A New Type I Peritrophic Membrane Protein from Larval Holotrichia oblita (Coleoptera: Melolonthidae) Binds to Chitin


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
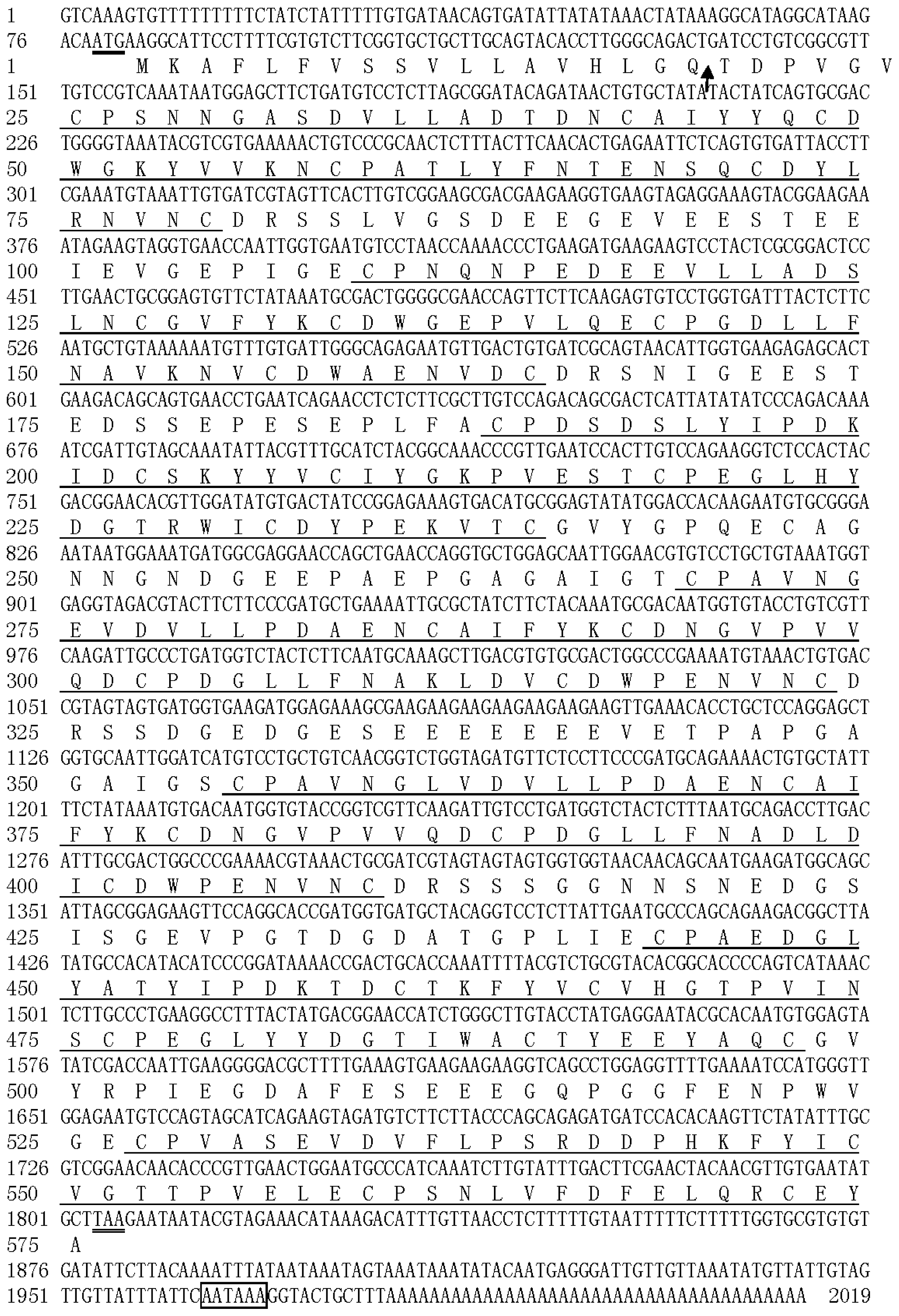
2.1. Identification of the cDNA Coding for a New H. oblita PM Protein, HoCBP76
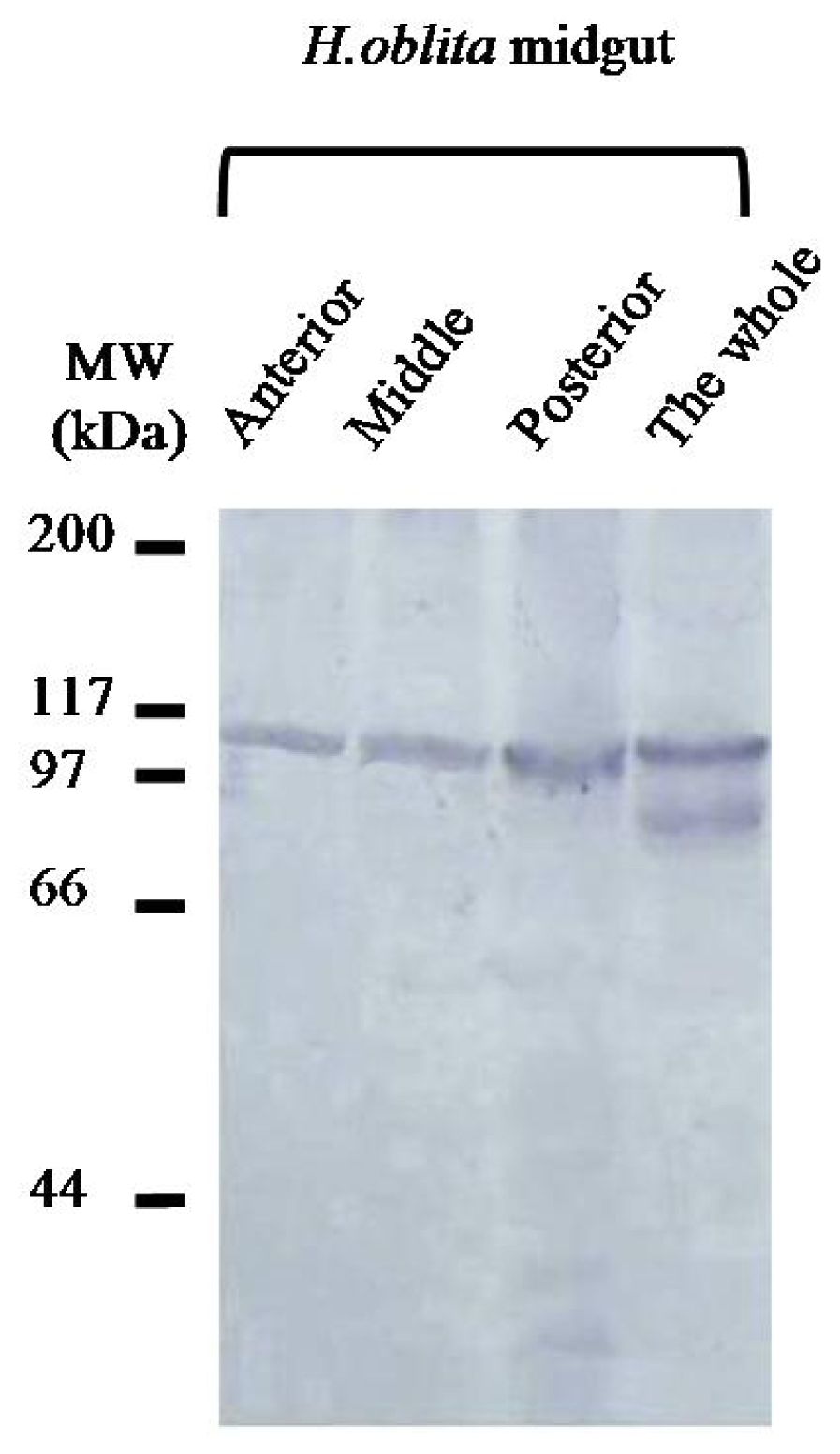
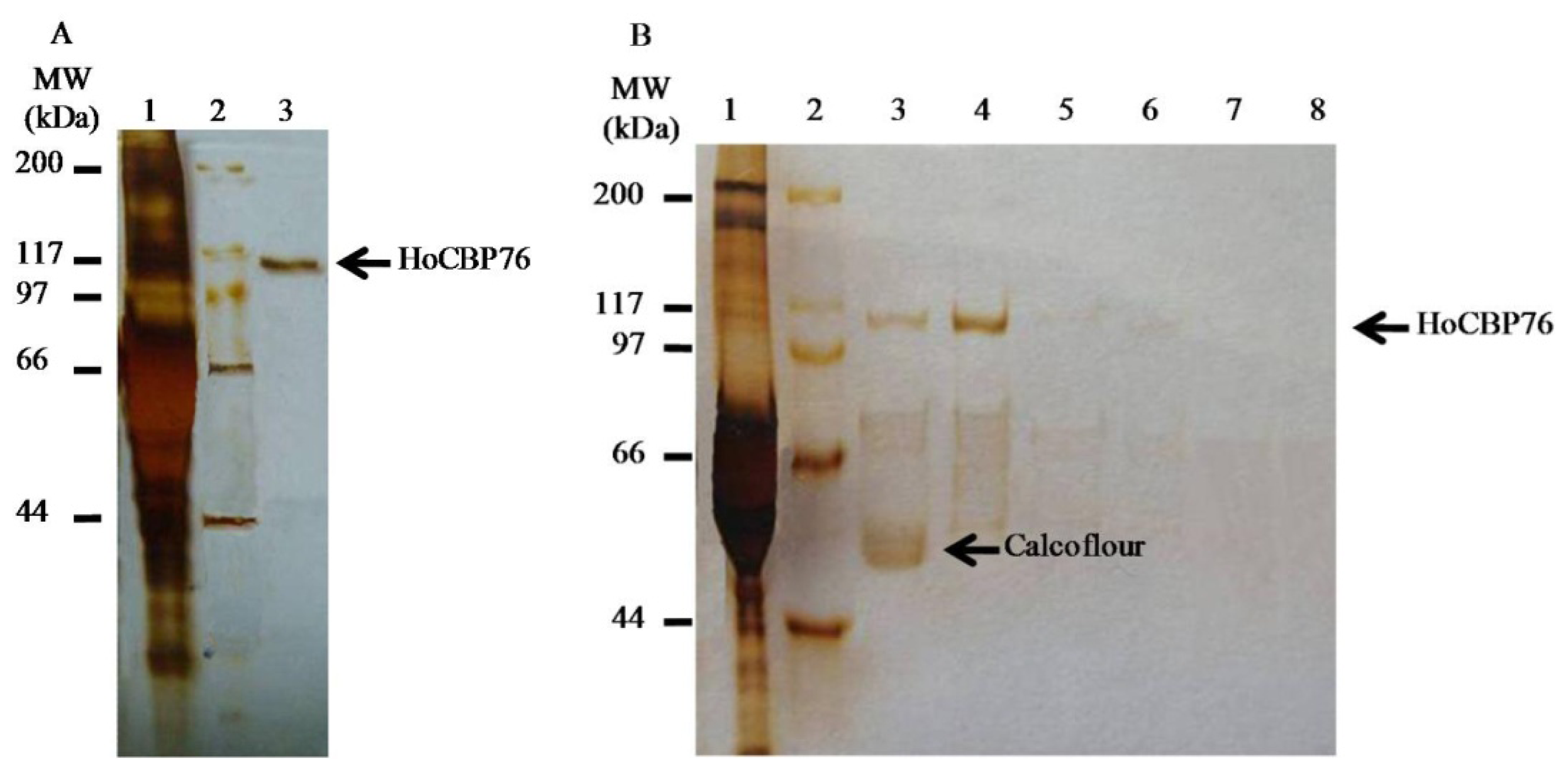
2.2. Identification of HoCBP76 from the H. oblita PM
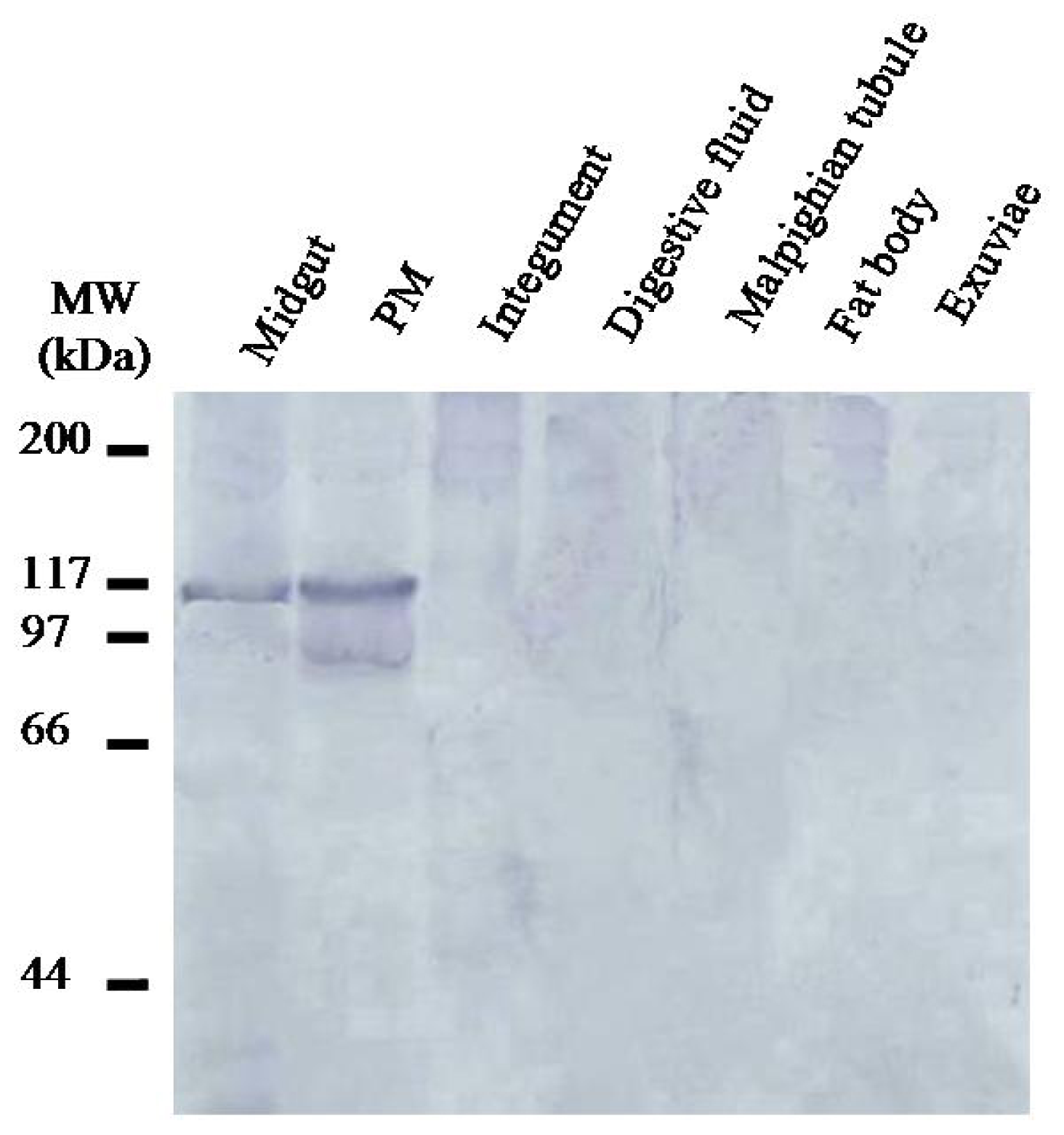
2.3. Localization of HoCBP76 in H. oblita Larvae
2.4. Binding of Recombinant HoCBP76 to Chitin
3. Experimental Section
3.1. Insect Larvae and Collection of Larval Tissues
3.2. Construction and Immunoscreening of a cDNA Expression Library
3.3. Recombinant HoCBP76 Expression
3.4. Regenerated Chitin Preparation and Chitin-Binding Assay
3.5. Preparation of Antibodies Reacting to HoCBP76
3.6. Immunolocalization of HoCBP76 in H. oblita Larvae
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Peters, W. Peritrophic Membranes; Springer-Verlag: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Tellam, R.L.; Wijffels, G.; Willadsen, P. Peritrophic matrix proteins. Insect Biochem. Mol. Biol 1999, 29, 87–101. [Google Scholar]
- Hegedus, D.; Erlandson, M.; Gillott, C.; Toprak, U. New insights into peritrophic matrix synthesis, architecture, and function. Annu. Rev. Entomol 2009, 54, 285–302. [Google Scholar]
- Shi, X.; Chamankhah, M.; Visal-Shah, S.; Hemmingsen, S.M.; Erlandson, M.; Braun, L.; Alting-Mees, M.; Khachatourians, G.G.; O’Grady, M.; Hegedus, D.D. Modeling the structure of the Type I peritrophic matrix: Characterization of a Mamestra configurata intestinal mucin and a novel peritrophin containing 19 chitin binding domains. Insect Biochem. Mol. Biol 2004, 34, 1101–1115. [Google Scholar]
- Wang, P.; Granados, R.R. Molecular cloning and sequencing of a novel invertebrate intestinal mucin cDNA. J. Biol. Chem 1997, 272, 16663–16669. [Google Scholar]
- Terra, W.R. The origin and functions of the insect peritrophic membrane and peritrophic gel. Arch. Insect Biochem. Physiol 2001, 47, 47–61. [Google Scholar]
- Yin, J.; Wei, Z.-J.; Li, K.-B.; Cao, Y.-Z.; Guo, W. Identification and molecular characterization of a new member of the peritrophic membrane proteins from the meadow moth, loxostege sticticalis. Int. J. Biol. Sci 2010, 6, 491–498. [Google Scholar]
- Shao, L.; Devenport, M.; Jacobs-Lorena, M. The peritrophic matrix of hematophagous insects. Arch. Insect Biochem. Physiol 2001, 47, 119–125. [Google Scholar]
- Devenport, M.; Fujioka, H.; Donnelly-Doman, M.; Shen, Z.; Jacobs-Lorena, M. Storage and secretion of Ag-Aper14, a novel peritrophic matrix protein, and Ag-Muc1 from the mosquito Anopheles gambiae. Cell Tissue Res 2005, 320, 175–185. [Google Scholar]
- Ryerse, J.; Purcell, J.; Sammons, R. Structure and formation of the peritrophic membrane in the larva of the southern corn rootwormDiabrotica undecimpunctata. Tissue Cell 1994, 26, 431–437. [Google Scholar]
- Tristram, J.N. Normal and cocoon-forming peritrophic membrane in larvae of the beetleGibbium psylloides. J. Insect Physiol 1977, 23, 79–87. [Google Scholar]
- Wijffels, G.; Eisemann, C.; Riding, G.; Pearson, R.; Jones, A.; Willadsen, P.; Tellam, R. A novel family of chitin-binding proteins from insect type 2 peritrophic matrix. J. Biol. Chem 2001, 276, 15527–15536. [Google Scholar]
- Lehane, M. Peritrophic matrix structure and function. Annu. Rev. Entomol 1997, 42, 525–550. [Google Scholar]
- Tellam, R.L.; Eisemann, C. Chitin is only a minor component of the peritrophic matrix from larvae of Lucilia cuprina. Insect Biochem. Mol. Biol 2000, 30, 1189–1201. [Google Scholar]
- Shen, Z.; Jacobs-Lorena, M. Evolution of chitin-binding proteins in invertebrates. J. Mol. Evol 1999, 48, 341–347. [Google Scholar]
- Elvin, C.M.; Vuocolo, T.; Pearson, R.D.; East, I.J.; Riding, G.A.; Eisemann, C.H.; Tellam, R.L. Characterization of a major peritrophic membrane protein, peritrophin-44, from the larvae of Lucilia cuprina. J. Biol. Chem 1996, 271, 8925–8935. [Google Scholar]
- Zhu, X.; Zhang, H.; Fukamizo, T.; Muthukrishnan, S.; Kramer, K.J. Properties of Manduca sexta chitinase and its C-terminal deletions. Insect Biochem. Mol. Biol 2001, 31, 1221–1230. [Google Scholar]
- Arakane, Y.; Zhu, Q.; Matsumiya, M.; Muthukrishnan, S.; Kramer, K.J. Properties of catalytic, linker and chitin-binding domains of insect chitinase. Insect Biochem. Mol. Biol 2003, 33, 631–648. [Google Scholar]
- Dyrløv Bendtsen, J.; Nielsen, H.; von Heijne, G.; Brunak, S. Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol 2004, 340, 783–795. [Google Scholar]
- Hansen, J.E.; Lund, O.; Tolstrup, N.; Gooley, A.A.; Williams, K.L.; Brunak, S. NetOglyc: Prediction of mucin type O-glycosylation sites based on sequence context and surface accessibility. Glycoconj. J 1998, 15, 115–130. [Google Scholar]
- Blom, N.; Sicheritz-Pontén, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics 2004, 4, 1633–1649. [Google Scholar]
- Jasrapuria, S.; Arakane, Y.; Osman, G.; Kramer, K.J.; Beeman, R.W.; Muthukrishnan, S. Genes encoding proteins with peritrophin A-type chitin-binding domains in Tribolium castaneum are grouped into three distinct families based on phylogeny, expression and function. Insect Biochem. Mol. Biol 2010, 40, 214–227. [Google Scholar]
- Gaines, P.J.; Walmsley, S.J.; Wisnewski, N. Cloning and characterization of five cDNAs encoding peritrophin-A domains from the cat fleaCtenocephalides felis. Insect Biochem. Mol. Biol 2003, 33, 1061–1073. [Google Scholar]
- Suetake, T.; Tsuda, S.; Kawabata, S.-I.; Miura, K.; Iwanaga, S.; Hikichi, K.; Nitta, K.; Kawano, K. Chitin-binding proteins in invertebrates and plants comprise a common chitin-binding structural motif. J. Biol. Chem 2000, 275, 17929–17932. [Google Scholar]
- Guo, W.; Li, G.; Pang, Y.; Wang, P. A novel chitin-binding protein identified from the peritrophic membrane of the cabbage looperTrichoplusia ni. Insect Biochem. Mol. Biol 2005, 35, 1224–1234. [Google Scholar]
- Wang, P.; Li, G.; Granados, R.R. Identification of two new peritrophic membrane proteins from larval Trichoplusia ni: Structural characteristics and their functions in the protease rich insect gut. Insect Biochem. Mol. Biol 2004, 34, 215–227. [Google Scholar]
- Sousa, M.E.C.; Santos, F.A.; Wanderley-Teixeira, V.; Teixeira, Á.A.; de Siqueira, H.Á.A.; Alves, L.C.; Torres, J.B. Histopathology and ultrastructure of midgut of Alabama argillacea (Hübner) (Lepidoptera: Noctuidae) fed Bt-cotton. J. Insect Physiol 2010, 56, 1913–1919. [Google Scholar]
- Michiels, K.; van Damme, E.J.; Smagghe, G. Plant-insect interactions: What can we learn from plant lectins? Arch. Insect Biochem. Physiol 2010, 73, 193–212. [Google Scholar]
- Yang, H.J.; Zhou, F.; Malik, F.A.; Bhaskar, R.; Li, X.H.; Hu, J.B.; Sun, C.G.; Miao, Y.G. Identification and characterization of two chitin-binding proteins from the peritrophic membrane of the silkworm, Bombyx mori L. Arch. Insect Biochem. Physiol 2010, 75, 221–230. [Google Scholar]
- Dinglasan, R.; Devenport, M.; Florens, L.; Johnson, J.; McHugh, C.; Donnelly-Doman, M.; Carucci, D.; Yates, J.; Jacobs-Lorena, M. The Anopheles gambiae adult midgut peritrophic matrix proteome. Insect Biochem. Mol. Biol 2009, 39, 125–134. [Google Scholar]
- Shao, L.; Devenport, M.; Fujioka, H.; Ghosh, A.; Jacobs-Lorena, M. Identification and characterization of a novel peritrophic matrix protein, Ae-Aper50, and the microvillar membrane protein, AEG12, from the mosquitoAedes aegypti. Insect Biochem. Mol. Biol 2005, 35, 947–959. [Google Scholar]
- Wang, P.; Granados, R.R. Molecular structure of the peritrophic membrane (PM): Identification of potential PM target sites for insect control. Arch. Insect Biochem. Physiol 2001, 47, 110–118. [Google Scholar]
- Molano, J.; Durán, A.; Cabib, E. A rapid and sensitive assay for chitinase using tritiated chitin. Anal. Biochem 1977, 83, 648–656. [Google Scholar]





© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, X.; Li, J.; Guo, W.; Li, R.; Zhao, D.; Li, X. A New Type I Peritrophic Membrane Protein from Larval Holotrichia oblita (Coleoptera: Melolonthidae) Binds to Chitin. Int. J. Mol. Sci. 2014, 15, 6831-6842. https://doi.org/10.3390/ijms15046831
Liu X, Li J, Guo W, Li R, Zhao D, Li X. A New Type I Peritrophic Membrane Protein from Larval Holotrichia oblita (Coleoptera: Melolonthidae) Binds to Chitin. International Journal of Molecular Sciences. 2014; 15(4):6831-6842. https://doi.org/10.3390/ijms15046831
Chicago/Turabian StyleLiu, Xiaomin, Jie Li, Wei Guo, Ruijun Li, Dan Zhao, and Xinna Li. 2014. "A New Type I Peritrophic Membrane Protein from Larval Holotrichia oblita (Coleoptera: Melolonthidae) Binds to Chitin" International Journal of Molecular Sciences 15, no. 4: 6831-6842. https://doi.org/10.3390/ijms15046831