A Caspase-Dependent Pathway Is Involved in Wnt/β-Catenin Signaling Promoted Apoptosis in Bacillus Calmette-Guerin Infected RAW264.7 Macrophages

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
2.1.1. Bacillus Calmette-Guerin (BCG) Activated Canonical Wnt Signaling in Macrophage RAW264.7 Cells
2.1.2. Wnt/β-Catenin Signaling Down-Regulates BCG-Induced IL-6 and TNF-α Production in RAW264.7 Cells
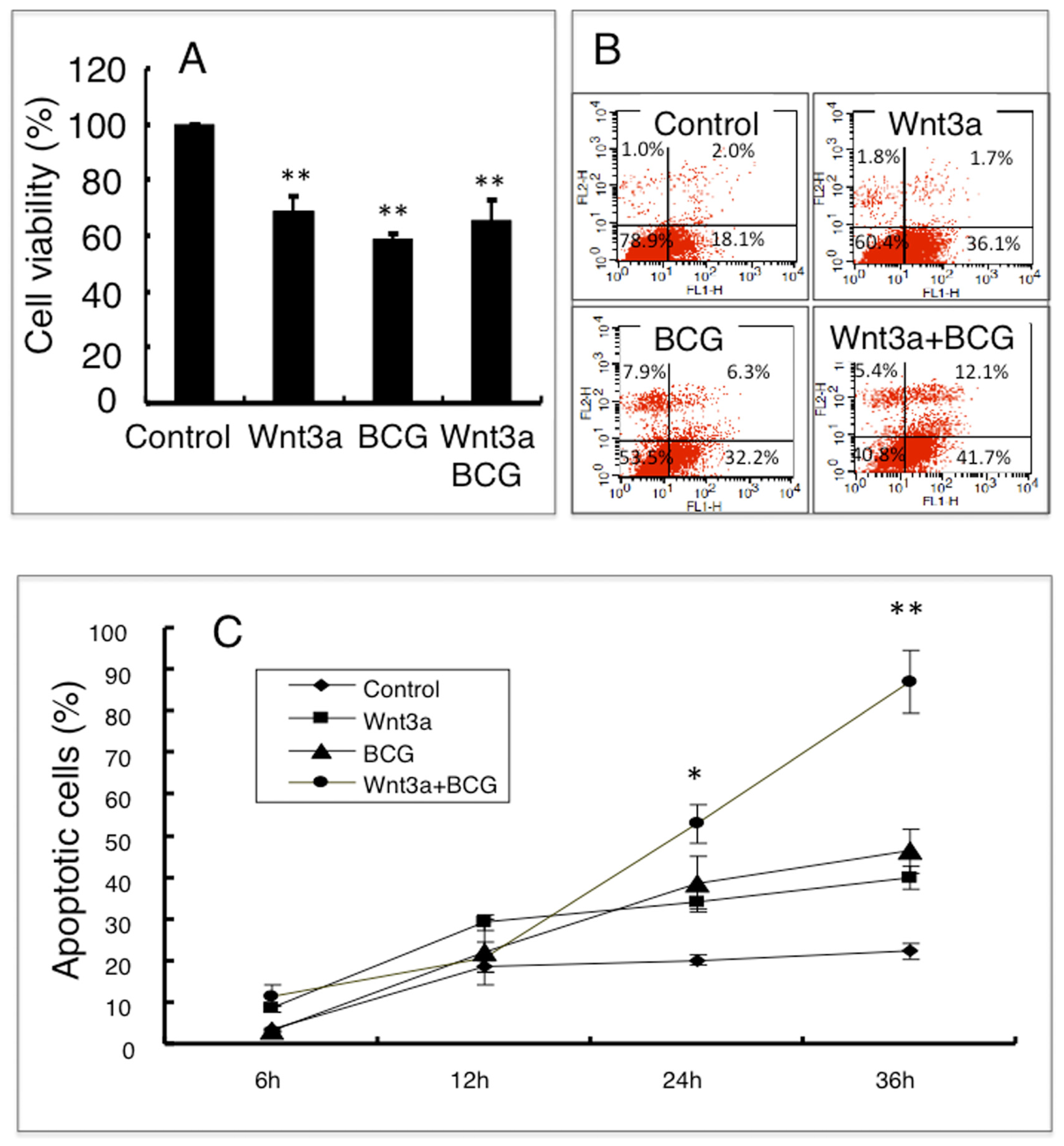
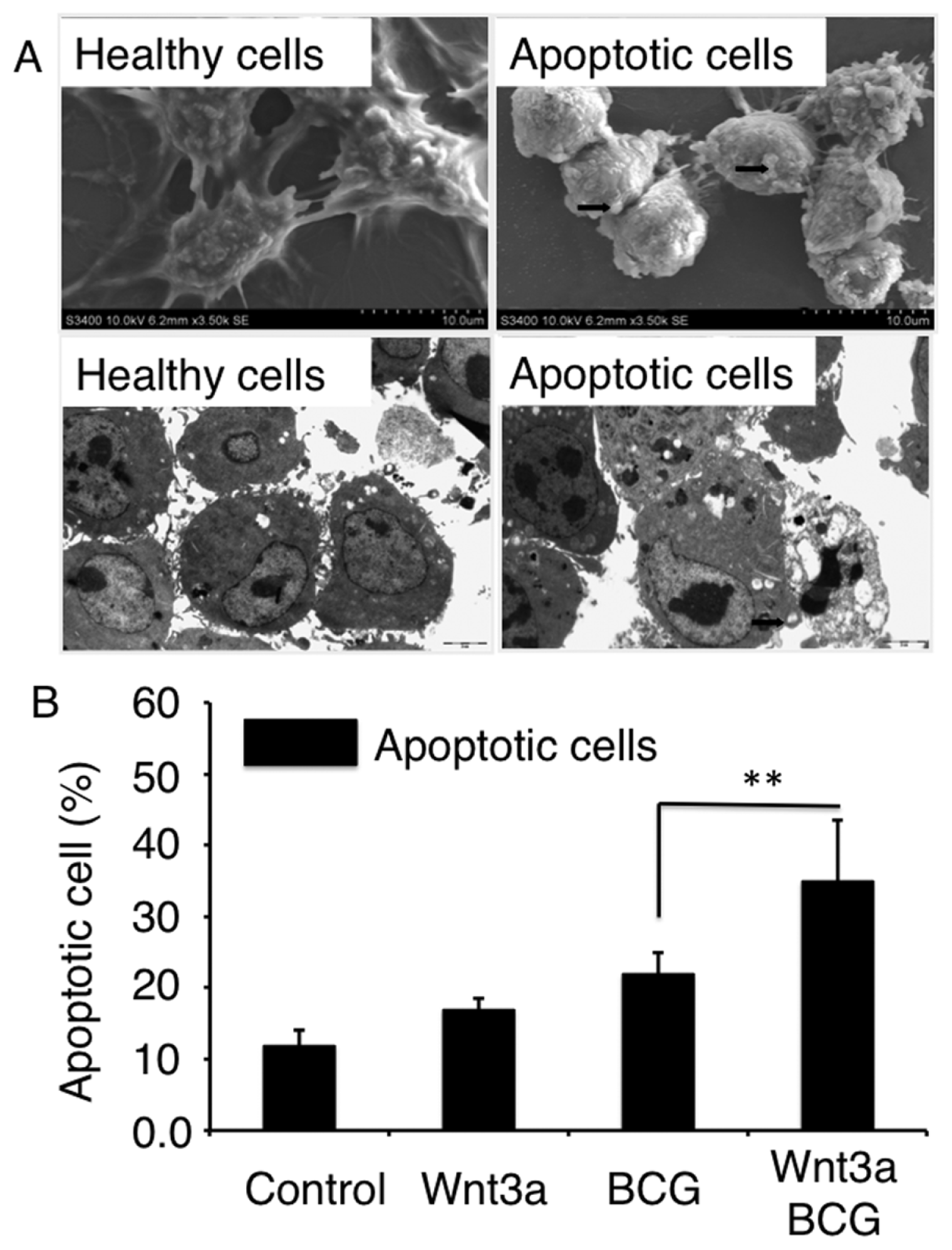
2.1.3. Wnt3a Promotes Apoptosis in RAW264.7 Cells Infected with BCG
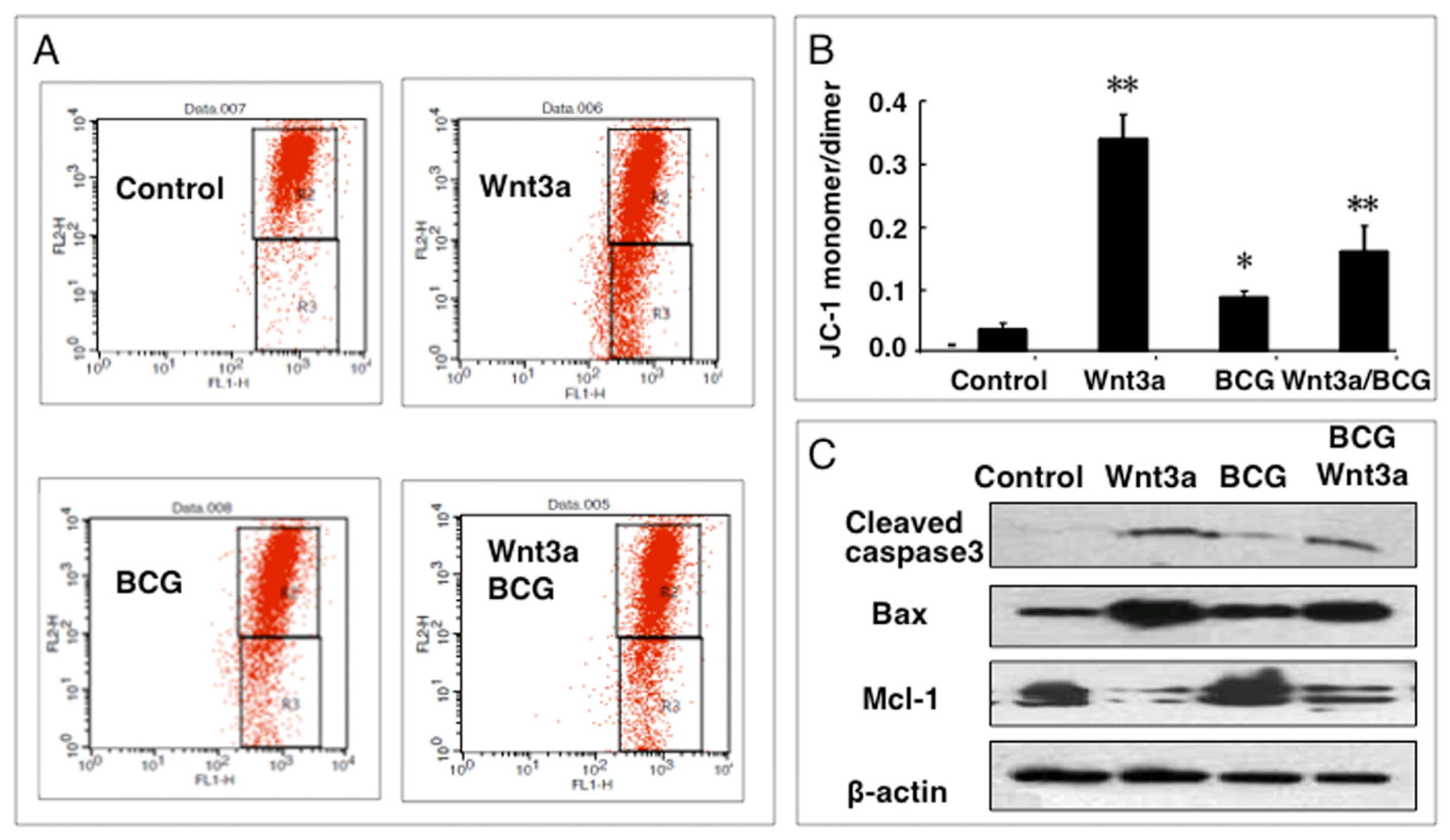
2.1.4. Wnt3a Affects Mitochondrial Membrane Potential (ΔΨm) in RAW264.7 Cells
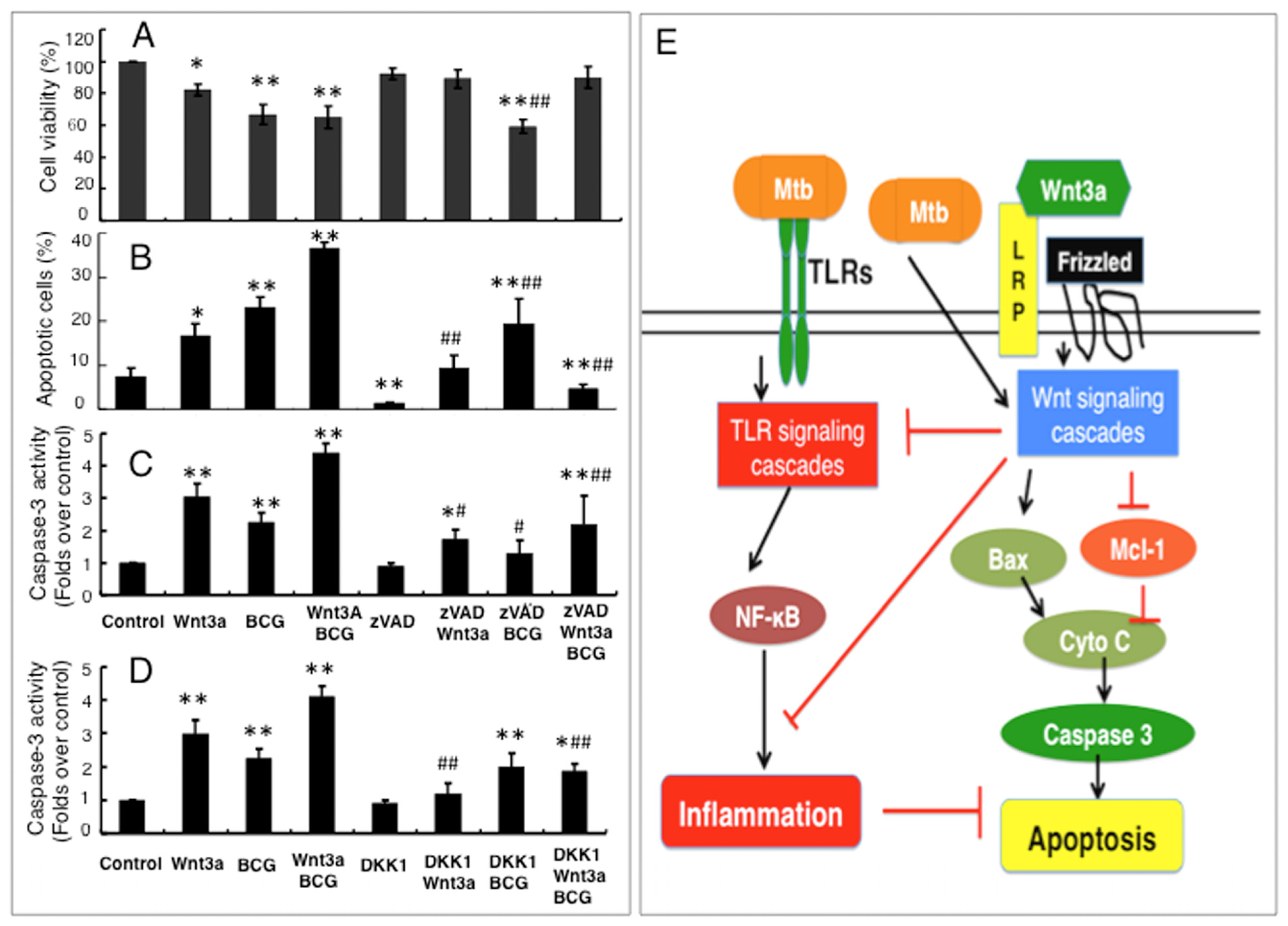
2.1.5. Wnt3a Promoted BCG-Infected RAW264.7 Cell Apoptosis in Part through a Caspase-Dependent Mechanism
2.2. Discussion
3. Experimental Section
3.1. Cell Lines and Wnt3a Conditioned Medium
3.2. Plasmids and Transfection
3.3. Infection of RAW264.7 Macrophage Cells with BCG
3.4. Dual-Luciferase Reporter Assay
3.5. Quantitative Reverse Transcription PCR (qRT-PCR)
3.6. Enzyme-Linked Immunosorbent Assay (ELISA)
3.7. MTT Assay
3.8. Flow Cytometry Analysis for Cell Apoptosis
3.9. Electronic Microscopy
3.10. Mitochondrial Membrane Potential (ΔΨm) Assay
3.11. Immnoblotting Analysis
3.12. Caspase-3 Activity Assay
3.13. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsXW, XL and YW conceived and designed the experiments; XW, GD, XG, JZ, CM and YH performed the experiments; CM, YL, XW analyzed the data; XW, XL and YW wrote the paper.
References
- Russell, D.G. Mycobacterium tuberculosis and the intimate discourse of a chronic infection. Immunol. Rev. 2011, 240, 252–268. [Google Scholar]
- Gengenbacher, M.; Kaufmann, S.H. Mycobacterium tuberculosis: Success through dormancy. FEMS Microbiol. Rev. 2012, 36, 514–532. [Google Scholar]
- Zimmerman, Z.F.; Kulikauskas, R.M.; Bomsztyk, K.; Moon, R.T.; Chien, A.J. Activation of wnt/beta-catenin signaling increases apoptosis in melanoma cells treated with trail. PLoS One 2013, 8, e69593. [Google Scholar]
- Lee, J.; Hartman, M.; Kornfeld, H. Macrophage apoptosis in tuberculosis. Yonsei Med. J. 2009, 50, 1–11. [Google Scholar]
- Butler, R.E.; Brodin, P.; Jang, J.; Jang, M.S.; Robertson, B.D.; Gicquel, B.; Stewart, G.R. The balance of apoptotic and necrotic cell death in Mycobacterium tuberculosis infected macrophages is not dependent on bacterial virulence. PLoS One 2012, 7, e47573. [Google Scholar]
- Kelly, D.M.; ten Bokum, A.M.; O’Leary, S.M.; O’Sullivan, M.P.; Keane, J. Bystander macrophage apoptosis after Mycobacterium tuberculosis H37Ra infection. Infect. Immun. 2008, 76, 351–360. [Google Scholar]
- Cardona, P.-J.; Lee, J.; Repasy, T.; Papavinasasundaram, K.; Sassetti, C.; Kornfeld, H. Mycobacterium tuberculosis induces an atypical cell death mode to escape from infected macrophages. PLoS One 2011, 6, e18367. [Google Scholar]
- Neumann, J.; Schaale, K.; Farhat, K.; Endermann, T.; Ulmer, A.J.; Ehlers, S.; Reiling, N. Frizzled1 is a marker of inflammatory macrophages and its ligand Wnt3a is involved in reprogramming Mycobacterium tuberculosis-infected macrophages. FASEB J. 2010, 24, 4599–4612. [Google Scholar]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar]
- Staal, F.J.; Clevers, H. Tcf/Lef transcription factors during T-cell development: Unique and overlapping functions. Hematol. J. 2000, 1, 3–6. [Google Scholar]
- Staal, F.J.; Luis, T.C.; Tiemessen, M.M. WNT signalling in the immune system: WNT is spreading its wings. Nat. Rev. Immunol. 2008, 8, 581–593. [Google Scholar]
- Pereira, C.P.; Bachli, E.B.; Schoedon, G. The wnt pathway: A macrophage effector molecule that triggers inflammation. Curr. Atheroscler. Rep. 2009, 11, 236–242. [Google Scholar]
- Aumiller, V.; Balsara, N.; Wilhelm, J.; Gunther, A.; Konigshoff, M. WNT/beta-catenin signaling induces IL-1beta expression by alveolar epithelial cells in pulmonary fibrosis. Am. J. Respir. Cell Mol. Biol. 2013, 49, 96–104. [Google Scholar]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar]
- Masckauchan, T.N.; Shawber, C.J.; Funahashi, Y.; Li, C.M.; Kitajewski, J. Wnt/beta-catenin signaling induces proliferation survival and interleukin-8 in human endothelial cells. Angiogenesis 2005, 8, 43–51. [Google Scholar]
- Wang, L.J.; Bai, L.; Su, D.; Zhang, T.; Mao, Z.Y. Proinflammatory conditions promote hepatocellular carcinoma onset and progression via activation of Wnt and EGFR signaling pathways. Mol. Cell. Biochem. 2013, 381, 173–181. [Google Scholar]
- Li, W.; Tong, H.; Huang, X.; Wang, W.; Wu, H.; Lin, S. High levels of beta-catenin promote IFNgamma-induced apoptosis in hepatocellular carcinoma cells. Oncol. Lett. 2012, 4, 1092–1096. [Google Scholar]
- Blumenthal, A.; Ehlers, S.; Lauber, J.; Buer, J.; Lange, C.; Goldmann, T.; Heine, H.; Brandt, E.; Reiling, N. The Wingless homolog WNT5A and its receptor Frizzled-5 regulate inflammatory responses of human mononuclear cells induced by microbial stimulation. Blood 2006, 108, 965–973. [Google Scholar]
- Schaale, K.; Neumann, J.; Schneider, D.; Ehlers, S.; Reiling, N. Wnt signaling in macrophages: Augmenting and inhibiting mycobacteria-induced inflammatory responses. Eur. J. Cell Biol. 2011, 90, 553–559. [Google Scholar]
- Oderup, C.; LaJevic, M.; Butcher, E.C. Canonical and noncanonical Wnt proteins program dendritic cell responses for tolerance. J. Immunol. 2013, 190, 6126–6134. [Google Scholar]
- Ming, M.; Wang, S.; Wu, W.; Senyuk, V.; Le Beau, M.M.; Nucifora, G.; Qian, Z. Activation of Wnt/beta-catenin protein signaling induces mitochondria-mediated apoptosis in hematopoietic progenitor cells. J. Biol. Chem. 2012, 287, 22683–22690. [Google Scholar]
- Wong, M.H.; Rubinfeld, B.; Gordon, J.I. Effects of forced expression of an NH2-terminal truncated beta-Catenin on mouse intestinal epithelial homeostasis. J. Cell Biol. 1998, 141, 765–777. [Google Scholar]
- Kim, K.; Pang, K.M.; Evans, M.; Hay, E.D. Overexpression of beta-catenin induces apoptosis independent of its transactivation function with LEF-1 or the involvement of major G1 cell cycle regulators. Mol. Biol. Cell 2000, 11, 3509–3523. [Google Scholar]
- Reya, T.; Duncan, A.W.; Ailles, L.; Domen, J.; Scherer, D.C.; Willert, K.; Hintz, L.; Nusse, R.; Weissman, I.L. A role for Wnt signalling in self-renewal of haematopoietic stem cells. Nature 2003, 423, 409–414. [Google Scholar]
- Willert, K.; Brown, J.D.; Danenberg, E.; Duncan, A.W.; Weissman, I.L.; Reya, T.; Yates, J.R., 3rd; Nusse, R. Wnt proteins are lipid-modified and can act as stem cell growth factors. Nature 2003, 423, 448–452. [Google Scholar]
- Trowbridge, J.J.; Xenocostas, A.; Moon, R.T.; Bhatia, M. Glycogen synthase kinase-3 is an in vivo regulator of hematopoietic stem cell repopulation. Nat. Med. 2006, 12, 89–98. [Google Scholar]
- Kirstetter, P.; Anderson, K.; Porse, B.T.; Jacobsen, S.E.; Nerlov, C. Activation of the canonical Wnt pathway leads to loss of hematopoietic stem cell repopulation and multilineage differentiation block. Nat. Immunol. 2006, 7, 1048–1056. [Google Scholar]
- Scheller, M.; Huelsken, J.; Rosenbauer, F.; Taketo, M.M.; Birchmeier, W.; Tenen, D.G.; Leutz, A. Hematopoietic stem cell and multilineage defects generated by constitutive beta-catenin activation. Nat. Immunol. 2006, 7, 1037–1047. [Google Scholar]
- Thornberry, N.A.; Lazebnik, Y. Caspases: Enemies within. Science 1998, 281, 1312–1316. [Google Scholar]
- Li, Y.; Gao, Q.; Yin, G.; Ding, X.; Hao, J. WNT/beta-catenin-signaling pathway stimulates the proliferation of cultured adult human Sertoli cells via upregulation of C-myc expression. Reprod. Sci. 2012, 19, 1232–1240. [Google Scholar]
- Zhang, S.; Li, Y.; Wu, Y.; Shi, K.; Bing, L.; Hao, J. Wnt/beta-catenin signaling pathway upregulates c-Myc expression to promote cell proliferation of P19 teratocarcinoma cells. Anat. Rec. 2012, 295, 2104–2113. [Google Scholar]
- Ponce, D.P.; Yefi, R.; Cabello, P.; Maturana, J.L.; Niechi, I.; Silva, E.; Galindo, M.; Antonelli, M.; Marcelain, K.; Armisen, R.; et al. CK2 functionally interacts with AKT/PKB to promote the beta-catenin-dependent expression of survivin and enhance cell survival. Mol. Cell. Biochem. 2011, 356, 127–132. [Google Scholar]
- Perry, J.M.; He, X.C.; Sugimura, R.; Grindley, J.C.; Haug, J.S.; Ding, S.; Li, L. Cooperation between both Wnt/{beta}-catenin and PTEN/PI3K/Akt signaling promotes primitive hematopoietic stem cell self-renewal and expansion. Genes Dev. 2011, 25, 1928–1942. [Google Scholar]
- Anne, S.L.; Govek, E.E.; Ayrault, O.; Kim, J.H.; Zhu, X.; Murphy, D.A.; van Aelst, L.; Roussel, M.F.; Hatten, M.E. WNT3 inhibits cerebellar granule neuron progenitor proliferation and medulloblastoma formation via MAPK activation. PLoS One 2013, 8, e81769. [Google Scholar]
- Rello, S.; Stockert, J.C.; Moreno, V.; Gamez, A.; Pacheco, M.; Juarranz, A.; Canete, M.; Villanueva, A. Morphological criteria to distinguish cell death induced by apoptotic and necrotic treatments. Apoptosis: Int. J. Program. Cell Death 2005, 10, 201–208. [Google Scholar]
- Raju, I.; Kannan, K.; Abraham, E.C. FoxO3a Serves as a biomarker of oxidative stress in human lens epithelial cells under conditions of hyperglycemia. PLoS One 2013, 8, e67126. [Google Scholar]





© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, X.; Deng, G.; Hao, X.; Li, Y.; Zeng, J.; Ma, C.; He, Y.; Liu, X.; Wang, Y. A Caspase-Dependent Pathway Is Involved in Wnt/β-Catenin Signaling Promoted Apoptosis in Bacillus Calmette-Guerin Infected RAW264.7 Macrophages. Int. J. Mol. Sci. 2014, 15, 5045-5062. https://doi.org/10.3390/ijms15035045
Wu X, Deng G, Hao X, Li Y, Zeng J, Ma C, He Y, Liu X, Wang Y. A Caspase-Dependent Pathway Is Involved in Wnt/β-Catenin Signaling Promoted Apoptosis in Bacillus Calmette-Guerin Infected RAW264.7 Macrophages. International Journal of Molecular Sciences. 2014; 15(3):5045-5062. https://doi.org/10.3390/ijms15035045
Chicago/Turabian StyleWu, Xiaoling, Guangcun Deng, Xiujing Hao, Yong Li, Jin Zeng, Chunyan Ma, Yulong He, Xiaoming Liu, and Yujiong Wang. 2014. "A Caspase-Dependent Pathway Is Involved in Wnt/β-Catenin Signaling Promoted Apoptosis in Bacillus Calmette-Guerin Infected RAW264.7 Macrophages" International Journal of Molecular Sciences 15, no. 3: 5045-5062. https://doi.org/10.3390/ijms15035045