Detection of Abnormal Extracellular Matrix in the Interstitium of Regenerating Renal Tubules

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
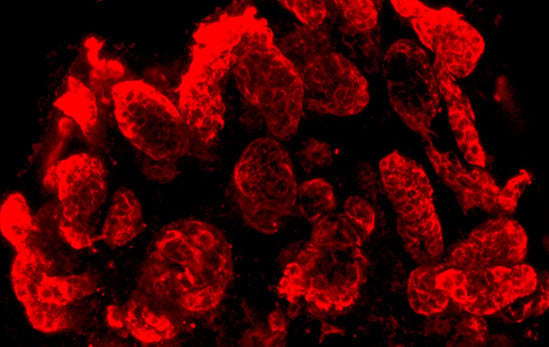
2.1. Extension of Tubules within an Artificial Interstitium

2.2. Extracellular Matrix in the Interstitium of Generated Tubules
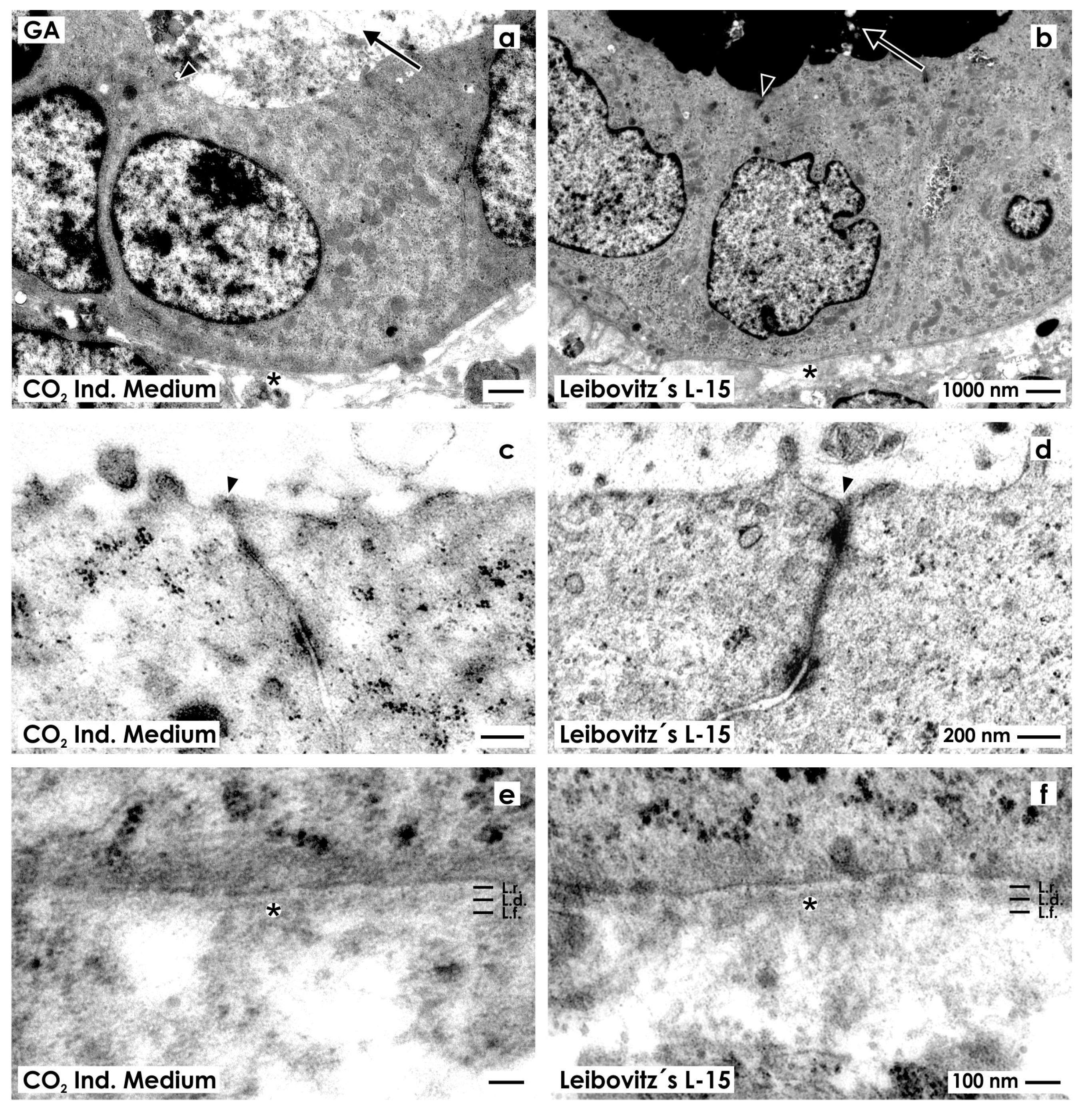
2.2.1. Fixation with Conventional Glutaraldehyde Solution

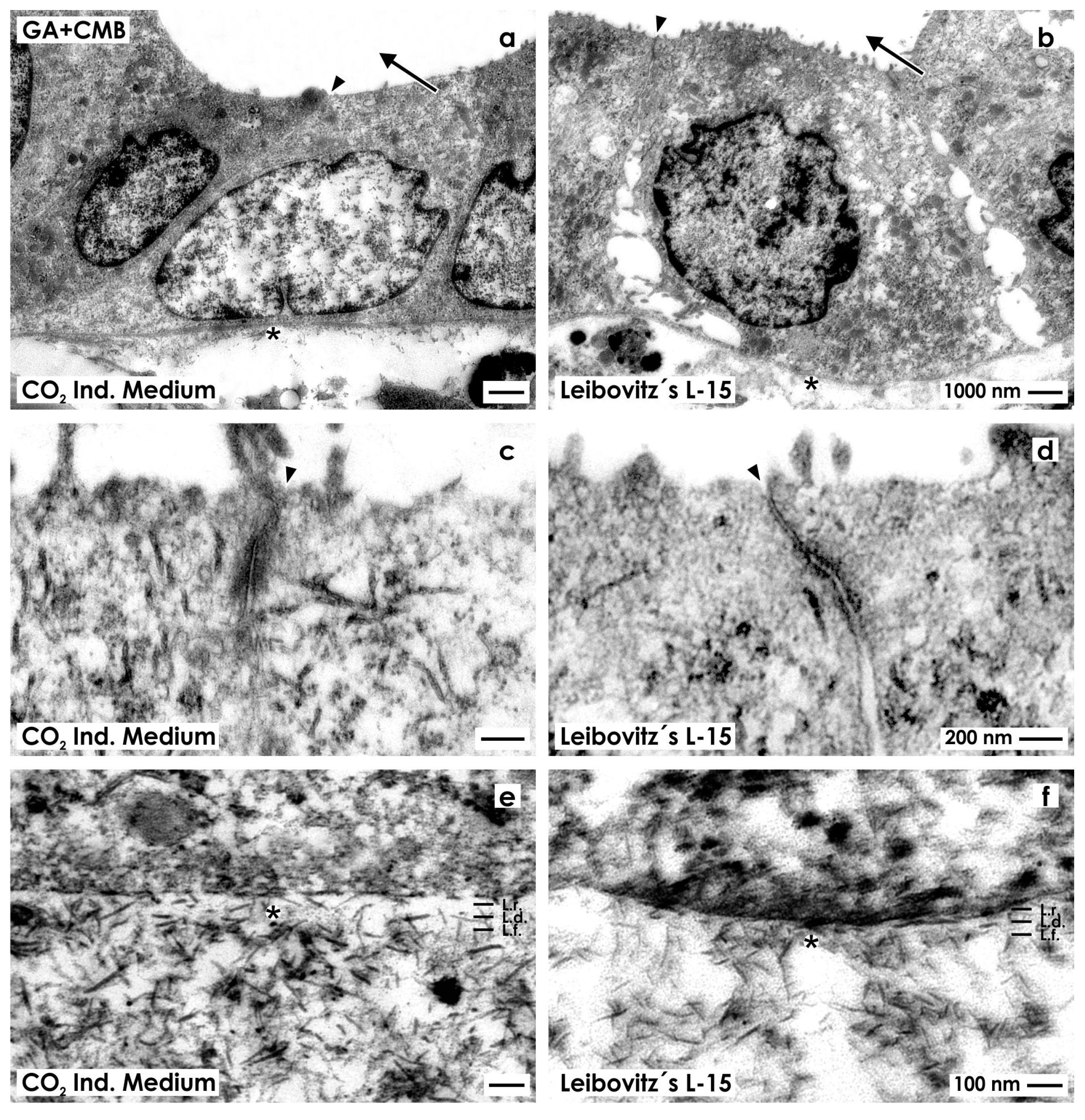
2.2.2. Fixation with Glutaraldehyde Containing Cupromeronic Blue

2.2.3. Fixation with Glutaraldehyde Containing Ruthenium Red

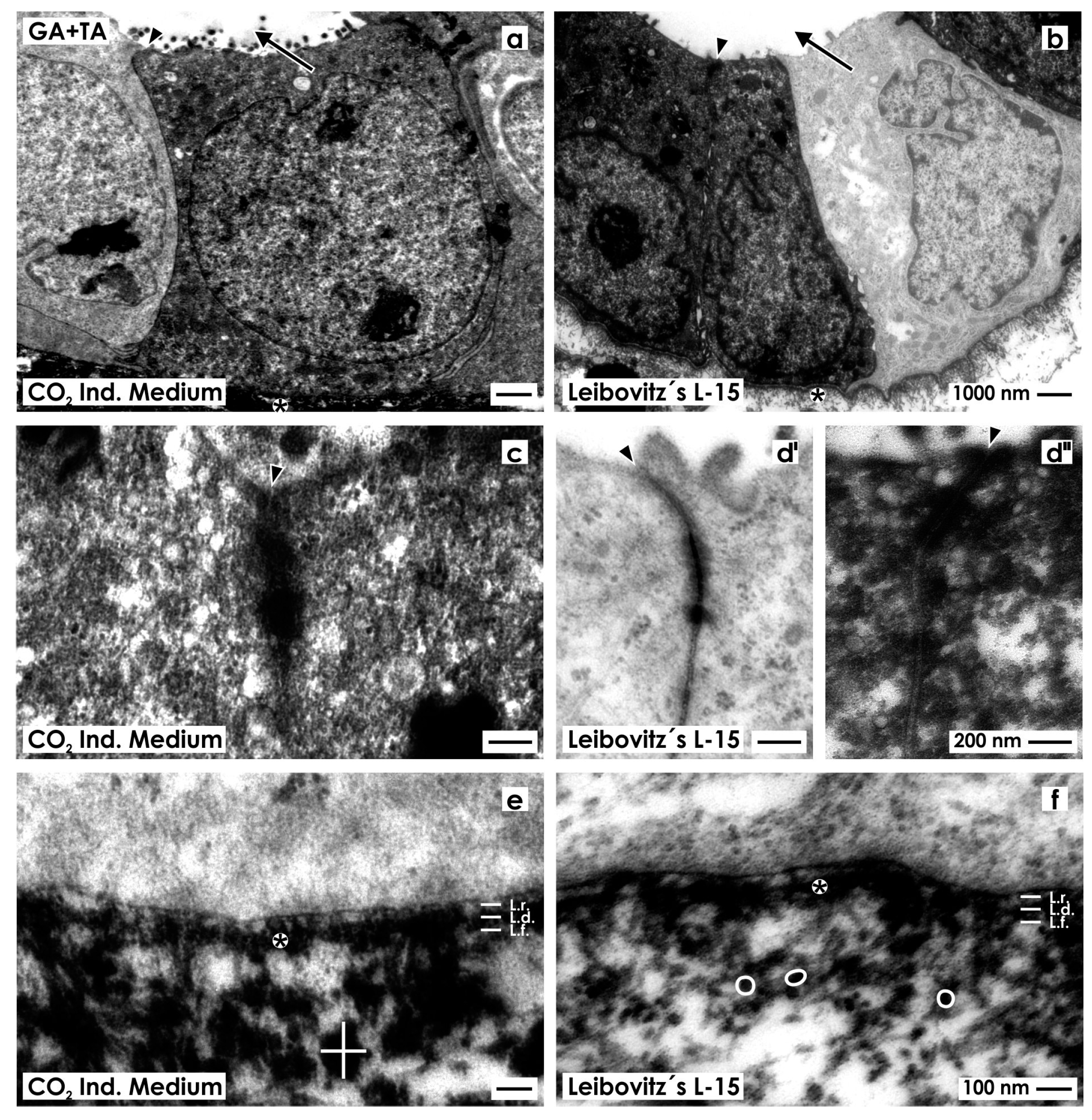
2.2.4. Fixation with Glutaraldehyde Containing Tannic Acid

2.2.5. Advanced Fixation in Matured Kidney Parenchyma versus Generated Tubules

2.3. Unveiling of Extracellular Matrix by Advanced Fixation
2.4. Recognition of Abnormal Features in Regenerating Tubules
3. Experimental Section
3.1. Isolation of Renal Stem/Progenitor Cells
3.2. Artificial Interstitium for Perfusion Culture

3.3. Histochemistry
3.4. Fixation and Embedding
- Series 1: 5% glutaraldehyde (Serva) buffered with 0.15 M sodium cacodylate, pH 7.4;
- Series 2: 5% glutaraldehyde buffered with 0.15 M sodium cacodylate, pH 7.4. Then incubation was performed with 0.1% cupromeronic blue (Santa Cruz, Heidelberg, Germany) and 0.1 M magnesium chloride hexahydrate (Sigma, Taufkirchen, Germany) in sodium acetate buffer, pH 5.6. Counterstain was performed with 0.5% sodium tungstate dehydrate (Sigma);
- Series 3: 5% glutaraldehyde buffered with 0.15 M sodium cacodylate, pH 7.4 containing 0.5% ruthenium red (Fluka);
- Series 4: 5% glutaraldehyde buffered with 0.15 M sodium cacodylate, pH 7.4 containing 1% tannic acid (Sigma).
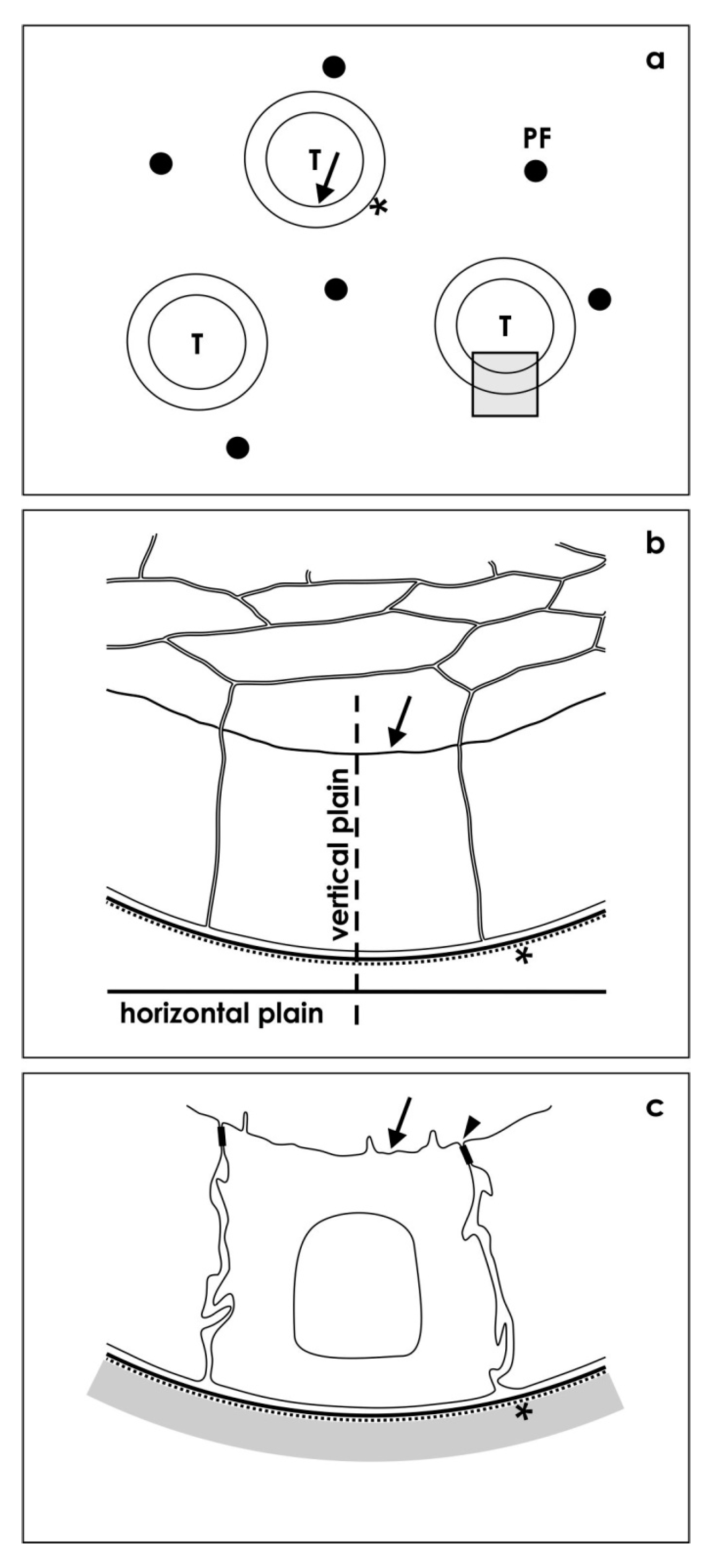
3.5. Positioning of Tissue for Morphological Analysis
3.6. Transmission Electron Microscopy (TEM)

3.7. Amount of Analyzed Specimens
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Morigi, M.; de Coppi, P. Cell Therapy for kidney injury: different options and mechanisms—Mesenchymal and amniotic fluid stem cells. Nephron. Exp. Nephrol. 2014, 126, s59–63. [Google Scholar]
- Winyard, P.J.D.; Rice, K.L. Experimental renal progenitor cells: repairing and creating kidneys? Pediatr. Nephrol. 2014, 29, 665–672. [Google Scholar]
- Maeshima, A.; Nakasatomi, M.; Nojima, Y. Regenerative medicine for the kidney: Renotropic factors, renal stem/progenitor cells, and stem cell therapy. BioMed Res. Int. 2014, 2014, 595493. [Google Scholar]
- Herrera, M.; Mirotsu, M. Stem cells: Potential and callenges for kidney repair. Am. J. Physiol. Renal. Physiol. 2014, 306, F12–F23. [Google Scholar]
- Burst, V.; Pütsch, F.; Kubacki, T.; Völker, L.A.; Bartram, M.P.; Müller, R.U.; Gillis, M.; Kurschat, C.E.; Grundmann, F.; Müller-Ehmsen, J.; et al. Survival and distribution of injected haematopoietic stem cells in acute kidney injury. Nephrol. Dial. Transplant. 2013, 28, 1131–1139. [Google Scholar]
- Wang, Y.; He, J.; Zhao, W. Systematic review and meta-analysis of mesenchymal stem/stromal cells therapy for impaired renal function in small animal models. Nephrology 2013, 18, 201–208. [Google Scholar]
- Ricardo, S.D.; van Goor, H.; Eddy, A.A. Macrophage diversity in renal injury and repair. J. Clin. Investig. 2008, 118, 3522–3530. [Google Scholar]
- Zoja, C; Garcia, P.B.; Remuzzi, G. The role of chemokines in progressive renal disease. Front. Biosci. 2009, 14, 1815–1822. [Google Scholar]
- Minuth, W.W.; Denk, L.; Gruber, M. Search for chemically defined culture medium to assist initial regeneration of diseased renal parenchyma after stem/progenitor cell implantation. Int. J. Stem Cell Res. Transplant. 2013, 1, 202. [Google Scholar]
- Roessger, A.; Denk, L.; Minuth, W.W. Potential of stem/progenitor cell cultures within polyester fleeces to regenerate renal tubules. Biomaterials 2009, 30, 3723–3732. [Google Scholar]
- Minuth, W.W.; Denk, L. Interstitial interfaces show marked differences in regenerating tubules, matured tubules, and the renal stem/progenitor cell niche. J. Biomed. Mater. Res. A 2012, 100A, 1115–1125. [Google Scholar]
- Minuth, W.W.; Denk, L. Initial steps to stabilize the microenvironment for implantation of stem/progenitor cells in diseased renal parenchyma. Transplant. Technol. 2013, 1, 2. [Google Scholar]
- Minuth, W.W.; Denk, L. Tannic acid label indicates abnormal cell development coinciding with regeneration of renal tubules. BMC Clin. Pathol. 2014, 14, 34. [Google Scholar]
- Gallegos, T.F.; Kouznetsova, V.; Kudlicka, K.; Sweeney, D.E.; Bush, K.T.; Willert, K.; Farquhar, M.G.; Nigam, S.K. A protein kinase A and Wnt-dependent network regulating an intermediate stage in epithelial tubulogenesis during kidney development. Dev. Biol. 2012, 364, 11–21. [Google Scholar]
- Liu, B.; Li, C.; Liu, Z.; Dai, Z.; Tao, Y. Increasing extracellular matrix collagen level and MMM activity induces cyst development in polycystic kidney disease. BMC Nephrol. 2012, 13, 109. [Google Scholar]
- Kumar, S.; Liu, J.; McMahon, A.P. Defining the acute kidney injury and repair transcriptome. Semin. Nephrol. 2014, 34, 404–417. [Google Scholar]
- Baulier, E.; Favreau, F.; le Corf, A.; Jayle, C.; Schneider, F.; Goujon, J.M.; Feraud, O.; Bennaceur-Griscelli, A.; Hauet, T.; Turhan, A.G. Amniotic fluid-derived mesenchymal stem cells prevent fibrosis and preserve renal function in a preclinical porcine model of kidney transplantation. Stem Cells Transl. Med. 2014, 3, 809–820. [Google Scholar]
- Li, W.; Hartwig, S.; Rosenblum, N.D. Develomental origins and functions of stromal cells in the normal and diseased mammalian kidney. Dev. Dyn. 2014, 243, 853–863. [Google Scholar]
- Heber, S.; Denk, L.; Hu, K.; Minuth, W.W. Modulating the development of renal tubules growing in serum-free culture medium at an artificial interstitium. Tissue Eng. 2007, 13, 281–292. [Google Scholar]
- Minuth, W.W.; Denk, L.; Glashauser, A. Promoting and harmful effects of steroid hormones on renal stem/progenitor cell devevelopment. J. Tissue Sci. Eng. 2010, 1, 101. [Google Scholar]
- Minuth, W.W.; Denk, L.; Meese, C.; Rachel, R.; Roessger, A. Ultrastructural insights in the interface between generated renal tubules and a polyester interstitium. Langmuir 2009, 25, 4621–4627. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minuth, W.W.; Denk, L. Detection of Abnormal Extracellular Matrix in the Interstitium of Regenerating Renal Tubules. Int. J. Mol. Sci. 2014, 15, 23240-23254. https://doi.org/10.3390/ijms151223240
Minuth WW, Denk L. Detection of Abnormal Extracellular Matrix in the Interstitium of Regenerating Renal Tubules. International Journal of Molecular Sciences. 2014; 15(12):23240-23254. https://doi.org/10.3390/ijms151223240
Chicago/Turabian StyleMinuth, Will W., and Lucia Denk. 2014. "Detection of Abnormal Extracellular Matrix in the Interstitium of Regenerating Renal Tubules" International Journal of Molecular Sciences 15, no. 12: 23240-23254. https://doi.org/10.3390/ijms151223240