A Chrysanthemum Heat Shock Protein Confers Tolerance to Abiotic Stress

Abstract
:1. Introduction
2. Results and Discussion
2.1. The CgHSP70 Sequence
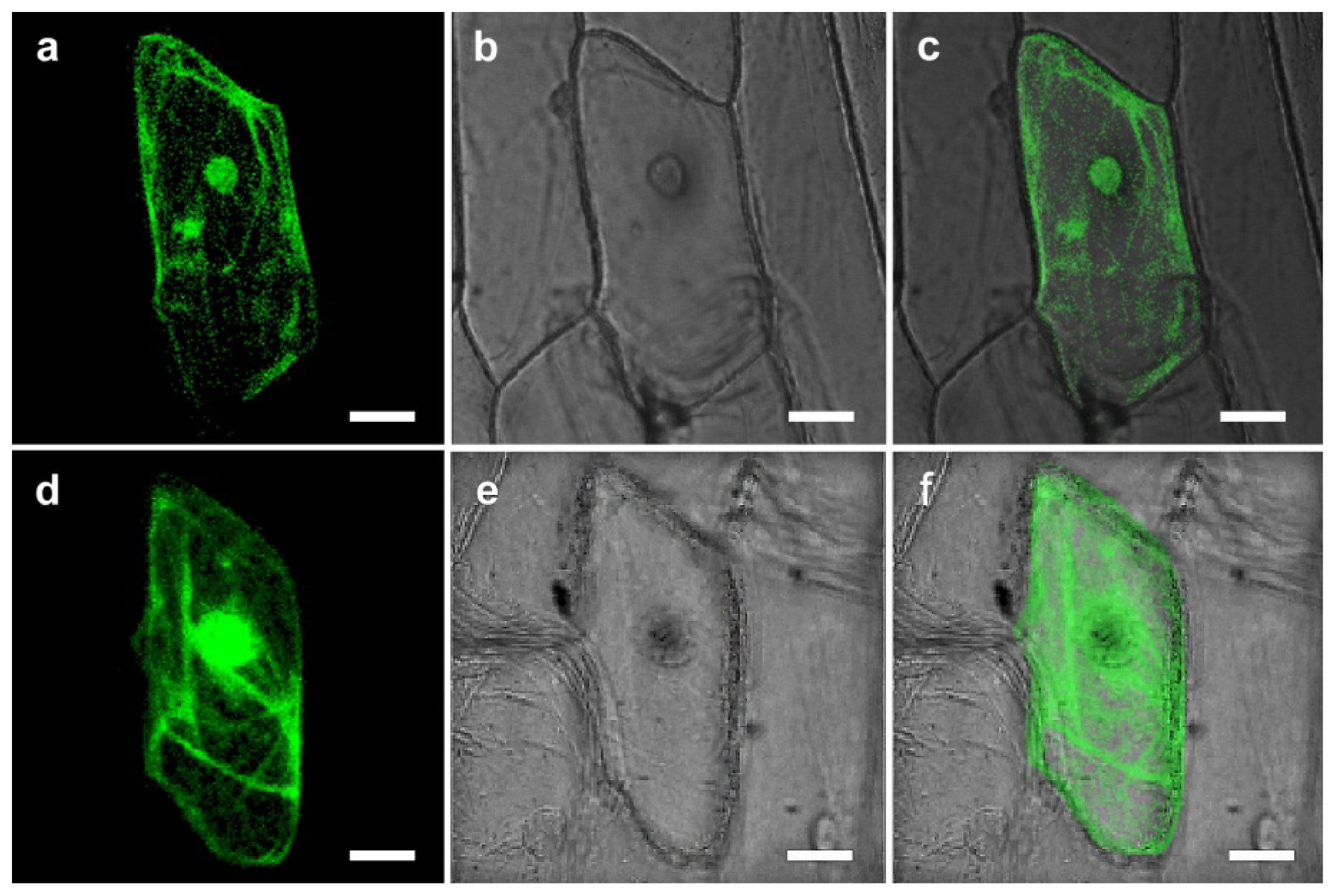
2.2. Sub-Cellular Localization of CgHSP70
2.3. Stress Tolerance of A. thaliana Plants Heterologously Expressing CgHSP70
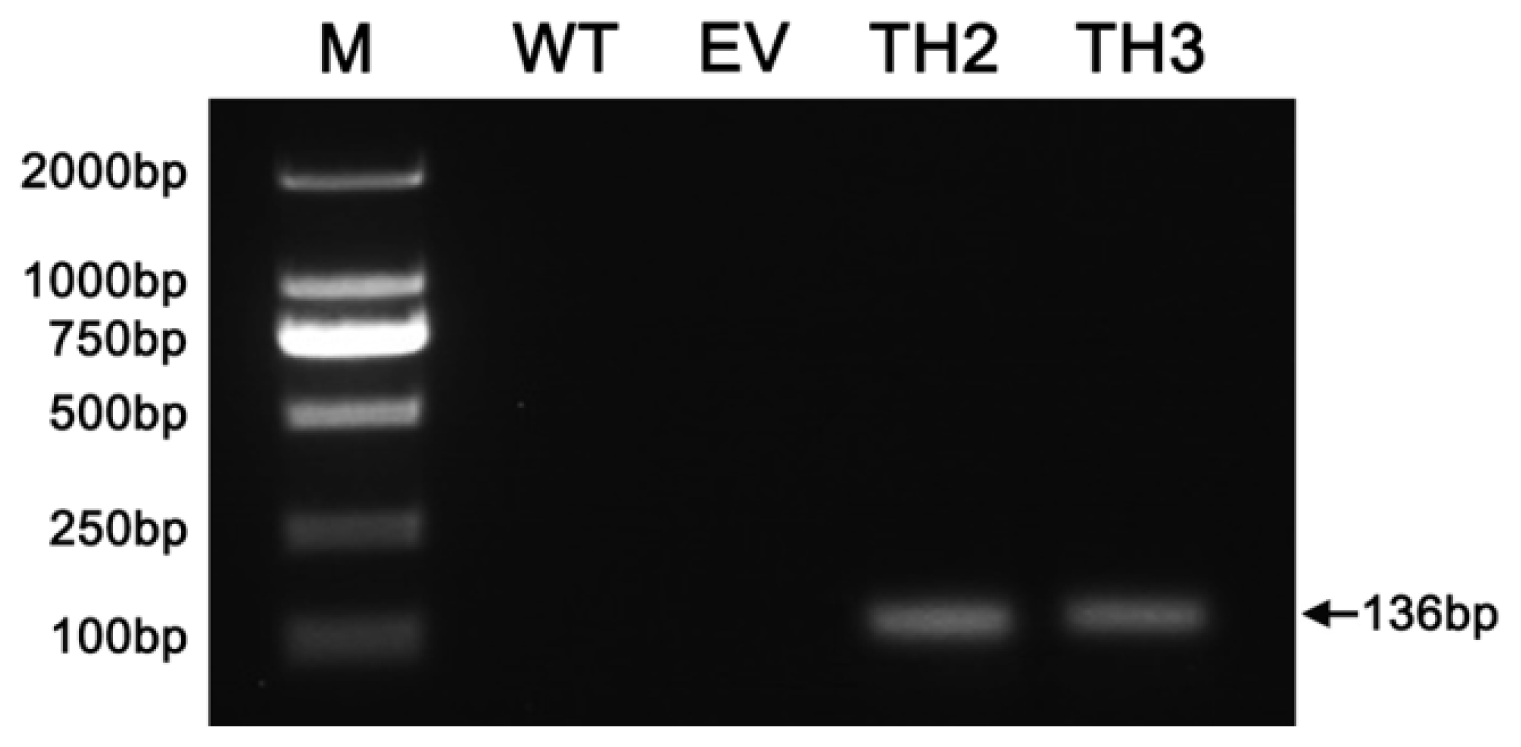
2.4. Transgene Transcription in Chrysanthemum
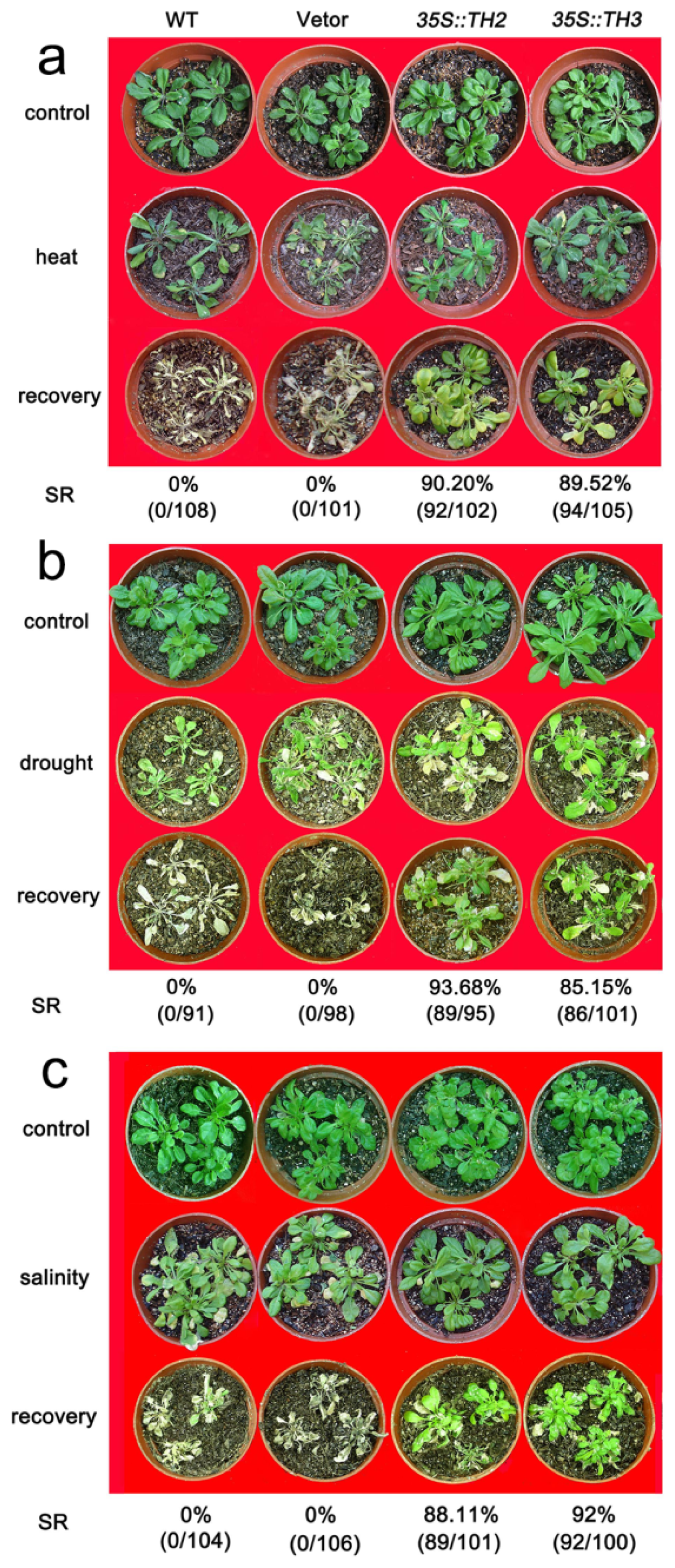
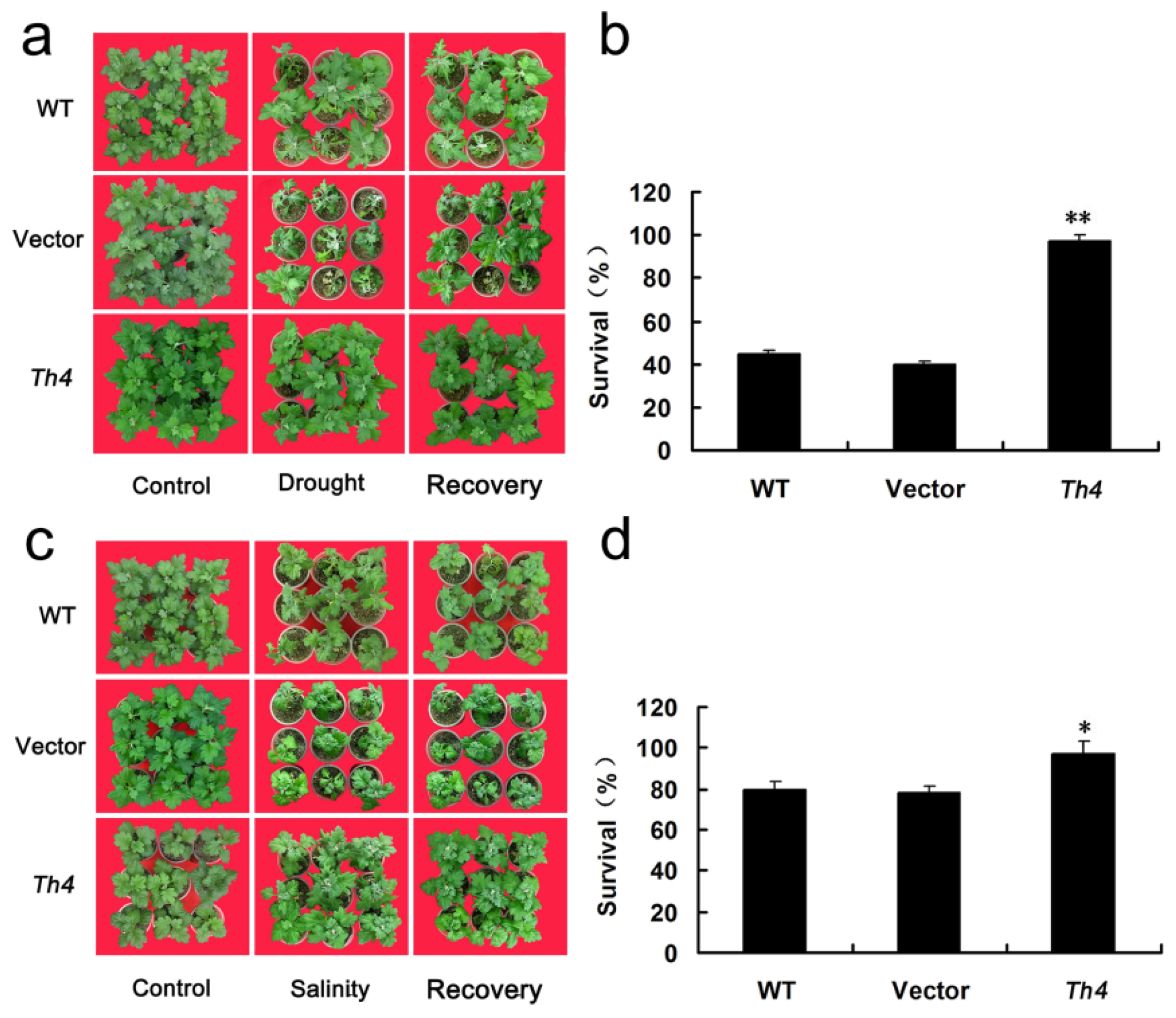
2.5. The Constitutive Expression of CgHSP70 Enhances the Abiotic Stress Tolerance of Chrysanthemum
2.6. Physiological Changes in CgHSP70 Over-Expression Chrysanthemum in Response to Abiotic Stresses
3. Experimental Section
3.1. Plant Materials
3.2. Isolation of CgHSP70
3.3. Intracellular Localization of CgHSP70
3.4. Transformation of A. thaliana
3.5. Stress Tolerance of Transgenic A. thaliana
3.6. Transformation of Chrysanthemum
3.7. Evaluation of the Abiotic Stress Tolerance of Transgenic Chrysanthemum
3.7.1. Heat Tolerance
3.7.2. Drought Tolerance
3.7.3. Salinity Tolerance
3.7.4. Measurement of Physiological Indices
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Schöffl, F.; Prandl, R.; Reindl, A. Molecular responses to heat stress. In Molecular Responses to Cold, Drought, Heat and Salt Stress in Higher Plants; RG Landes Co.: Austin, TX, USA, 1999; Volume 81. [Google Scholar]
- Boston, R.S.; Viitanen, P.V.; Vierling, E. Molecular chaperones and protein folding in plants. In Post-Transcriptional Control of Gene Expression in Plants; Springer: Dordrecht, The Netherlands, 1996; pp. 191–222. [Google Scholar]
- Flaherty, K.M.; DeLuca-Flaherty, C.; McKay, D.B. Three-dimensional structure of the ATPase fragment of a 70 K heat-shock cognate protein. Nature 1990, 346, 623–628. [Google Scholar]
- Hartl, F.U.; Hayer-Hartl, M. Molecular chaperones in the cytosol: From nascent chain to folded protein. Science 2002, 295, 1852–1858. [Google Scholar]
- Zhu, X.; Zhao, X.; Burkholder, W.F.; Gragerov, A.; Ogata, C.M.; Gottesman, M.E.; Hendrickson, W.A. Structural analysis of substrate binding by the molecular chaperone DnaK. Science 1996, 272, 1606–1614. [Google Scholar]
- Lee, J.H.; Schöffl, F. An Hsp70 antisense gene affects the expression of HSP70/HSC70 the regulation of HSF and the acquisition of thermotolerance in transgenic Arabidopsis thaliana. Mol. Gen. Genet. 1996, 252, 11–19. [Google Scholar]
- Ono, K.; Hibino, T.; Kohinata, T.; Suzuki, S.; Tanaka, Y.; Nakamura, T.; Takabe, T. Overexpression of DnaK from a halotolerant cyanobacterium Aphanothece halophytica enhances the high-temperatue tolerance of tobacco during germination and early growth. Plant Sci. 2001, 160, 455–461. [Google Scholar]
- Sung, D.Y.; Guy, C.L. Physiological and molecular assessment of altered expression of Hsc70-1 in Arabidopsis Evidence for pleiotropic consequences. Plant Physiol. 2003, 132, 979–987. [Google Scholar]
- Alvim, F.C.; Carolino, S.M.; Cascardo, J.C.; Nunes, C.C.; Martinez, C.A.; Otoni, W.C.; Fontes, E.P. Enhanced accumulation of BiP in transgenic plants confers tolerance to water stress. Plant Physiol. 2001, 126, 1042–1054. [Google Scholar]
- Sugino, M.; Hibino, T.; Tanaka, Y.; Nii, N.; Takabe, T. Overexpression of DnaK from a halotolerant cyanobacterium Aphanothece halophytica acquires resistance to salt stress in transgenic tobacco plants. Plant Sci. 1999, 146, 81–88. [Google Scholar]
- Cho, E.K.; Choi, Y.J. A nuclear-localized HSP70 confers thermoprotective activity and drought-stress tolerance on plants. Biotechnol. Lett. 2009, 31, 597–606. [Google Scholar]
- Montero-Barrientos, M.; Hermosa, R.; Nicolás, C.; Cardoza, R.E.; Gutiérrez, S.; Monte, E. Overexpression of a Trichoderma HSP70 gene increases fungal resistance to heat and other abiotic stresses. Fungal Genet. Biol. 2008, 45, 1506–1513. [Google Scholar]
- Boorstein, W.R.; Ziegelhoffer, T.; Craig, E.A. Molecular evolution of the HSP70 multigene family. J. Mol. Evol. 1994, 38, 1–17. [Google Scholar]
- Tom, M.; Douek, J.; Yankelevich, I.; Bosch, T.C.; Rinkevich, B. Molecular characterization of the first heat shock protein 70 from a reef coral. Biochem. Biophy. Res. Commun. 1999, 262, 103–108. [Google Scholar]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar]
- Mayer, M.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar]
- Rensing, S.; Maier, U.-G. Phylogenetic analysis of the stress-70 protein family. J. Mol. Evol. 1994, 39, 80–86. [Google Scholar]
- Laszlo, A.; Davidson, T.; Hu, A.; Landry, J.; Bedford, J. Putative determinants of the cellular response to hyperthermia. Int. J. Radiat. Biol. 1993, 63, 569–581. [Google Scholar]
- Snoeckx, L.; Contard, F.; Samuel, J.; Marotte, F.; Rappaport, L. Expression and cellular distribution of heat-shock and nuclear oncogene proteins in rat hearts. Am. J. Physiol. Heart C 1991, 261, H1443–H1451. [Google Scholar]
- Wang, S.; Xie, W.; Rylander, M.N.; Tucker, P.W.; Aggarwal, S.; Diller, K.R. HSP70 kinetics study by continuous observation of HSP-GFP fusion protein expression on a perfusion heating stage. Biotechnol. Bioeng. 2008, 99, 146–154. [Google Scholar]
- Guinez, C.; Morelle, W.; Michalski, J.C.; Lefebvre, T. O-GlcNAc glycosylation: A signal for the nuclear transport of cytosolic proteins? Int. J. Biochem. Cell Biol. 2005, 37, 765–774. [Google Scholar]
- Qi, Y.; Wang, H.; Zou, Y.; Liu, C.; Liu, Y.; Wang, Y.; Zhang, W. Over-expression of mitochondrial heat shock protein 70 suppresses programmed cell death in rice. FEBS Lett. 2011, 585, 231–239. [Google Scholar]
- Park, H.-S.; Jeong, W.-J.; Kim, E.; Jung, Y.; Lim, J.M.; Hwang, M.S.; Park, E.-J.; Ha, D.-S.; Choi, D.-W. Heat shock protein gene family of the Porphyra seriata and enhancement of heat stress tolerance by PsHSP70 in Chlamydomonas. Mar. Biotechnol. 2012, 14, 332–342. [Google Scholar]
- Cho, E.K.; Hong, C.B. Over-expression of tobacco NtHSP70–1 contributes to drought-stress tolerance in plants. Plant Cell Rep. 2006, 25, 349–358. [Google Scholar]
- Chen, S.; Cui, X.; Chen, Y.; Gu, C.; Miao, H.; Gao, H.; Chen, F.; Liu, Z.; Guan, Z.; Fang, W. CgDREBa transgenic chrysanthemum confers drought and salinity tolerance. Environ. Exp. Bot. 2011, 74, 255–260. [Google Scholar]
- Hong, B.; Ma, C.; Yang, Y.; Wang, T.; Yamaguchi-Shinozaki, K.; Gao, J. Over-expression of AtDREB1A in chrysanthemum enhances tolerance to heat stress. Plant Mol. Biol. 2009, 70, 231–240. [Google Scholar]
- Chen, L.; Chen, Y.; Jiang, J.; Chen, S.; Chen, F.; Guan, Z.; Fang, W. The constitutive expression of Chrysanthemum dichrum ICE1 in Chrysanthemum grandiflorum improves the level of low temperature salinity and drought tolerance. Plant Cell Rep. 2012, 31, 1747–1758. [Google Scholar]
- Rivero, R.M.; Mestre, T.C.; Mittler, R.; Rubio, F.; Garcia-Sanchez, F.; Martinez, V. The combined effect of salinity and heat reveals a specific physiological biochemical and molecular response in tomato plants. Plant Cell Environ. 2013. [Google Scholar] [CrossRef]
- Sun, J.; Gu, J.; Zeng, J.; Han, S.; Song, A.; Chen, F.; Fang, W.; Jiang, J.; Chen, S. Changes in leaf morphology antioxidant activity and photosynthesis capacity in two different drought-tolerant cultivars of chrysanthemum during and after water stress. Sci. Hortic. 2013, 161, 249–258. [Google Scholar]
- Turóczy, Z.; Kis, P.; Török, K.; Cserháti, M.; Lendvai, A.; Dudits, D.; Horváth, G.V. Overproduction of a rice aldo–keto reductase increases oxidative and heat stress tolerance by malondialdehyde and methylglyoxal detoxification. Plant Mol. Biol. 2011, 75, 399–412. [Google Scholar]
- Farmer, E.E.; Mueller, M.J. ROS-mediated lipid peroxidation and RES-activated signaling. Annu. Rev. Plant Biol. 2013, 64, 429–450. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: metabolism oxidative stress and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar]
- Guo, S.; Wharton, W.; Moseley, P.; Shi, H. Heat shock protein 70 regulates cellular redox status by modulating glutathione-related enzyme activities. Cell Stress Chaperon. 2007, 12, 245. [Google Scholar]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar]
- Schroda, M.; Vallon, O.; Wollman, F.-A.; Beck, C.F. A chloroplast-targeted heat shock protein 70 (HSP70) contributes to the photoprotection and repair of photosystem II during and after photoinhibition. Plant Cell 1999, 11, 1165–1178. [Google Scholar]
- Chen, S.; Miao, H.; Chen, F.; Jiang, B.; Lu, J.; Fang, W. Analysis of expressed sequence tags (ESTs) collected from the inflorescence of chrysanthemum. Plant Mol. Biol. Rep. 2009, 27, 503–510. [Google Scholar]
- Earley, K.W.; Haag, J.R.; Pontes, O.; Opper, K.; Juehne, T.; Song, K.; Pikaard, C.S. Gateway-compatible vectors for plant functional genomics and proteomics. Plant J. 2006, 45, 616–629. [Google Scholar]
- Gao, H.; Song, A.; Zhu, X.; Chen, F.; Jiang, J.; Chen, Y.; Sun, Y.; Shan, H.; Gu, C.; Li, P. The heterologous expression in Arabidopsis of a Chrysanthemum Cys2/His2 zinc finger protein gene confers salinity and drought tolerance. Planta 2012, 235, 979–993. [Google Scholar]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method forAgrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar]
- Jung, C.; Seo, J.S.; Han, S.W.; Koo, Y.J.; Kim, C.H.; Song, S.I.; Nahm, B.H.; Choi, Y.D.; Cheong, J.-J. Overexpression of AtMYB44 enhances stomatal closure to confer abiotic stress tolerance in transgenic Arabidopsis. Plant Physiol. 2008, 146, 623–635. [Google Scholar]
- He, J.; Chen, F.; Chen, S.; Fang, W.; Miao, H.; Luo, H. Transformation of Lycoris longituba agglutinin gene to cut chrysanthemum and identification of aphid resistance in the transgenic plants. Acta Bot. Boreali-Occident. Sin. 2009, 29, 2318–2325. [Google Scholar]
- Gu, C.; Chen, S.; Liu, Z.; Shan, H.; Luo, H.; Guan, Z.; Chen, F. Reference gene selection for quantitative real-time PCR in chrysanthemum subjected to biotic and abiotic stress. Mol. Biotechnol. 2011, 49, 192–197. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−ΔΔC(T)) Method. Methods 2001, 25, 402–408. [Google Scholar]
- Gai, J. Statistics for Experiments; Agric Press: Beijing, China, 2000. [Google Scholar]
- Hong, B.; Tong, Z.; Ma, N.; Li, J.; Kasuga, M.; Yamaguchi-Shinozaki, K.; Gao, J. Heterologous expression of the AtDREB1A gene in chrysanthemum increases drought and salt stress tolerance. Sci. China C Life Sci. 2006, 49, 436–445. [Google Scholar]
- He, J.; Chen, F.; Chen, S.; Lv, G.; Deng, Y.; Fang, W.; Liu, Z.; Guan, Z.; He, C. Chrysanthemum leaf epidermal surface morphology and antioxidant and defense enzyme activity in response to aphid infestation. J. Plant Physiol. 2011, 168, 687–693. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]
- Yin, D.; Chen, S.; Chen, F.; Guan, Z.; Fang, W. Morphological and physiological responses of two chrysanthemum cultivars differing in their tolerance to waterlogging. Environ. Exp. Bot. 2009, 67, 87–93. [Google Scholar]
- Guha, A.; Sengupta, D.; Rasineni, G.K.; Reddy, A.R. An integrated diagnostic approach to understand drought tolerance in mulberry (Morus indica L). Flora 2010, 205, 144–151. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant lines | Semi-lethal temperature LT50 (°C) |
|---|---|
| WT | 47.7 ± 02 A |
| Vector | 47.9 ± 0.6 A |
| Th1 | 50.2 ± 0.2 B |
| Th4 | 50.3 ± 0.3 B |
| Primer | Sequences (5′-3′) | Annotation |
|---|---|---|
| HSP1 | TCAGTCCAAAGCGACATCAAA | For 3′-RACE |
| HSP2 | CAGCCTACTTCAACGACTCAC | For 3′-RACE |
| Adaptor-R | AGCAGTGGTATCAACGCAGAG | Universal primers for 3′-RACE |
| Q2-F | CACAAACCCTAATTCTTTTTCATACA | Full length cloning |
| Q2-R | AGTAGAACAGATAAATATCGACCACA | Full length cloning |
| HSP-ORF-F2 | CcccgggATGGCTGGTAAAGGGTGAA | Open reading frame (ORF) cloning |
| HSP-ORF-R2 | GCtctagaGTCGACCTCTTCGATCTTGG | Open reading frame (ORF) cloning |
| CgHSP-Dra-F | AGGCtttaaaATGGCTGGTAAAGGTGA | Intracellular localisation of CgHSP70 |
| CgHSP-Not-R | GTgcggccgcGAGTCGACCTCT | Intracellular localisation of CgHSP70 |
| 1301-F | TCCcccgggGTTATGACGCTGGGAATGTTTT | pCAMBIA1301 vector modified |
| 1301-R1 | GCtctagaAGATAATGCCACAGCACCTCTT | pCAMBIA1301 vector modified |
| 1301-R2 | cGAGCTCtctagaAGATAATGCCACAGCACCTCTT | pCAMBIA1301 vector modified |
| GAPDH-F | GCTGTATCCCCATTCGTT | qRT-PCR |
| GAPDH-R | AGAAGGCAAGCTCAAGGG | qRT-PCR |
| HSP-F | GCTTGCTGAGGCTGATGAGT | qRT-PCR |
| HSP-R | ACCTGATGGTGCGGGTTCCTC | qRT-PCR |
| HptII-F | CTCGATGAG CTGATGCTTTGGG | verifying positive transformants |
| HptII-R | GCTTCTGCGGGCGATTTGTGTA | Verifying positive transformants |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Song, A.; Zhu, X.; Chen, F.; Gao, H.; Jiang, J.; Chen, S. A Chrysanthemum Heat Shock Protein Confers Tolerance to Abiotic Stress. Int. J. Mol. Sci. 2014, 15, 5063-5078. https://doi.org/10.3390/ijms15035063
Song A, Zhu X, Chen F, Gao H, Jiang J, Chen S. A Chrysanthemum Heat Shock Protein Confers Tolerance to Abiotic Stress. International Journal of Molecular Sciences. 2014; 15(3):5063-5078. https://doi.org/10.3390/ijms15035063
Chicago/Turabian StyleSong, Aiping, Xirong Zhu, Fadi Chen, Haishun Gao, Jiafu Jiang, and Sumei Chen. 2014. "A Chrysanthemum Heat Shock Protein Confers Tolerance to Abiotic Stress" International Journal of Molecular Sciences 15, no. 3: 5063-5078. https://doi.org/10.3390/ijms15035063