lucas, R.; Valdivia, E.; Gálvez, A. Enhanced bactericidal activity of enterocin AS-48 in combination with essential oils, natural bioactive compounds, and chemical preservatives against Listeria monocytogenes in ready-to-eat salads. Food Chem. Toxicol. 2009, 47, 2216–2223. [Google Scholar] [CrossRef] [PubMed]
- Cobo Molinos, A.; Lucas, R.; Abriouel, H.; Ben Omar, N.; Valdivia, E.; Gálvez, A. Inhibition of Salmonella enterica cells in deli-type salad by enterocin AS-48 in combination with other antimicrobials. Probiot. Antimicrob. Prot. 2009, 1, 85–90. [Google Scholar]
- Grande, M.J.; Lucas, R.; Abriouel, H.; Valdivia, E.; Ben Omar, N.; Maqueda, M.; Martínez-Cañamero, M.; Gálvez, A. Treatment of vegetable sauces with enterocin AS-48 alone or in combination with phenolic compounds to inhibit proliferation of Staphylococcus aureus. J. Food Prot. 2007, 70, 405–411. [Google Scholar] [PubMed]
- Martínez-Viedma, P.; Abriouel, H.; Ben Omar, N.; Valdivia, E.; Lucas López, R.; Gálvez, A. Inactivation of exopolysaccharide and 3- hydroxypropionaldehyde-producing lactic acid bacteria in apple juice and apple cider by enterocin AS-48. Food Chem. Toxicol. 2008, 46, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Viedma, P.; Abriouel, H.; Sobrino, A.; Ben Omar, N.; Lucas López, R.; Valdivia, E.; Martín Belloso, O.; Gálvez, A. Effect of enterocin AS-48 in combination with High-Intensity Pulsed-Electric Field treatment against the spoilage bacterium Lactobacillus diolivorans in apple juice. Food Microbiol. 2009, 26, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Viedma, P.; Sobrino, A.; Abriouel, H.; Ben Omar, N.; Lucas López, R.; Martín Belloso, O.; Gálvez, A. Increased inactivation of exopolysaccharide-producing Pediococcus parvulus in apple juice by combined treatment with enterocin AS-48 and high-intensity pulsed-electric field. J. Food Prot. 2009, 73, 39–43. [Google Scholar]
- Martínez-Viedma, P.; Abriouel, H.; Ben Omar, N.; Lucas López, R.; Valdivia, E.; Gálvez, A. Antibacterial protection by enterocin AS-48 in sport and energy drinks with less acidic pH values. J. Food Prot. 2009, 72, 881–884. [Google Scholar]
- Grande, M.J.; Abriouel, H.; Lucas, R.; Valdivia, E.; Ben Omar, N.; Martínez-Cañamero, M.; Gálvez, A. Efficacy of enterocin AS-48 against bacilli in ready-to-eat vegetable soups and purees. J. Food Prot. 2007, 70, 2339–2345. [Google Scholar] [PubMed]
- Lucas, R.; Grande, M.J.; Abriouel, H.; Maqueda, M.; Ben Omar, N.; Valdivia, E.; Martínez-Cañamero, M.; Gálvez, A. Application of the broad-spectrum bacteriocin enterocin AS-48 to inhibit Bacillus coagulans in low-pH canned fruit and vegetable foods. Food Chem. Toxicol. 2006, 44, 1774–1781. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Viedma, P.; Abriouel, H.; Ben Omar, N.; Lucas, R.; Gálvez, A. Inhibition of spoilage and toxinogenic Bacillus species in dough from wheat flour by the cyclic peptide enterocin AS-48. Food Control 2011, 22, 756–761. [Google Scholar] [CrossRef]
- Martínez-Viedma, P.; Abriouel, H.; Ben Omar, N.; Lucas, R.; Gálvez, A. Antistaphylococcal effect of enterocin AS-48 in bakery ingredients of vegetable origin, alone and in combination with selected antimicrobials. J. Food Sci. 2009, 74, M384–M389. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Viedma, P.; Abriouel, H.; Ben Omar, N.; Lucas, R.; Valdivia, E.; Gálvez, A. Assay of enterocin AS-48 for inhibition of foodborne pathogens in desserts. J. Food Prot. 2009, 72, 1654–1659. [Google Scholar] [PubMed]
- Montalbán-López, M.; Sánchez-Hidalgo, M.; Valdivia, E.; Martínez-Bueno, M.; Maqueda, M. Are bacteriocins underexploited? Novel applications for old antimicrobials. Curr. Pharm. Biotechnol. 2011, 12, 1205–1220. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grande Burgos, M.J.; Pulido, R.P.; Del Carmen López Aguayo, M.; Gálvez, A.; Lucas, R. The Cyclic Antibacterial Peptide Enterocin AS-48: Isolation, Mode of Action, and Possible Food Applications. Int. J. Mol. Sci. 2014, 15, 22706-22727. https://doi.org/10.3390/ijms151222706
Grande Burgos MJ, Pulido RP, Del Carmen López Aguayo M, Gálvez A, Lucas R. The Cyclic Antibacterial Peptide Enterocin AS-48: Isolation, Mode of Action, and Possible Food Applications. International Journal of Molecular Sciences. 2014; 15(12):22706-22727. https://doi.org/10.3390/ijms151222706
Chicago/Turabian StyleGrande Burgos, María José, Rubén Pérez Pulido, María Del Carmen López Aguayo, Antonio Gálvez, and Rosario Lucas. 2014. "The Cyclic Antibacterial Peptide Enterocin AS-48: Isolation, Mode of Action, and Possible Food Applications" International Journal of Molecular Sciences 15, no. 12: 22706-22727. https://doi.org/10.3390/ijms151222706