Genome-Wide Identification, Evolution and Expression Analysis of the Grape (Vitis vinifera L.) Zinc Finger-Homeodomain Gene Family

Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification of V. vinifera ZHD (VvZHD) Genes
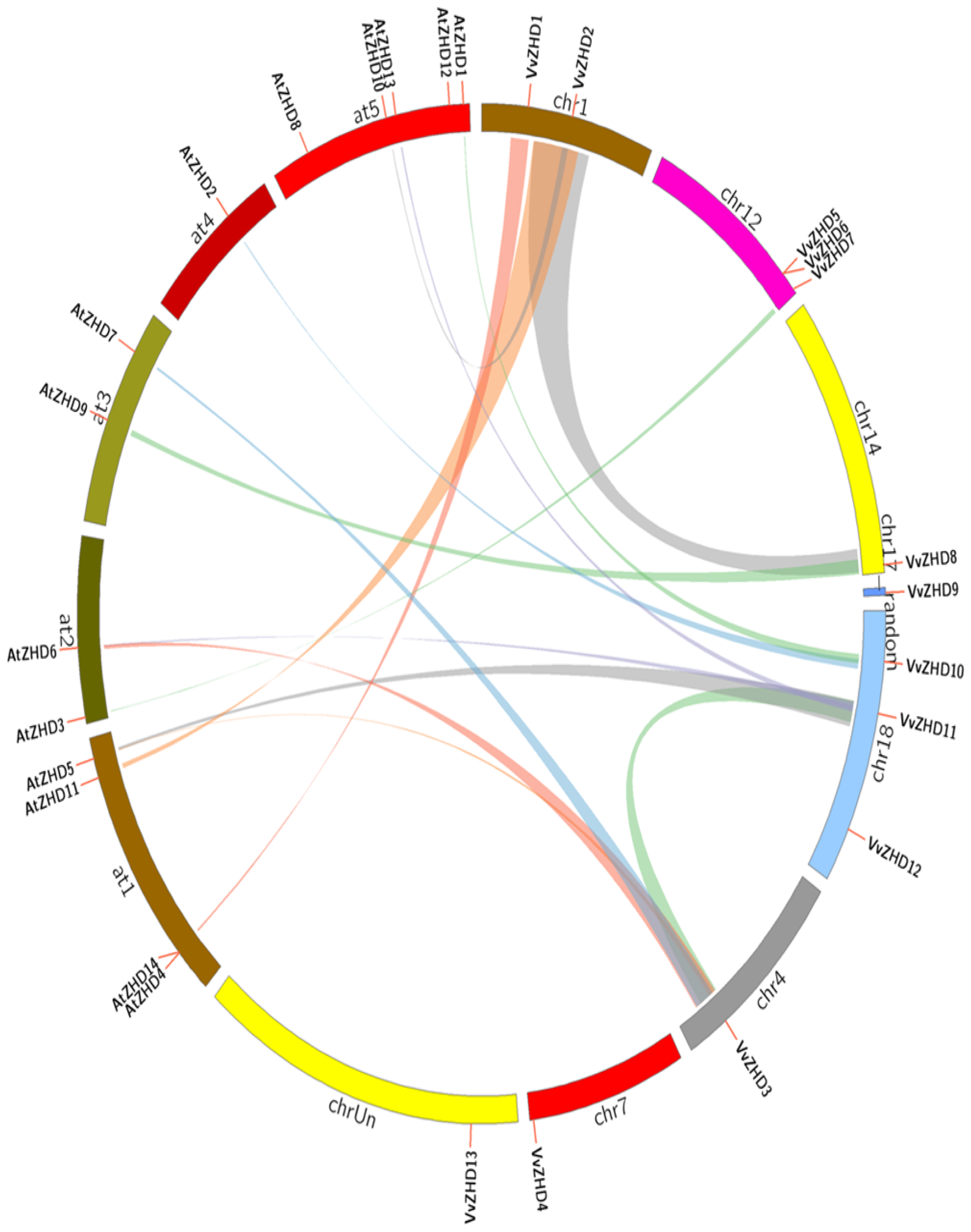
2.2. Expansion Patterns of the Grape ZHD Gene Family
2.3. Evolutionary Relationship of Grape and Arabidopsis ZHD Genes
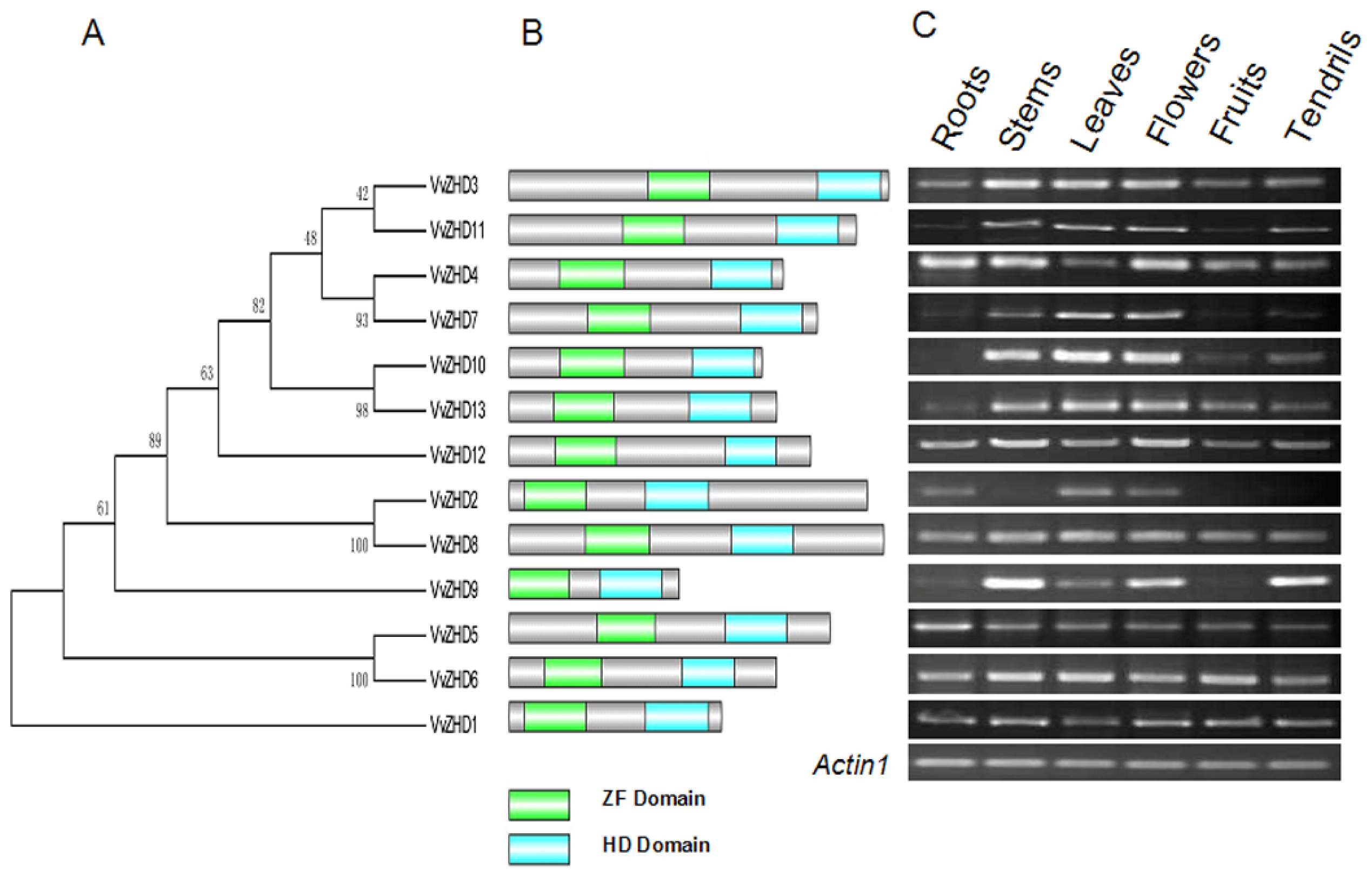
2.4. Phylogenetic Analysis of Grape ZHD Genes
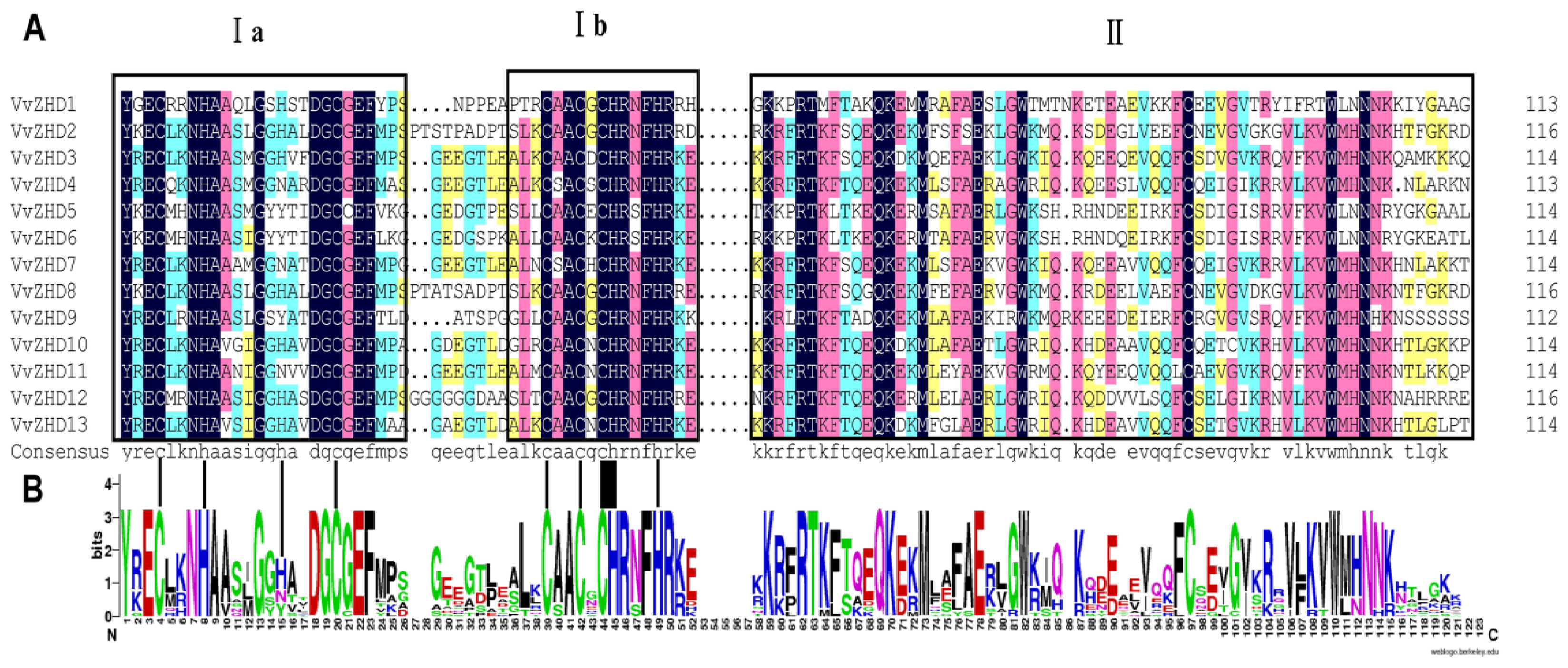
2.5. Sequence Analysis of Grape ZHD Genes
2.6. Expression Profiles of Grape ZHD Genes in Different Organs
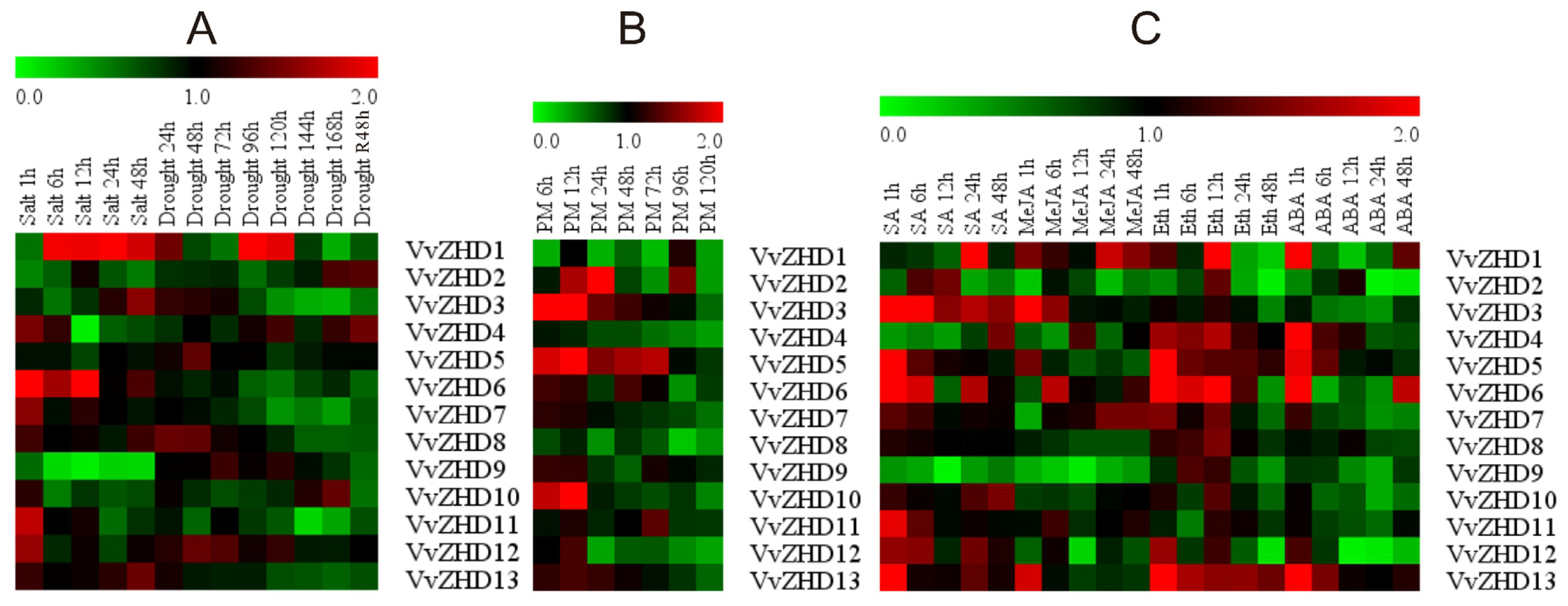
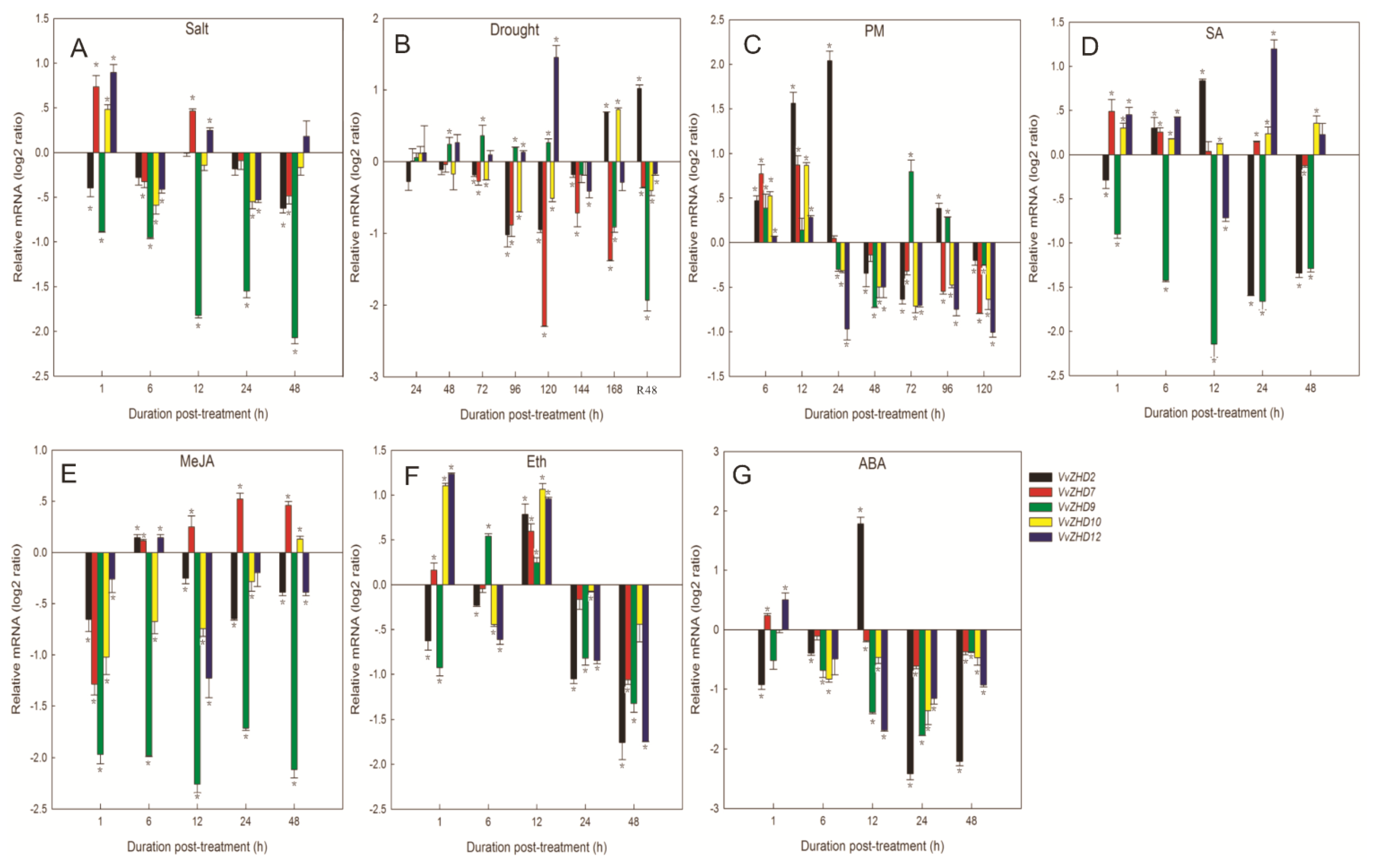
2.7. Expression Patterns of Grape ZHD Genes under Different Stress Conditions and Following Exogenous Hormone Treatments
2.7.1. Abiotic Stresses
2.7.2. Biotic Stress
2.7.3. Hormone Treatments
3. Discussion
3.1. Evolution of Grape VvZHD Genes
3.2. Expansion and Synteny of the VvZHD Gene Family
3.3. Spatial Expression Patterns of VvZHD Genes in Various Grape Tissues
3.4. VvZHD Proteins Play Important Roles in a Range of Biological Processes
4. Experimental Section
4.1. Identification and Annotation of Grape ZHD (VvZHD) Genes
4.2. Determination of Chromosomal Location and Synteny Analysis
4.3. Sequence Alignments, Phylogenetic Analysis and Domain Location of VvZHD Genes
4.4. Plant Material
4.5. Abiotic, Hormone and Biotic Stress Treatment
4.6. Semi-Quantitative RT-PCR and Real-Time PCR Analysis
5. Conclusions
Supplementary Information
ijms-15-05730-s001.pdfAcknowledgments
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| SA | salicylic acid |
| Eth | ethylene |
| MeJA | methyl jasmonate |
| ORF | open reading frame |
| qRT-PCR | quantitative reverse transcription PCR |
| ZHD | zinc finger-homeodomain |
References
- Espley, R.V.; Hellens, R.P.; Putterill, J.; Stevenson, D.E.; Kutty-Amma, S.; Allan, A.C. Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J 2007, 49, 414–427. [Google Scholar]
- Czemmel, S.; Heppel, S.C.; Bogs, J. R2R3 MYB transcription factors: Key regulators of the flavonoid biosynthetic pathway in grapevine. Protoplasma 2012, 249, 109–118. [Google Scholar]
- Bürglin, T.R.A. Comprehensive classification of homeobox genes. In Guidebook to the Homeobox Genes; Oxford University Press: Oxford, UK, 1994; pp. 25–71. [Google Scholar]
- Williams, R.W. Plant homeobox genes: many functions stem from a common motif. Bioessays 1998, 20, 280–282. [Google Scholar]
- Ito, M.; Sato, Y.; Matsuoka, M. Involvement of homeobox genes in early body plan of monocot. Int. Rev. Cytol 2002, 218, 1–35. [Google Scholar]
- Akin, Z.N.; Nazarali, A.J. Hox genes and their candidate downstream targets in the developing central nervous system. Cell. Mol. Neurobiol 2005, 25, 697–741. [Google Scholar]
- Hunter, C.S.; Rhodes, S.J. LIM-homeodomain genes in mammalian development and human disease. Mol. Biol. Rep 2005, 32, 67–77. [Google Scholar]
- Ariel, F.D.; Manavella, P.A.; Dezar, C.A.; Chan, R.L. The true story of the HD-Zip family. Trends Plant Sci 2007, 12, 419–426. [Google Scholar]
- Takatsuji, H. Zinc-finger proteins: The classical zinc finger emerges in contemporary plant science. Plant Mol. Biol 1999, 39, 1073–1078. [Google Scholar]
- Krishna, S.S.; Majumdar, I.; Grishin, N.V. Structural classification of zinc fingers: Survey and summary. Nucleic Acids Res 2003, 31, 532–550. [Google Scholar]
- Klug, A.; Schwabe, J.W. Protein motifs 5. Zinc fingers. FASEB J.: Off. Public. Feder. Am. Soc. Exp. Biol 1995, 9, 597–604. [Google Scholar]
- Halbach, T.; Scheer, N.; Werr, W. Transcriptional activation by the PHD finger is inhibited through an adjacent leucine zipper that binds 14-3-3 proteins. Nucleic Acids Res 2000, 28, 3542–3550. [Google Scholar]
- Li, J.; Jia, D.; Chen, X. HUA1, a regulator of stamen and carpel identities in Arabidopsis, codes for a nuclear RNA binding protein. Plant Cell 2001, 13, 2269–2281. [Google Scholar]
- Kosarev, P.; Mayer, K.F.; Hardtke, C.S. Evaluation and classification of RING-finger domains encoded by the Arabidopsis genome. Genome Biol 2002, 3, RESEARCH0016. [Google Scholar]
- Englbrecht, C.C.; Schoof, H.; Bohm, S. Conservation, diversification and expansion of C2H2 zinc finger proteins in the Arabidopsis thaliana genome. BMC Genomics 2004, 5, 39. [Google Scholar]
- Yanagisawa, S. Dof domain proteins: Plant-specific transcription factors associated with diverse phenomena unique to plants. Plant Cell Physiol 2004, 45, 386–391. [Google Scholar]
- Sakamoto, H.; Maruyama, K.; Sakuma, Y.; Meshi, T.; Iwabuchi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis Cys2/His2-type zinc-finger proteins function as transcription repressors under drought, cold, and high-salinity stress conditions. Plant Physiol 2004, 136, 2734–2746. [Google Scholar]
- Mittler, R.; Kim, Y.; Song, L.; Coutu, J.; Coutu, A.; Ciftci-Yilmaz, S.; Lee, H.; Stevenson, B.; Zhu, J.K. Gain- and loss-of-function mutations in Zat10 enhance the tolerance of plants to abiotic stress. FEBS Lett 2006, 580, 6537–6542. [Google Scholar]
- Huang, J.; Yang, X.; Wang, M.M.; Tang, H.J.; Ding, L.Y.; Shen, Y.; Zhang, H.S. A novel rice C2H2-type zinc finger protein lacking DLN-box/EAR-motif plays a role in salt tolerance. Biochim. Biophys. Acta 2007, 1769, 220–227. [Google Scholar]
- Xu, D.Q.; Huang, J.; Guo, S.Q.; Yang, X.; Bao, Y.M.; Tang, H.J.; Zhang, H.S. Overexpression of a TFIIIA-type zinc finger protein gene ZFP252 enhances drought and salt tolerance in rice (Oryza sativa L.). FEBS Lett 2008, 582, 1037–1043. [Google Scholar]
- Windhovel, A.; Hein, I.; Dabrowa, R.; Stockhaus, J. Characterization of a novel class of plant homeodomain proteins that bind to the C4 phosphoenolpyruvate carboxylase gene of Flaveria trinervia. Plant Mol. Biol 2001, 45, 201–214. [Google Scholar]
- Hong, S.-Y.; Kim, O.-K.; Kim, S.-G.; Yang, M.-S.; Park, C.-M. Nuclear import and DNA binding of the ZHD5 transcription factor is modulated by a competitive peptide inhibitor in Arabidopsis. J. Biol. Chem 2011, 286, 1659–1668. [Google Scholar]
- Figueiredo, D.D.; Barros, P.M.; Cordeiro, A.M.; Serra, T.S.; Lourenço, T.; Chander, S.; Oliveira, M.M.; Saibo, N.J. Seven zinc-finger transcription factors are novel regulators of the stress responsive gene OsDREB1B. J. Exp. Bot 2012, 63, 3643–3656. [Google Scholar]
- Park, H.C.; Kim, M.L.; Lee, S.M.; Bahk, J.D.; Yun, D.J.; Lim, C.O.; Hong, J.C.; Lee, S.Y.; Cho, M.J.; Chung, W.S. Pathogen-induced binding of the soybean zinc finger homeodomain proteins GmZF-HD1 and GmZF-HD2 to two repeats of ATTA homeodomain binding site in the calmodulin isoform 4 (GmCaM4) promoter. Nucleic Acids Res 2007, 35, 3612–3623. [Google Scholar]
- Abu-Romman, S. Molecular cloning and expression analysis of zinc finger-homeodomain transcription factor TaZFHD1 in wheat. S. Afr. J. Bot 2014, 91, 32–36. [Google Scholar]
- Tan, Q.K.-G.; Irish, V.F. The Arabidopsis zinc finger-homeodomain genes encode proteins with unique biochemical properties that are coordinately expressed during floral development. Plant Physiol 2006, 140, 1095–1108. [Google Scholar]
- Tran, L.S.P.; Nakashima, K.; Sakuma, Y.; Osakabe, Y.; Qin, F.; Simpson, S.D.; Maruyama, K.; Fujita, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Co-expression of the stress-inducible zinc finger homeodomain ZFHD1 and NAC transcription factors enhances expression of the ERD1 gene in Arabidopsis. Plant J 2007, 49, 46–63. [Google Scholar]
- Hu, W.; dePamphilis, C.W.; Ma, H. Phylogenetic analysis of the plant-specific zinc finger-homeobox and mini zinc finger gene families. J. Integr. Plant Biol 2008, 50, 1031–1045. [Google Scholar]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, U463–U465. [Google Scholar]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Yong, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families inArabidopsis thaliana. BMC Plant Biol 2004, 4, 10. [Google Scholar]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol 2006, 9, 436–442. [Google Scholar]
- Huang, D.Q.; Wu, W.R.; Abrams, S.R.; Cutler, A.J. The relationship of drought-related gene expression in Arabidopsis thaliana to hormonal and environmental factors. J. Exp. Bot 2008, 59, 2991–3007. [Google Scholar]
- Finkelstein, R.R.; Gampala, S.S.L.; Rock, C.D. Abscisic acid signaling in seeds and seedlings. Plant Cell 2002, 14, S15–S45. [Google Scholar]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exon-intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar]
- Maere, S.; de Bodt, S.; Raes, J.; Casneuf, T.; van Montagu, M.; Kuiper, M.; van de Peer, Y. Modeling gene and genome duplications in eukaryotes. Proc. Natl. Acad. Sci. USA 2005, 102, 5454–5459. [Google Scholar]
- Blanc, G.; Hokamp, K.; Wolfe, K.H. A recent polyploidy superimposed on older large-scale duplications in the Arabidopsis genome. Genome Res 2003, 13, 137–144. [Google Scholar]
- Velasco, R.; Zharkikh, A.; Troggio, M.; Cartwright, D.A.; Cestaro, A.; Pruss, D.; Pindo, M.; FitzGerald, L.M.; Vezzulli, S.; Reid, J.; et al. A high quality draft consensus sequence of the genome of a heterozygous grapevine variety. PLoS One 2007, 2, e1326. [Google Scholar]
- Lyons, E.; Pedersen, B.; Kane, J.; Alam, M.; Ming, R.; Tang, H.; Wang, X.; Bowers, J.; Paterson, A.; Lisch, D.; et al. Finding and comparing syntenic regions among Arabidopsis and the outgroups papaya, poplar, and grape: CoGe with rosids. Plant Physiol 2008, 148, 1772–1781. [Google Scholar]
- Zhang, Y.C.; Gao, M.; Singer, S.D.; Fei, Z.J.; Wang, H.; Wang, X.P. Genome-wide identification and analysis of the TIFY gene family in grape. PLoS One 2012, 7, e44465. [Google Scholar]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol 2005, 43, 205–227. [Google Scholar]
- Mauch-Mani, B.; Mauch, F. The role of abscisic acid in plant-pathogen interactions. Curr. Opin. Plant Biol 2005, 8, 409–414. [Google Scholar]
- Davies, W.J.; Jones, H.G. Abscisic Acid: Physiology and Biochemistry; BIOS Scientific Publishers: Oxford, UK, 1991. [Google Scholar]
- Huang, W.Z.; Ma, X.R.; Wang, Q.L.; Gao, Y.F.; Xue, Y.; Niu, X.L.; Yu, G.R.; Liu, Y.S. Significant improvement of stress tolerance in tobacco plants by overexpressing a stress-responsive aldehyde dehydrogenase gene from maize (Zea mays). Plant Mol. Biol 2008, 68, 451–463. [Google Scholar]
- Kotchoni, S.O.; Kuhns, C.; Ditzer, A.; Kirch, H.H.; Bartels, D. Over-expression of different aldehyde dehydrogenase genes in Arabidopsis thaliana confers tolerance to abiotic stress and protects plants against lipid peroxidation and oxidative stress. Plant Cell Environ 2006, 29, 1033–1048. [Google Scholar]
- Rodrigues, S.M.; Andrade, M.O.; Gomes, A.P.S.; DaMatta, F.M.; Baracat-Pereira, M.C.; Fontes, E.P.B. Arabidopsis and tobacco plants ectopically expressing the soybean antiquitin-like ALDH7 gene display enhanced tolerance to drought, salinity, and oxidative stress. J. Exp. Bot 2006, 57, 1909–1918. [Google Scholar]
- Wellcome Trust Sanger Institute. Available online: http://www.sanger.ac.uk (accessed on 8 August 2013).
- HMMER. Available online: http://hmmer.janelia.org/search/hmmsearch (accessed on 8 September 2013).
- Genome Database for Rosaceae. Available online: http://www.rosaceae.org/projects/grape_genome (accessed on 8 September 2013).
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.Z.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar]
- Zhang, Y.C.; Mao, L.Y.; Wang, H.; Brocker, C.; Yin, X.J.; Vasiliou, V.; Fei, Z.J.; Wang, X.P. Genome-wide identification and analysis of grape aldehyde dehydrogenase (ALDH) gene superfamily. PLoS One 2012, 7, e32153. [Google Scholar]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res 2012, 40, e49. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 1997, 25, 4876–4882. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol 2011, 28, 2731–2739. [Google Scholar]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res 2012, 40, D302–D305. [Google Scholar]
- SMART. Available online: http://smart.embl-heidelberg.de/smart/set_mode.cgi?NORMAL=1 (accessed on 18 August 2013).
- Ren, J.; Wen, L.P.; Gao, X.J.; Jin, C.J.; Xue, Y.; Yao, X.B. DOG 1.0: Illustrator of protein domain structures. Cell Res 2009, 19, 271–273. [Google Scholar]
- DOG 1.0—Protein Domain Structure Visualization. Available online: http://dog.biocuckoo.org/ (accessed on 18 August 2013).
- Boneh, U.; Biton, I.; Zheng, C.L.; Schwartz, A.; Ben-Ari, G. Characterization of potential ABA receptors in Vitis vinifera. Plant Cell Rep 2012, 31, 311–321. [Google Scholar]
- Upreti, K.K.; Murti, G.S.R. Response of grape rootstocks to salinity: Changes in root growth, polyamines and abscisic acid. Biol. Plant 2010, 54, 730–734. [Google Scholar]
- Cramer, G.R.; Ergul, A.; Grimplet, J.; Tillett, R.L.; Tattersall, E.A.R.; Bohlman, M.C.; Vincent, D.; Sonderegger, J.; Evans, J.; Osborne, C.; et al. Water and salinity stress in grapevines: Early and late changes in transcript and metabolite profiles. Funct. Integr. Genomics 2007, 7, 111–134. [Google Scholar]
- Yang, Y.Z.; He, M.Y.; Zhu, Z.G.; Li, S.X.; Xu, Y.; Zhang, C.H.; Singer, S.D.; Wang, Y.J. Identification of the dehydrin gene family from grapevine species and analysis of their responsiveness to various forms of abiotic and biotic stress. BMC Plant Biol 2012, 12, 140. [Google Scholar]
- Li, H.E.; Xu, Y.; Xiao, Y.; Zhu, Z.G.; Xie, X.Q.; Zhao, H.Q.; Wang, Y.J. Expression and functional analysis of two genes encoding transcription factors, VpWRKY1 and VpWRKY2, isolated from Chinese wild Vitis pseudoreticulata. Planta 2010, 232, 1325–1337. [Google Scholar]
- Wang, L.J.; Li, S.H. Thermotolerance and related antioxidant enzyme activities induced by heat acclimation and salicylic acid in grape (Vitis vinifera L.) leaves. Plant Growth Regul 2006, 48, 137–144. [Google Scholar]
- Xiao, H.G.; Nassuth, A. Stress- and development-induced expression of spliced and unspliced transcripts from two highly similar dehydrin 1 genes in V-riparia and V-vinifera. Plant Cell Rep 2006, 25, 968–977. [Google Scholar]
- Wang, Y.; Liu, Y.; He, P.; Chen, J.; Lamikanra, O.; Lu, J. Evaluation of foliar resistance to uncinula necator in chinese wild vitis species. Vitis 1995, 34, 159–164. [Google Scholar]
- Zhang, J.J.; Wang, Y.J.; Wang, X.P.; Yang, K.Q.; Yang, J.X. An improved method for rapidly extracting total RNA from Vitis. Fruit Sci 2003, 53, 771–787. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene name | Gene locus | Accession number | Chromosome location | Strand | CDS (bp) | Protein (aa) |
|---|---|---|---|---|---|---|
| VvZHD1 | - | XM_002281662.1 | chr1:6167005...6167589 | − | 585 | 194 |
| VvZHD2 | GSVIVT01019981001 | XM_002273766.1 | chr1:12042374...12043354 | − | 981 | 326 |
| VvZHD3 | GSVIVT01018947001 | XM_002266541.2 | chr4:18452642...18453679 | + | 1038 | 345 |
| VvZHD4 | GSVIVT01000250001 | XM_002267711.1 | chr7:20539010...20539762 | − | 753 | 250 |
| VvZHD5 | GSVIVT01023289001 | XM_002264255.1 | chr12:20065353...20066231 | + | 879 | 292 |
| VvZHD6 | - | XM_002264214.1 | chr12:20068964...20069695 | − | 732 | 243 |
| VvZHD7 | GSVIVT01023137001 | XM_003633364.1 | chr12:22175301...22176146 | − | 846 | 281 |
| VvZHD8 | GSVIVT01011413001 | XM_002283497.2 | chr14:29460743...29461768 | + | 1026 | 341 |
| VvZHD9 | GSVIVT01000386001 | XM_002263430.1 | chr17:random 525867...526334 | + | 468 | 155 |
| VvZHD10 | GSVIVT01009128001 | XM_002285673.1 | chr18:5343830...5344522 | + | 693 | 230 |
| VvZHD11 | GSVIVT01009721001 | XM_002281335.2 | chr18:10783147...10784097 | + | 951 | 316 |
| VvZHD12 | GSVIVT01012772001 | XM_002276544.1 | chr18:23508908...23509735 | − | 828 | 275 |
| VvZHD13 | GSVIVT01003614001 | XM_003635009.1 | chrUn:11277853...11280416 | − | 735 | 244 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, H.; Yin, X.; Li, X.; Wang, L.; Zheng, Y.; Xu, X.; Zhang, Y.; Wang, X. Genome-Wide Identification, Evolution and Expression Analysis of the Grape (Vitis vinifera L.) Zinc Finger-Homeodomain Gene Family. Int. J. Mol. Sci. 2014, 15, 5730-5748. https://doi.org/10.3390/ijms15045730
Wang H, Yin X, Li X, Wang L, Zheng Y, Xu X, Zhang Y, Wang X. Genome-Wide Identification, Evolution and Expression Analysis of the Grape (Vitis vinifera L.) Zinc Finger-Homeodomain Gene Family. International Journal of Molecular Sciences. 2014; 15(4):5730-5748. https://doi.org/10.3390/ijms15045730
Chicago/Turabian StyleWang, Hao, Xiangjing Yin, Xiaoqin Li, Li Wang, Yi Zheng, Xiaozhao Xu, Yucheng Zhang, and Xiping Wang. 2014. "Genome-Wide Identification, Evolution and Expression Analysis of the Grape (Vitis vinifera L.) Zinc Finger-Homeodomain Gene Family" International Journal of Molecular Sciences 15, no. 4: 5730-5748. https://doi.org/10.3390/ijms15045730