Gentamicin Blocks the ACh-Induced BK Current in Guinea Pig Type II Vestibular Hair Cells by Competing with Ca2+ at the l-Type Calcium Channel


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
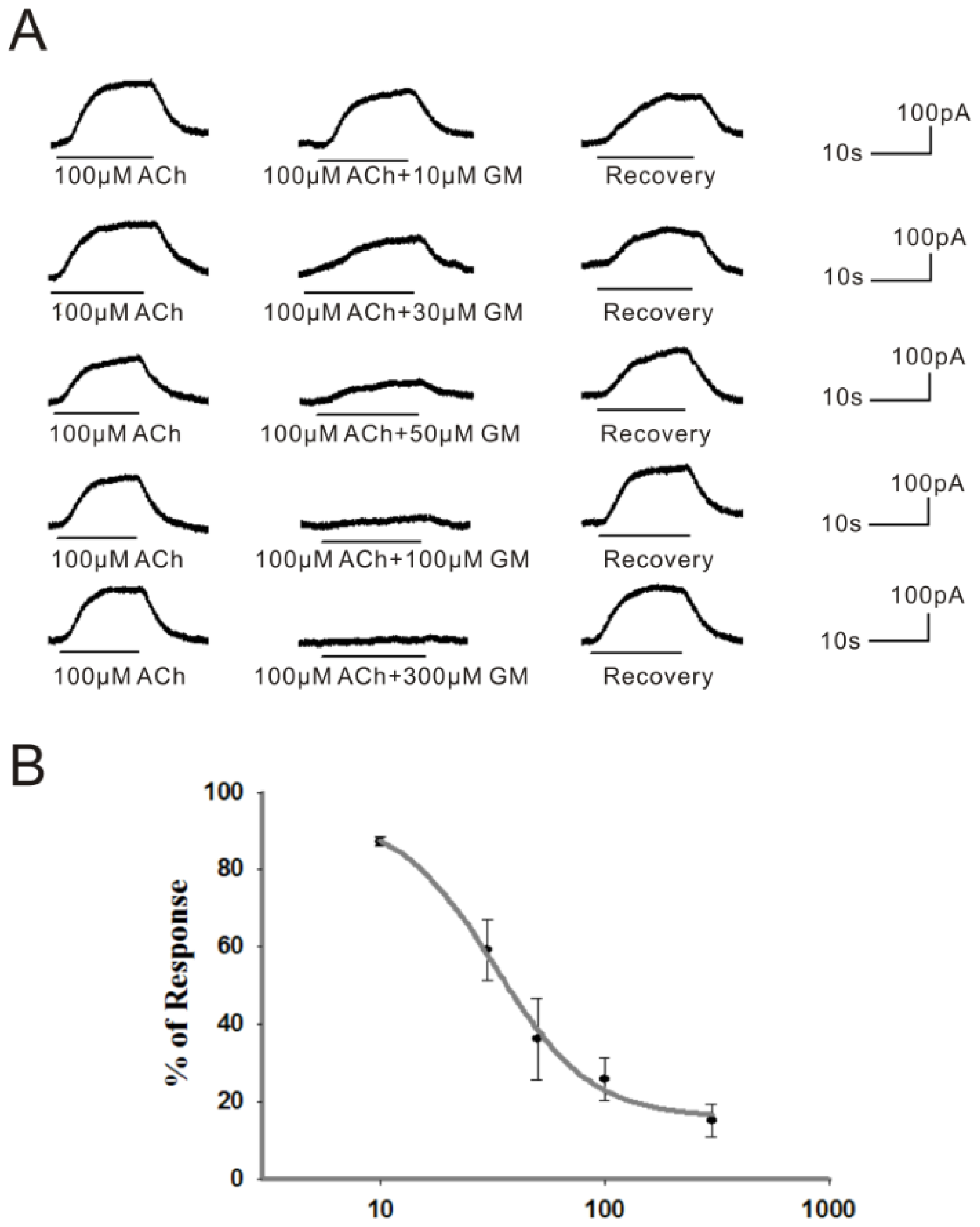
2.1. GM Reversibly Blocked the ACh-Induced BK Current in Guinea Pig VHCs II in a Dose-Dependent and Voltage-Independent Manner
2.2. Inhibition of GM Is not Affected by ACh Concentration
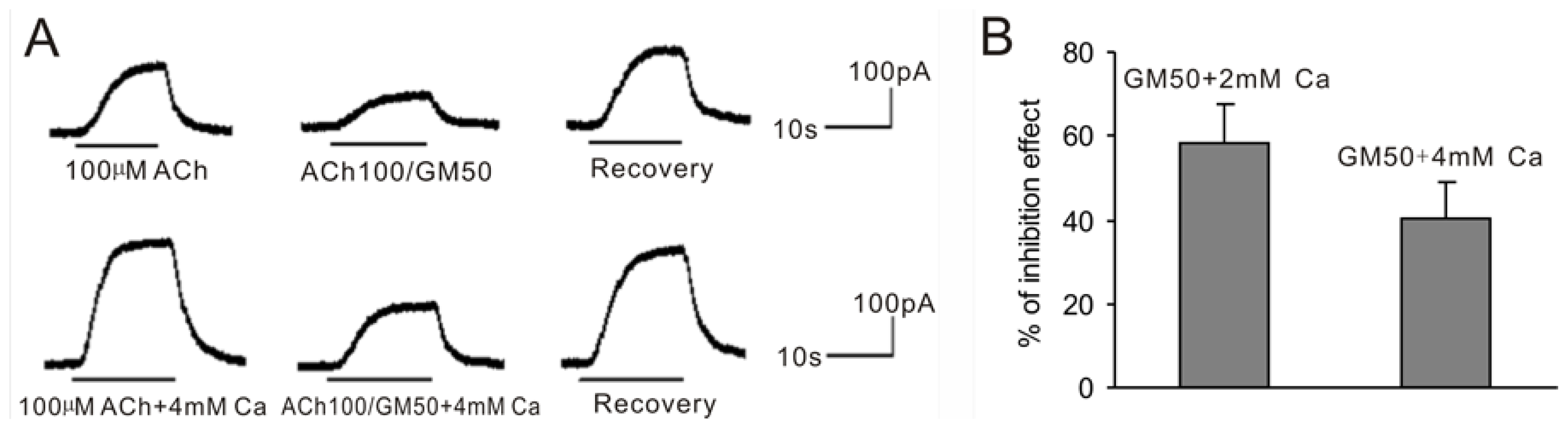
2.3. Increasing the Extracellular Ca2+ Concentration Antagonizes GM Inhibition and GM Can Block Ca2+ Evoked by (−)-Bay-K8644
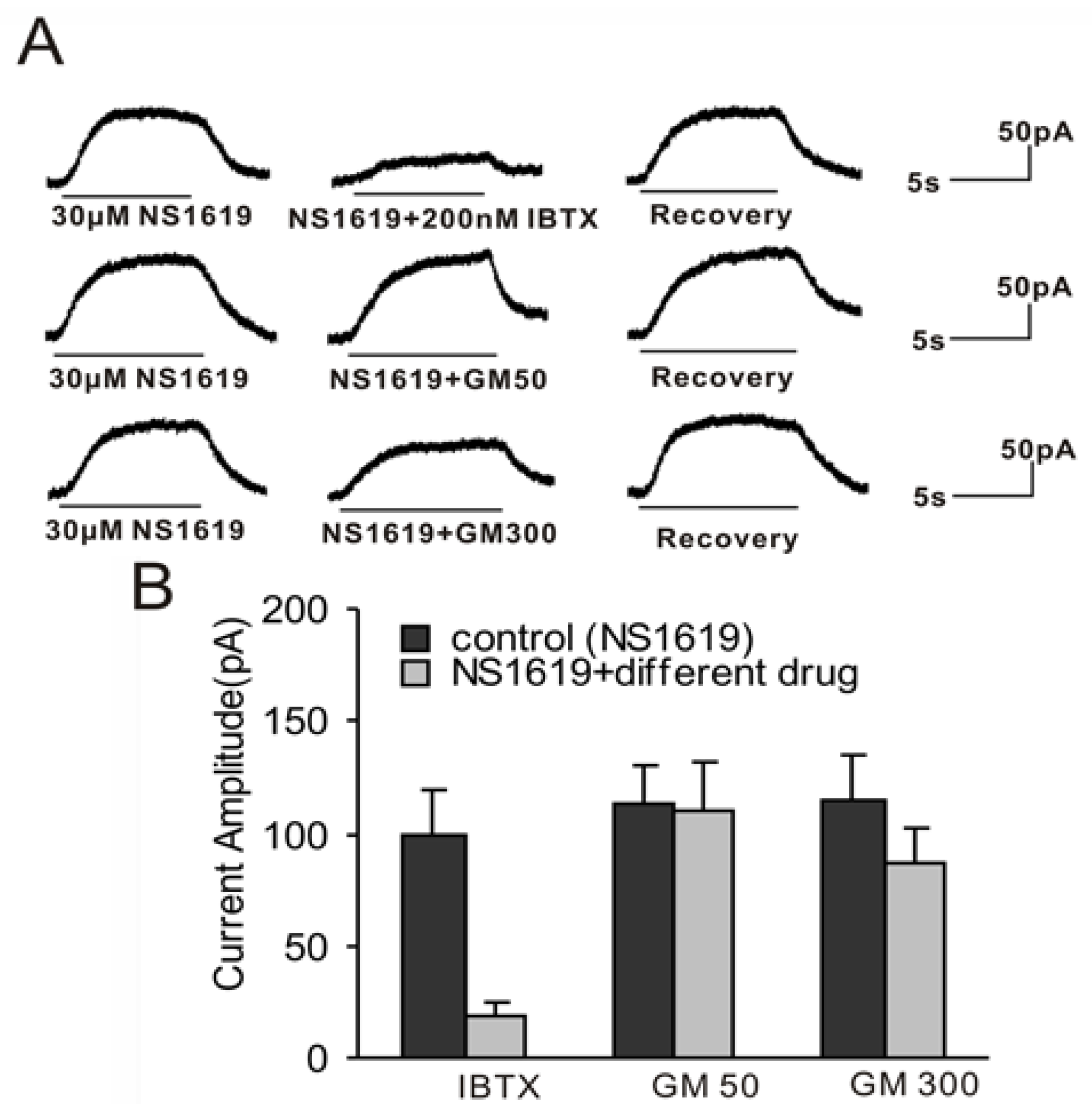
2.4. BK Current Evoked by NS1619 Insensitive to 50 μM GM and Only Slightly Blocked by 300 μM GM
3. Discussion
4. Methods
4.1. Ethics Statement
4.2. Animal Procedures and VHCs II Preparation
4.3. Electrophysiology
4.4. Drug Application
4.5. Data Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Webster, J.C.; McGee, T.M.; Carroll, R.; Benitez, J.T.; Williams, M.L. Ototoxicity of gentamicin. Histopathologic and functional results in the cat. Trans. Am. Acad. Ophthalmol. Otolaryngol 1970, 74, 1155–1165. [Google Scholar]
- Blakley, B.W. Update on intratympanic gentamicin for Ménière’s disease. Laryngoscope 2000, 110, 236–240. [Google Scholar]
- Straube, A. Pharmacology of vertigo/nystagmus/oscillopsia. Curr. Opin. Neurol 2005, 18, 11–14. [Google Scholar]
- Dobrev, D.; Ravens, U. Therapeutically relevant concentrations of neomycin selectively inhibit P-type Ca2+ channels in rat striatum. Eur. J. Pharmacol 2003, 461, 105–111. [Google Scholar]
- Fiekers, J.F. Effects of the aminoglycoside antibiotics, streptomycin and neomycin, on neuromuscular transmission. II. Postsynaptic considerations. J. Pharmacol. Exp. Ther 1983, 225, 496–502. [Google Scholar]
- Parsons, T.D.; Obaid, A.L.; Salzberg, B.M. Aminoglycoside antibiotics block voltage-dependent calcium channels in intact vertebrate nerve terminals. J. Gen. Physiol 1992, 99, 491–504. [Google Scholar]
- Pichler, M.; Wang, Z.; Grabner-Weiss, C.; Reimer, D.; Hering, S.; Grabner, M.; Glossmann, H.; Striessnig, J. Block of P/Q-type calcium channels by therapeutic concentrations of aminoglycoside antibiotics. Biochemistry 1996, 35, 14659–14664. [Google Scholar]
- Kroese, A.B.; Das, A.; Hudspeth, A.J. Blockage of the transduction channels of hair cells in the bullfrog’s sacculus by aminoglycoside antibiotics. Hear Res 1989, 37, 203–217. [Google Scholar]
- Marcotti, W.; Netten, S.M.V.; Kros, C.J. The aminoglycoside antibiotic dihydrostreptomycin rapidly enters mouse outer hair cells through the mechano-electrical transducer channels. J. Physiol 2005, 567, 505–521. [Google Scholar]
- Ohmori, H. Mechano-electrical transduction currents in isolated vestibular hair cells of the chick. J. Physiol 1985, 359, 189–217. [Google Scholar]
- Wackym, P.A.; Chen, C.T.; Ishiyama, A.; Pettis, R.M.; López, I.A.; Hoffman, L. Muscarinic acetylcholine receptor subtype mRNAs in the human and rat vestibular periphery. Cell Biol. Int 1996, 20, 187–192. [Google Scholar]
- Winegar, B.D.; Haws, C.M.; Lansman, J.B. Subconductance block of single mechanosensitive ion channels in skeletal muscle fibers by aminoglycoside antibiotics. J. Gen. Physiol 1996, 107, 433–443. [Google Scholar]
- Amici, M.; Eusebi, F.; Miledi, R. Effects of the antibiotic gentamicin on nicotinic acetylcholine receptors. Neuropharmacology 2005, 49, 627–637. [Google Scholar]
- Blanchet, C.; Erostequi, C.; Suqasawa, M.; Dulon, D. Gentamicin blocks ACh-evoked K+ current in guinea-pig outer hair cells by impairing Ca2+ entry at the cholinergic receptor. J. Physiol 2000, 525, 641–654. [Google Scholar]
- Rothlin, C.V.; Katz, E.; Verbotsky, M.; Vetter, D.E.; Heinemann, S.F.; Elqoyhen, A.B. Block of the alpha9 nicotinic receptor by ototoxic aminoglycosides. Neuropharmacology 2000, 39, 2525–2532. [Google Scholar]
- Smith, D.W.; Erre, J.P.; Aran, J.M. Rapid, reversible elimination of medial olivocochlear efferent function following single injections of gentamicin in the guinea pig. Brain Res 1994, 652, 243–248. [Google Scholar]
- Yoshida, N.; Liberman, M.C.; Brown, M.C.; Sewell, W.F. Gentamicin blocks both fast and slow effects of olivocochlear activation in anesthetized guinea pigs. J. Neurophysiol 1999, 82, 3168–3174. [Google Scholar]
- Mulders, W.H.; Robertson, D. Gentamicin abolishes all cochlear effects of electrical stimulation of the inferior colliculus. Exp. Brain Res 2006, 174, 35–44. [Google Scholar]
- Avan, P.; Erre, J.P.; da Costa, D.L.; Aran, J.M.; Popelár, J. The efferent-mediated suppression of otoacoustic emissions in awake guinea pigs and its reversible blockage by gentamicin. Exp. Brain Res 1996, 109, 9–16. [Google Scholar]
- Martini, M.; Canella, R.; Priqioni, I.; Russo, G.; Tavazzani, E.; Fesce, R.; Rossi, M.L. Acute effects of gentamicin on the ionic currents of semicircular canal hair cells in the frog. Hear Res 2011, 282, 151–160. [Google Scholar]
- Guth, P.S.; Perin, P.; Norris, C.H.; Valli, P. The vestibular hair cells: Post-transductional signal processing. Prog. Neurobiol 1998, 54, 193–247. [Google Scholar]
- Ishiyama, A.; Lopez, I.; Wackym, P.A. Molecular characterization of muscarinic receptors in the human vestibular periphery. Implications for pharmacotherapy. Am. J. Otol 1997, 18, 648–654. [Google Scholar]
- Yao, Q.; Cheng, H.; Guo, C.; Zhou, T.; Huang, X.; Kong, W. Muscarinic acetylcholine receptor subtype expression in type II vestibular hair cells of guinea pigs. J. Huazhong Uni. Sci. Technol 2011, 31, 682–686. [Google Scholar]
- Anderson, A.D.; Troyanovskaya, M.; Wackym, P.A. Differential expression of alpha2–7, alpha9 and beta2–4 nicotinic acetylcholine receptor subunit mRNA in the vestibular end-organs and Scarpa’s ganglia of the rat. Brain Res 1997, 778, 409–413. [Google Scholar]
- Elgoyhen, A.B.; Vetter, D.E.; Katz, E.; Rothlin, C.V.; Heinemann, S.F.; Boulter, J. alpha10: A determinant of nicotinic cholinergic receptor function in mammalian vestibular and cochlear mechanosensory hair cells. Proc. Natl. Acad. Sci. USA 2001, 98, 3501–3506. [Google Scholar]
- Simmons, D.D.; Morley, B.J. Spatial and temporal expression patterns of nicotinic acetylcholine alpha9 and alpha10 subunits in the embryonic and early postnatal inner ear. Neuroscience 2011, 194, 326–336. [Google Scholar]
- Guo, C.K.; Wang, Y.; Zhou, T.; Yu, H.; Zhang, W.J.; Kong, W.J. M2 muscarinic ACh receptors sensitive BK channels mediate cholinergic inhibition of type II vestibular hair cells. Hear Res 2012, 285, 13–19. [Google Scholar]
- Kong, W.J.; Guo, C.K.; Zhang, S.; Hao, J.; Wang, Y.J.; Li, Z.W. The properties of ACh-induced BK currents in guinea pig type II vestibular hair cells. Hear Res 2005, 209, 1–9. [Google Scholar]
- Kong, W.J.; Guo, C.K.; Zhang, X.W.; Chen, X.; Zhang, S.; Li, G.Q.; Li, Z.W.; van Cauwenberqe, P. The coupling of acetylcholine-induced BK channel and calcium channel in guinea pig saccular type II vestibular hair cells. Brain Res 2007, 1129, 110–115. [Google Scholar]
- Wanamaker, H.H.; Gruenwald, L.; Damm, K.J.; Oqata, Y.; Slepecky, N. Dose-related vestibular and cochlear effects of transtympanic gentamicin. Am. J. Otol 1998, 19, 170–179. [Google Scholar]
- Dulon, D.; Zajic, G.; Aran, J.M.; Schacht, J. Aminoglycoside antibiotics impair calcium entry but not viability and motility in isolated cochlear outer hair cells. J. Neurosci. Res 1989, 24, 338–346. [Google Scholar]
- Tan, C.T.; Lee, S.Y.; Yao, C.J.; Liu, S.H.; Lin-Shiau, S.Y. Effects of gentamicin and pH on [Ca2+]i in apical and basal outer hair cells from guinea pigs. Hear Res 2001, 154, 81–87. [Google Scholar]
- Zhou, T.; Wang, Y.; Guo, C.K.; Zhang, W.J.; Yu, H.; Zhang, K.; Kong, W.J. Two distinct channels mediated by m2 mAChR and α9nAChR co-exist in type II vestibular hair cells of guinea pig. Int. J. Mol. Sci 2013, 14, 8818–8831. [Google Scholar]
- Liu, S.Q.; Kaczmarek, L.K. Aminoglycosides block the Kv3.1 potassium channel and reduce the ability of inferior colliculus neurons to fire at high frequencies. J. Neurobiol 2005, 62, 439–452. [Google Scholar]
- Nomura, K.; Naruse, K.; Watanabe, K.; Sokabe, M. Aminoglycoside blockade of Ca2+-activated K+ channel from rat brain synaptosomal membranes incorporated into planar bilayers. J. Membr. Biol 1990, 115, 241–251. [Google Scholar]
- Scott, E.M.; Johnson, M.; Meredith, F.L.; Rennie, K.J. Inhibition of K+ Currents in Type I Vestibular Hair Cells by Gentamicin and Neomycin. Audiol. Neurotol 2013, 18, 317–326. [Google Scholar]
- Scarfone, E.; Ulfendahl, M.; Löfstrand, P.; Flock, A. Light- and electron microscopy of isolated vestibular hair cells from the guinea pig. Cell Tissue Res 1991, 266, 51–58. [Google Scholar]
- Lopez, I.; Ayala, C.; Honrubia, V. Synaptophysin immunohistochemistry during vestibular hair cell recovery after gentamicin treatment. Audiol. Neurootol 2003, 8, 80–90. [Google Scholar]









© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yu, H.; Guo, C.-K.; Wang, Y.; Zhou, T.; Kong, W.-J. Gentamicin Blocks the ACh-Induced BK Current in Guinea Pig Type II Vestibular Hair Cells by Competing with Ca2+ at the l-Type Calcium Channel. Int. J. Mol. Sci. 2014, 15, 6757-6771. https://doi.org/10.3390/ijms15046757
Yu H, Guo C-K, Wang Y, Zhou T, Kong W-J. Gentamicin Blocks the ACh-Induced BK Current in Guinea Pig Type II Vestibular Hair Cells by Competing with Ca2+ at the l-Type Calcium Channel. International Journal of Molecular Sciences. 2014; 15(4):6757-6771. https://doi.org/10.3390/ijms15046757
Chicago/Turabian StyleYu, Hong, Chang-Kai Guo, Yi Wang, Tao Zhou, and Wei-Jia Kong. 2014. "Gentamicin Blocks the ACh-Induced BK Current in Guinea Pig Type II Vestibular Hair Cells by Competing with Ca2+ at the l-Type Calcium Channel" International Journal of Molecular Sciences 15, no. 4: 6757-6771. https://doi.org/10.3390/ijms15046757