A20 Overexpression Inhibits Lipopolysaccharide-Induced NF-κB Activation, TRAF6 and CD40 Expression in Rat Peritoneal Mesothelial Cells

Abstract
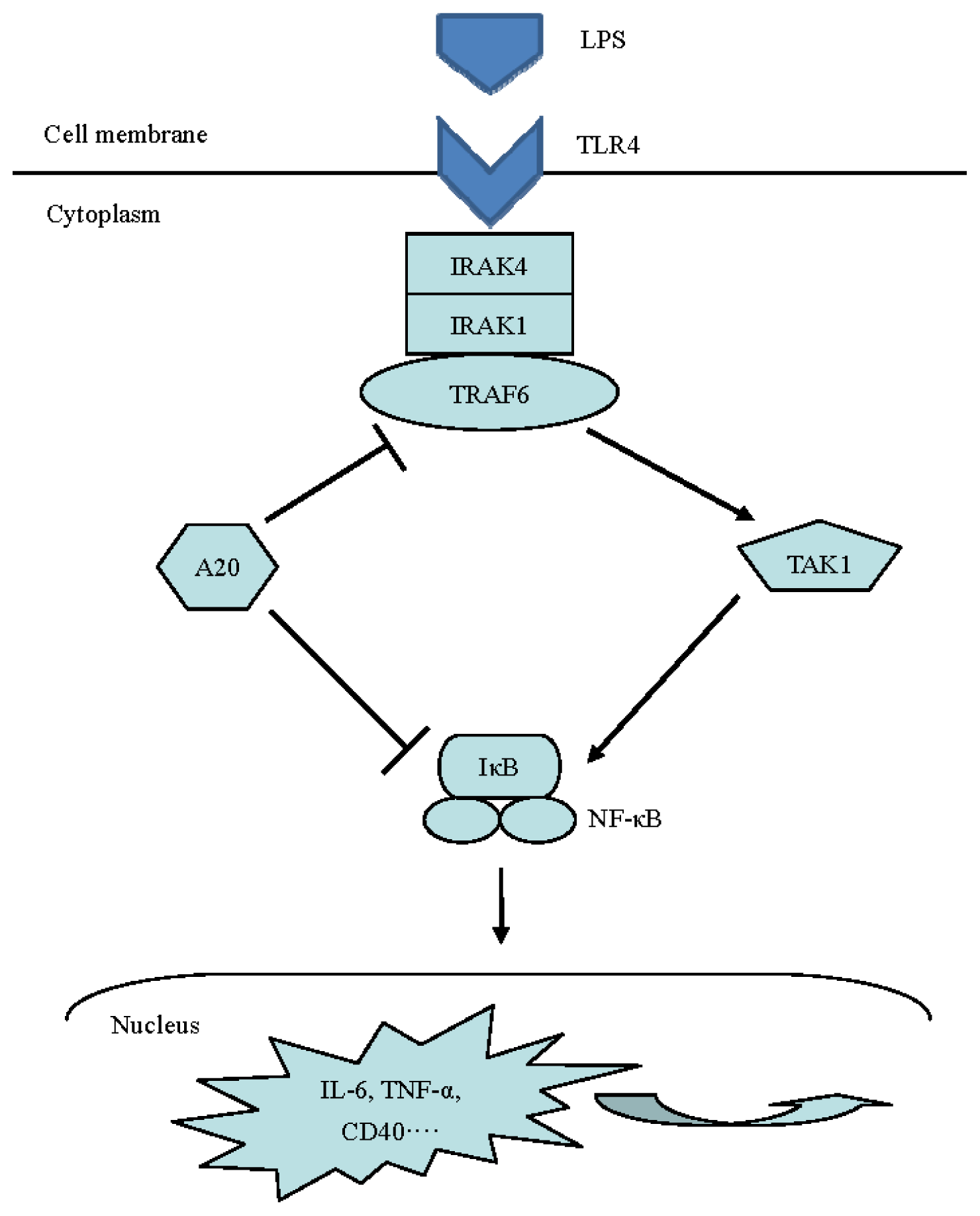
:1. Introduction
2. Results
2.1. Culture of RPMCs
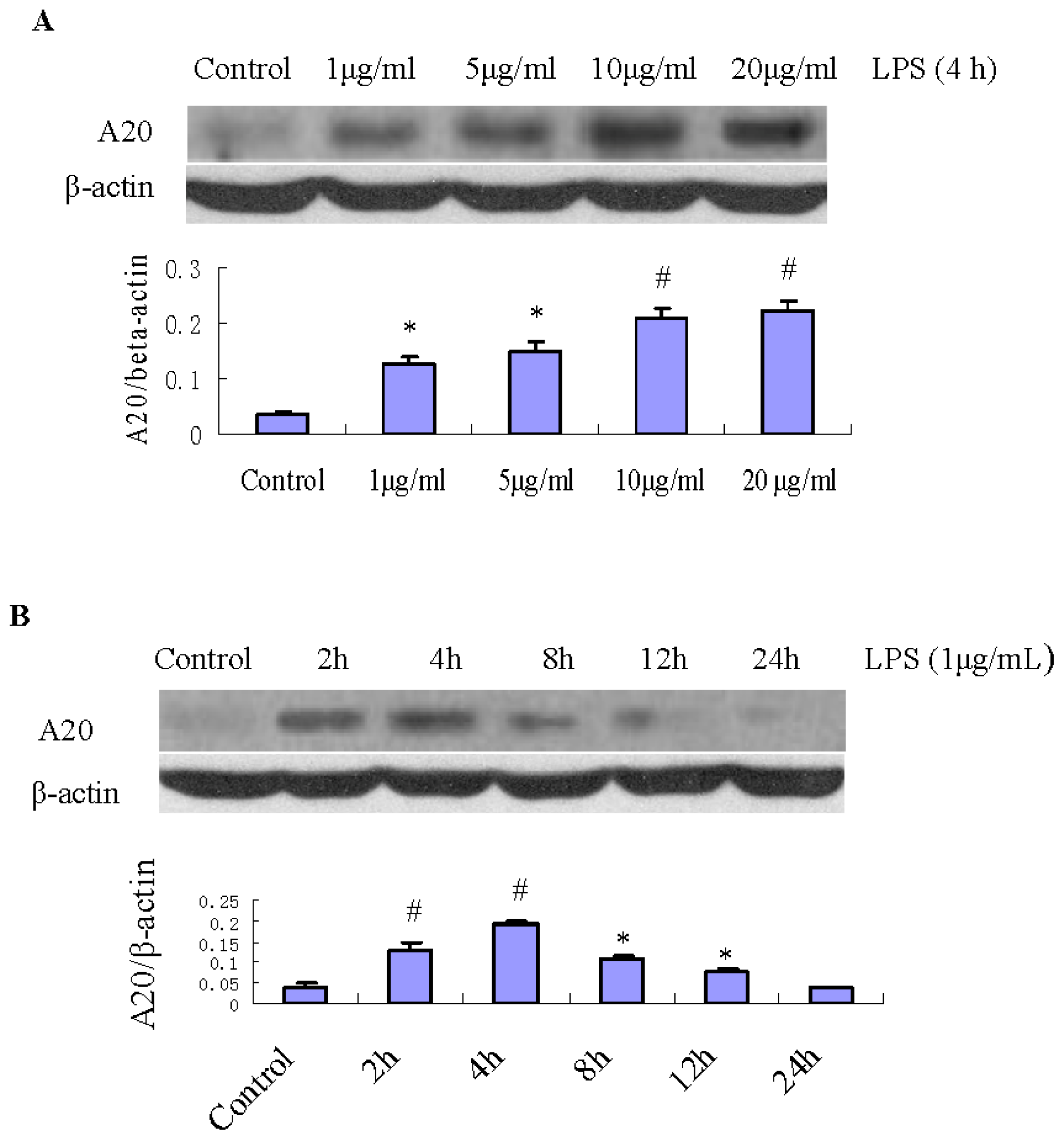
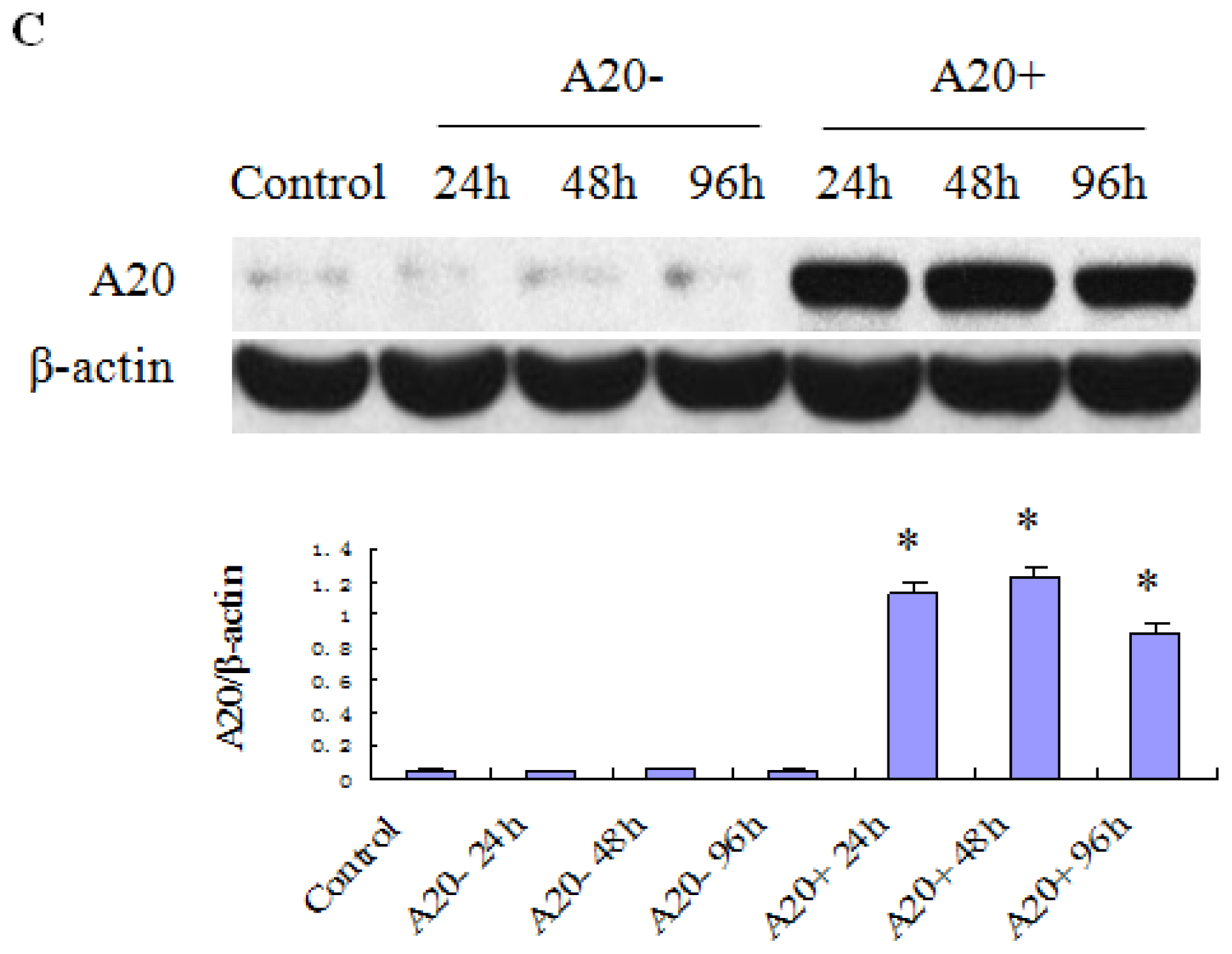
2.2. Expression of A20 Protein in RPMCs
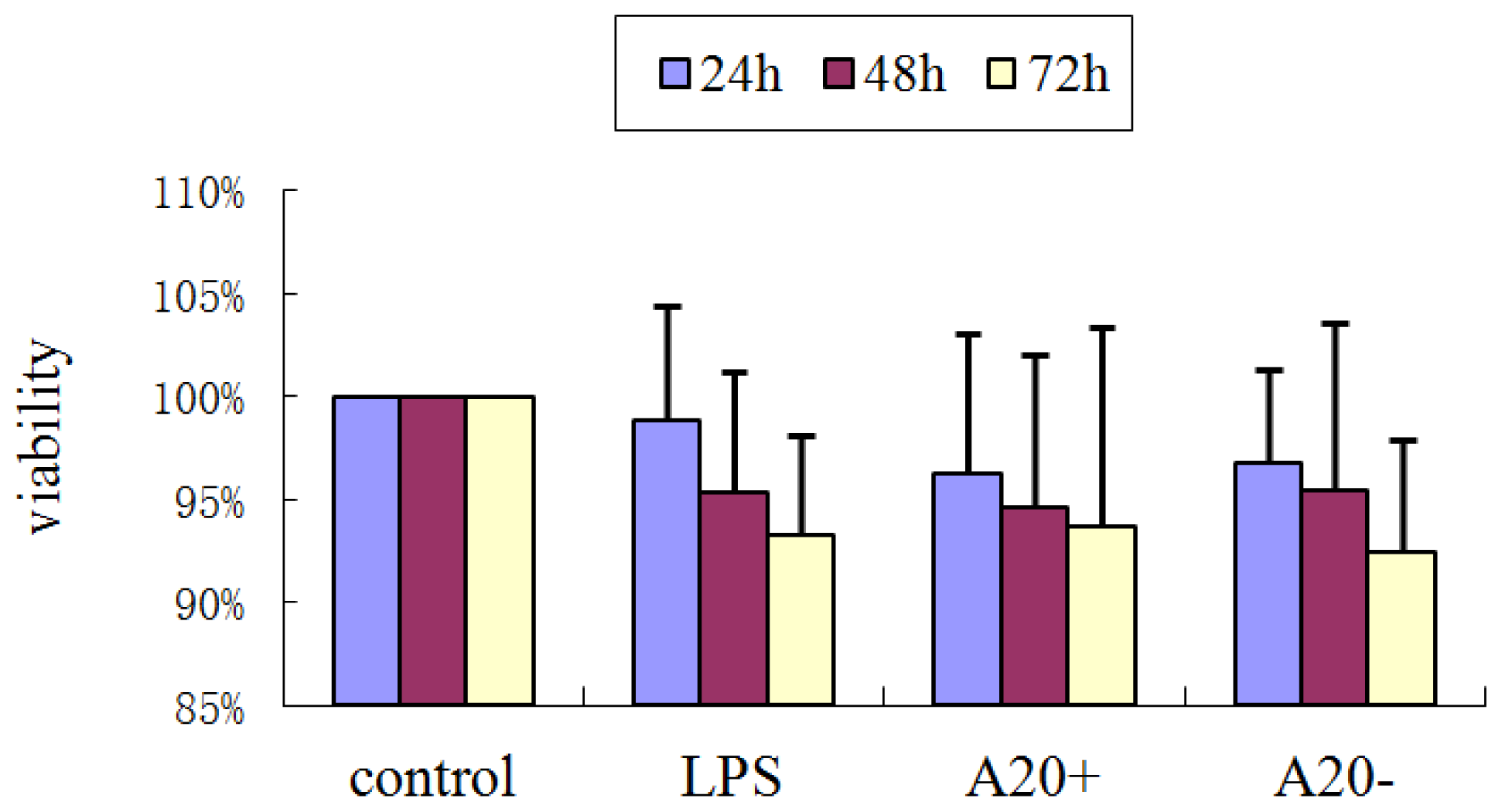
2.3. Effects of Various Treatments on RPMC Viability
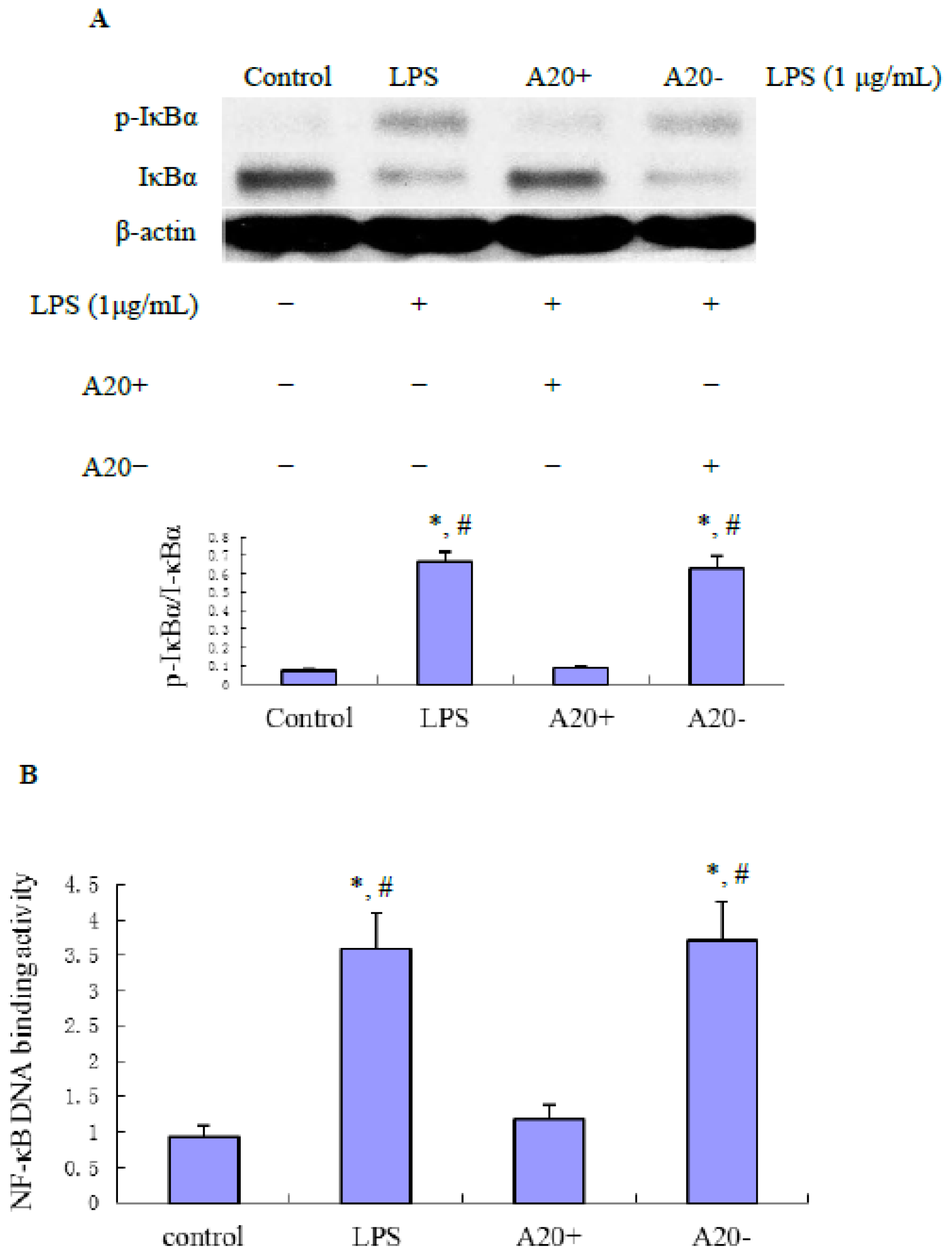
2.4. A20 Overexpression Effectively Suppresses LPS-Induced NF-κB Activation in RPMCs
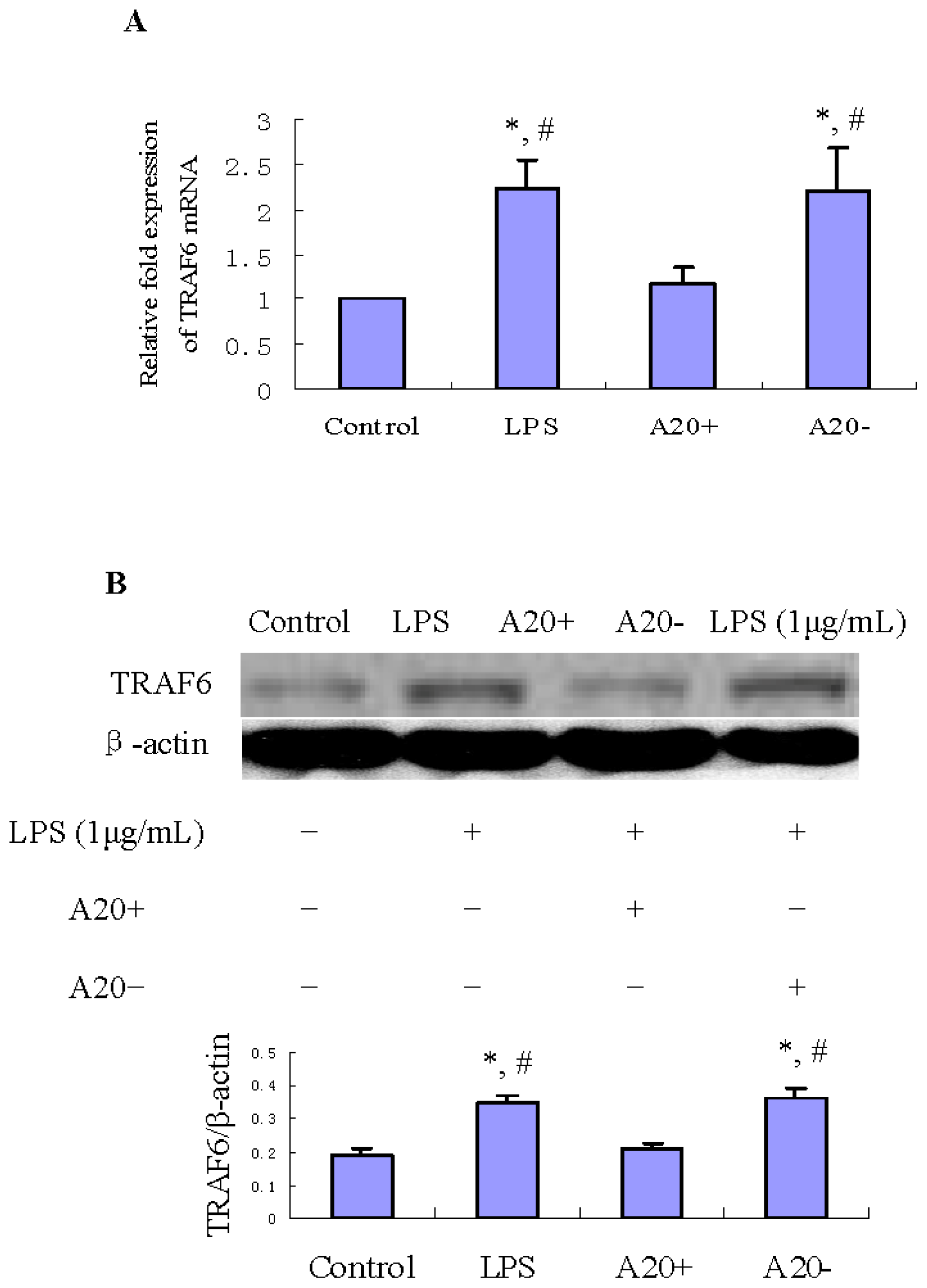
2.5. A20 Overexpression Suppresses LPS-Induced TRAF6 Expression in RPMCs
2.6. A20 Overexpression Suppresses LPS-Induced CD40 Expression in RPMCs
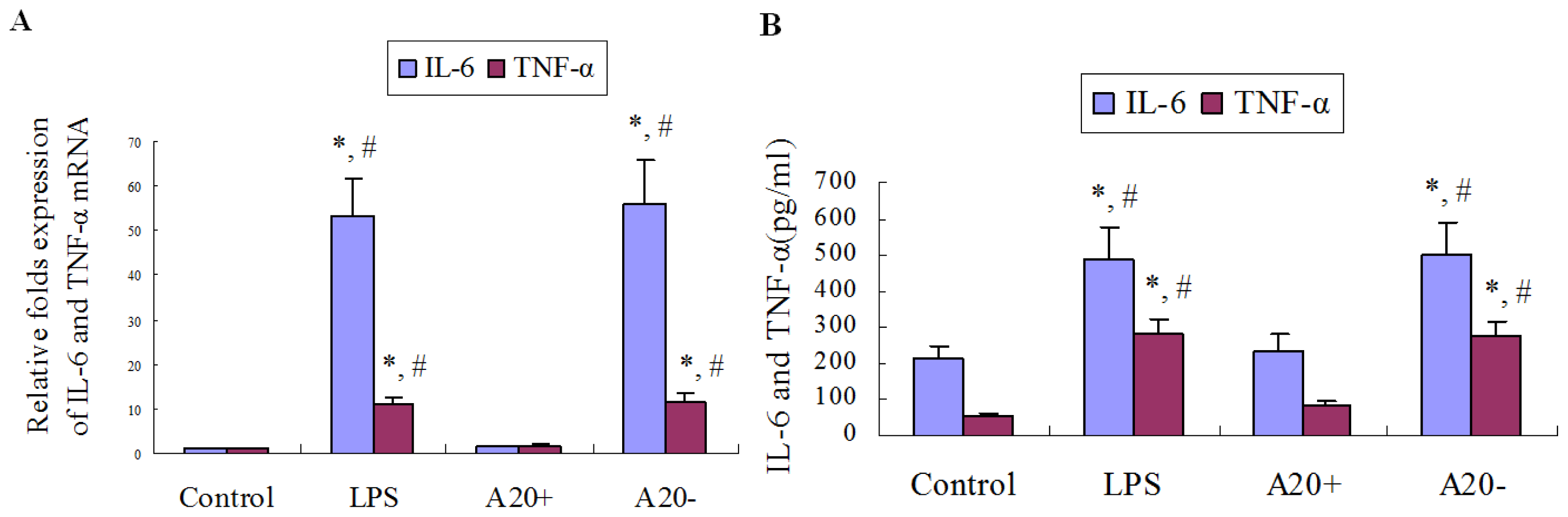
2.7. A20 Overexpression Suppresses LPS-Induced IL-6 and TNF-α Expression in RPMCs
3. Discussion
4. Materials and Methods
4.1. Antibodies and Reagents
4.2. RPMCs Isolation and Culture
4.3. Effect of LPS on Expression of A20 in RPMCs
4.4. Transfection Experiments
4.5. Assessment of Cell Viability
4.6. Effect of A20 on LPS-Induced p-IκBα, IκBα, TRAF6, CD40, IL-6 and TNF-α Expression
4.7. Western Blot Assay
4.8. NF-κB DNA Binding Activity
4.9. Real-Time PCR
4.10. Cytokine Assays
4.11. Statistical Analysis
5. Conclusions
Supplementary Information
ijms-15-06592-s001.pdfAcknowledgments
Conflicts of Interest
- Author ContributionsX.L.Z. and D.A.P. are responsible for conception and design of the research; J.Z.Y. and P.W. performed the experiments; X.L.Z. and D.A.P. analyzed the data, interpreted the results of the experiments and drafted the manuscript; X.L.Z. and J.Z.Y. prepared the figures; X.L.Z. and G.X. edited and revised the manuscript; X.L.Z. approved the final version of the manuscript.
References
- Krediet, R.T. 30 years of peritoneal dialysis development: The past and the future. Perit. Dial. Int 2007, 27, S35–S41. [Google Scholar]
- Gokal, R. Peritoneal dialysis in the 21st century: An analysis of current problems and future developments. J. Am. Soc. Nephrol 2002, 13, S104–S116. [Google Scholar]
- Szeto, C.C.; Chow, K.M. Gram-negative peritonitis—The Achilles heel of peritoneal dialysis? Perit. Dial. Int 2007, 27, S267–S271. [Google Scholar]
- Devuyst, O.; Margetts, P.J.; Topley, N. The pathophysiology of the peritoneal membrane. J. Am. Soc. Nephrol 2010, 21, 1077–1085. [Google Scholar]
- Park, J.H.; Kim, Y.G.; Shaw, M.; Kanneganti, T.D.; Fujimoto, Y.; Fukase, K.; Inohara, N.; Núñez, G. Nod1/RICK and TLR signaling regulate chemokine and antimicrobial innate immune responses in mesothelial cells. J. Immunol 2007, 179, 514–521. [Google Scholar]
- Kato, S.; Yuzawa, Y.; Tsuboi, N.; Maruyama, S.; Morita, Y.; Matsuguchi, T.; Matsuo, S. Endotoxin-induced chemokine expression in murine peritoneal mesothelial cells: The role of toll-like receptor 4. J. Am. Soc. Nephrol 2004, 15, 1289–1299. [Google Scholar]
- Kawagoe, T.; Sato, S.; Matsushita, K.; Kato, H.; Matsui, K.; Kumagai, Y.; Saitoh, T.; Kawai, T.; Takeuchi, O.; Akira, S. Sequential control of Toll-like receptor-dependent responses by IRAK1 and IRAK2. Nat. Immunol 2008, 9, 684–691. [Google Scholar]
- O’Neill, L.A. When signaling pathways collide: Positive and negative regulation of Toll-like receptor signal transduction. Immunity 2008, 29, 12–20. [Google Scholar]
- Li, X. IRAK4 in TLR/IL-1R signaling: Possible clinical applications. Eur. J. Immunol 2008, 38, 614–618. [Google Scholar]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-κB signaling. Cell 2008, 132, 344–362. [Google Scholar]
- Lomaga, M.A.; Yeh, W.C.; Sarosi, I.; Duncan, G.S.; Furlonger, C.; Ho, A.; Morony, S.; Capparelli, C.; Van, G.; Kaufman, S.; et al. TRAF6 deficiency results in osteopetrosis and defective interleukin-1, CD40, and LPS signaling. Genes Dev 1999, 13, 1015–1024. [Google Scholar]
- Laman, J.D.; de Boer, M.; Hart, B.A. CD40 in clinical inflammation: From multiple sclerosis to atherosclerosis. Dev. Immunol 1998, 6, 215–222. [Google Scholar]
- Zhang, Y.F.; Yang, X.; Zhang, Y.J.; Sun, Y.L.; Zou, X.L.; Kong, Q.Y.; Dong, X.Q.; Ye, X.Q.; Yu, X.Q. Peroxisome proliferator-activated receptor-gamma is expressed by rat peritoneal mesothelial cells: Its potential role in peritoneal cavity local defense. Am. J. Nephrol 2006, 26, 602–611. [Google Scholar]
- Wu, J.; Yang, X.; Zhang, Y.F.; Zhou, S.F.; Zhang, R.; Dong, X.Q.; Fan, J.J.; Liu, M.; Yu, X.Q. Angiotensin II upregulates Toll-like receptor 4 and enhances lipopolysaccharide-induced CD40 expression in rat peritoneal mesothelial cells. Inflamm. Res 2009, 58, 473–482. [Google Scholar]
- Rowland, S.L.; Tremblay, M.M.; Ellison, J.M.; Stunz, L.L.; Bishop, G.A.; Hostager, B.S. A novel mechanism for TNFR-associated factor 6-dependent CD40 signaling. J. Immunol 2007, 179, 4645–4653. [Google Scholar]
- Vereecke, L.; Beyaert, R.; van Loo, G. The ubiquitin-editing enzyme A20 (TNFAIP3) is a central regulator of immunopathology. Trends Immunol 2009, 30, 383–391. [Google Scholar]
- Coornaert, B.; Carpentier, I.; Beyaert, R. A20: Central gatekeeper in inflammation and immunity. J. Biol. Chem 2009, 284, 8217–8221. [Google Scholar]
- Lee, E.G.; Boone, D.L.; Chai, S.; Libby, S.L.; Chien, M.; Lodolce, J.P.; Ma, A. Failure to regulate TNF-induced NF-kappaB and cell death responses in A20-deficient mice. Science 2000, 289, 2350–2354. [Google Scholar]
- Kang, N.I.; Yoon, H.Y.; Lee, Y.R.; Won, M.; Chung, M.J.; Park, J.W.; Hur, G.M.; Lee, H.K.; Park, B.H. A20 attenuates allergic airway inflammation in mice. J. Immunol 2009, 183, 1488–1495. [Google Scholar]
- Hah, Y.S.; Lee, Y.R.; Jun, J.S.; Lim, H.S.; Kim, H.O.; Jeong, Y.G.; Hur, G.M.; Lee, S.Y.; Chung, M.J.; Park, J.W.; et al. A20 suppresses inflammatory responses and bone destruction in human fibroblast-like synoviocytes and in mice with collagen-induced arthritis. Arthritis Rheum 2010, 62, 2313–2321. [Google Scholar]
- Gui, J.; Yue, Y.; Chen, R.; Xu, W.; Xiong, S. A20 (TNFAIP3) alleviates CVB3-induced myocarditis via inhibiting NF-κB signaling. PLoS One 2012, 7, e46515. [Google Scholar]
- Loniewski, K.J.; Patial, S.; Parameswaran, N. Sensitivity of TLR4- and -7-induced NFkappa B1 p105-TPL2-ERK pathway to TNF-receptor-associated-factor-6 revealed by RNAi in mouse macrophages. Mol. Immunol 2007, 44, 3715–3723. [Google Scholar]
- Oshima, N.; Ishihara, S.; Rumi, M.A.; Aziz, M.M.; Mishima, Y.; Kadota, C.; Moriyama, I.; Ishimura, N.; Amano, Y.; Kinoshita, Y. A20 is an early responding negative regulator of Toll-like receptor 5 signalling in intestinal epithelial cells during inflammation. Clin. Exp. Immunol 2010, 159, 185–198. [Google Scholar]
- Turer, E.E.; Tavares, R.M.; Mortier, E.; Hitotsumatsu, O.; Advincula, R.; Lee, B.; Shifrin, N.; Malynn, B.A.; Ma, A. Homeostatic MyD88 dependent signals cause lethal inflammation in the absence of A20. J. Exp. Med 2008, 205, 451–464. [Google Scholar]
- Werner, S.L.; Kearns, J.D.; Zadorozhnaya, V.; Lynch, C.; O’Dea, E.; Boldin, M.P.; Ma, A.; Baltimore, D.; Hoffmann, A. Encoding NF-kappaB temporal control in response to TNF: Distinct roles for the negative regulators IkappaBalpha and A20. Genes Dev 2008, 22, 2093–2101. [Google Scholar]
- Xiong, Y.; Qiu, F.; Piao, W.; Song, C.; Wahl, L.M.; Medvedev, A.E. Endotoxin tolerance impairs IL-1 receptor-associated kinase (IRAK) 4 and TGF-beta-activated kinase 1 activation, K63-linked polyubiquitination and assembly of IRAK1, TNF receptor-associated factor 6, and IkappaB kinasegamma and increases A20 expression. J. Biol. Chem 2011, 286, 7905–7916. [Google Scholar]
- Chen, Z.J. Ubiquitin signalling in the NF-kappaB pathway. Nat. Cell Biol 2005, 7, 758–765. [Google Scholar]
- Jakus, P.B.; Kalman, N.; Antus, C.; Radnai, B.; Tucsek, Z.; Gallyas, F.; Sumegi, B., Jr.; Veres, B. TRAF6 is functional in inhibition of TLR4-mediated NF-κB activation by resveratrol. J. Nutr. Biochem 2013, 24, 819–823. [Google Scholar]
- Mabilleau, G.; Chappard, D.; Sabokbar, A. Role of the A20-TRAF6 axis in lipopolysaccharide-mediated osteoclastogenesis. J. Biol. Chem 2011, 286, 3242–3249. [Google Scholar]
- Shi, C.S.; Kehrl, J.H. Traf6 and A20 differentially regulate TLR4-induced autophagy by affecting the ubiquitination of Beclin 1. Autophagy 2010, 6, 986–987. [Google Scholar]
- Komander, D.; Barford, D. Structure of the A20 OTU domain and mechanistic insights into deubiquitination. Biochem. J 2008, 409, 77–85. [Google Scholar]
- Lin, S.C.; Chung, J.Y.; Lamothe, B.; Rajashankar, K.; Lu, M.; Lo, Y.C.; Lam, A.Y.; Darnay, B.G.; Wu, H. Molecular basis for the unique deubiquitinating activity of the NF-kappaB inhibitor A20. J. Mol. Biol 2008, 376, 526–540. [Google Scholar]
- Iha, H.; Peloponese, J.M.; Verstrepen, L.; Zapart, G.; Ikeda, F.; Smith, C.D.; Starost, M.F.; Yedavalli, V.; Heyninck, K.; Dikic, I.; et al. Inflammatory cardiac valvulitis in TAX1BP1-deficient mice through selective NF-kappaB activation. EMBO J 2008, 27, 629–641. [Google Scholar]
- Glik, A.; Mazar, J.; Rogachev, B.; Zlotnik, M.; Douvdevani, A. CD40 ligand expression correlates with resolution of peritonitis and mononuclear cell recruitment. Perit. Dial. Int 2005, 25, 240–247. [Google Scholar]
- Tavares, R.M.; Turer, E.E.; Liu, C.L.; Advincula, R.; Scapini, P.; Rhee, L.; Barrera, J.; Lowell, C.A.; Utz, P.J.; Malynn, B.A.; et al. The ubiquitin modifying enayme A20 restricts B cell survival and prevents autoimmunity. Immunity 2010, 33, 181–191. [Google Scholar]
- Wajant, H.; Henkler, F.; Scheurich, P. The TNF-receptor-associated factor family: Scaffold mole-cules for cytokine receptors, kinases and theirregulators. Cell Signal 2001, 13, 389–400. [Google Scholar]
- Aggarwal, B.B. Signalling pathways of the TNF superfamily: A double-edged sword. Nat. Rev. Immunol 2003, 3, 745–756. [Google Scholar]
- Chung, J.Y.; Park, Y.C.; Ye, H.; Wu, H. All TRAFs are not created equal: Common and distinct molecular mechanisms of TRAF-mediated signal transduction. J. Cell Sci 2002, 115, 679–688. [Google Scholar]
- Bishop, G.A.; Hostager, B.S.; Brown, K.D. Mechanisms of TNF receptor-associated factor (TRAF) regulation in B lymphocytes. J. Leukoc. Biol 2002, 72, 19–23. [Google Scholar]
- Ishida, T.; Mizushima, S.; Azuma, S.; Kobayashi, N.; Tojo, T.; Suzuki, K.; Aizawa, S.; Watanabe, T.; Mosialos, G.; Kieff, E.; et al. Identification of TRAF6, a novel tumor necrosis factor receptor-associated factor protein that mediates signaling from an amino-terminal domain of the CD40 cytoplasmic region. J. Biol. Chem 1996, 271, 28745–28748. [Google Scholar]
- Qin, H.; Wilson, C.A.; Lee, S.J.; Zhao, X.; Benveniste, E.N. LPS induces CD40 gene expression through the activation of NF-kappaB and STAT-1alpha in macrophages and microglia. Blood 2005, 106, 3114–3122. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer sequence |
|---|---|
| TRAF6 | Forward: 5′-GATCGGGTTGTGTGTGTCTG-3′ Reverse: 5′-AGAGACACCCCAGCAGCTAA-3′ |
| CD40 | Forward: 5′-GAATTCTCAGCCCAGTGGAA-3′ Reverse: 5′-GCAGGGATGACAGACGGTAT-3′ |
| IL-6 | Forward: 5′-AGTTGCCTTCTTGGGACTGA-3′ Reverse: 5′-CAGAATTGCCATTGCACAAC-3′ |
| TNF-α | Forward: 5′-GATTATGGCTCAGGGTCCAA-3′ Reverse: 5′-CTCCCTTTGCAGAACTCAGG-3′ |
| GAPDH | Forward: 5′-AACTTTGGCATTGTGGAAGG-3′ Reverse: 5′-CACATTGGGGGTAGGAACAC-3′ |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zou, X.-L.; Pei, D.-A.; Yan, J.-Z.; Xu, G.; Wu, P. A20 Overexpression Inhibits Lipopolysaccharide-Induced NF-κB Activation, TRAF6 and CD40 Expression in Rat Peritoneal Mesothelial Cells. Int. J. Mol. Sci. 2014, 15, 6592-6608. https://doi.org/10.3390/ijms15046592
Zou X-L, Pei D-A, Yan J-Z, Xu G, Wu P. A20 Overexpression Inhibits Lipopolysaccharide-Induced NF-κB Activation, TRAF6 and CD40 Expression in Rat Peritoneal Mesothelial Cells. International Journal of Molecular Sciences. 2014; 15(4):6592-6608. https://doi.org/10.3390/ijms15046592
Chicago/Turabian StyleZou, Xun-Liang, De-An Pei, Ju-Zhen Yan, Gang Xu, and Ping Wu. 2014. "A20 Overexpression Inhibits Lipopolysaccharide-Induced NF-κB Activation, TRAF6 and CD40 Expression in Rat Peritoneal Mesothelial Cells" International Journal of Molecular Sciences 15, no. 4: 6592-6608. https://doi.org/10.3390/ijms15046592