Genome-Wide Identification, 3D Modeling, Expression and Enzymatic Activity Analysis of Cell Wall Invertase Gene Family from Cassava (Manihot esculenta Crantz)

 , ,
, ,
Abstract
:
1. Introduction
2. Results
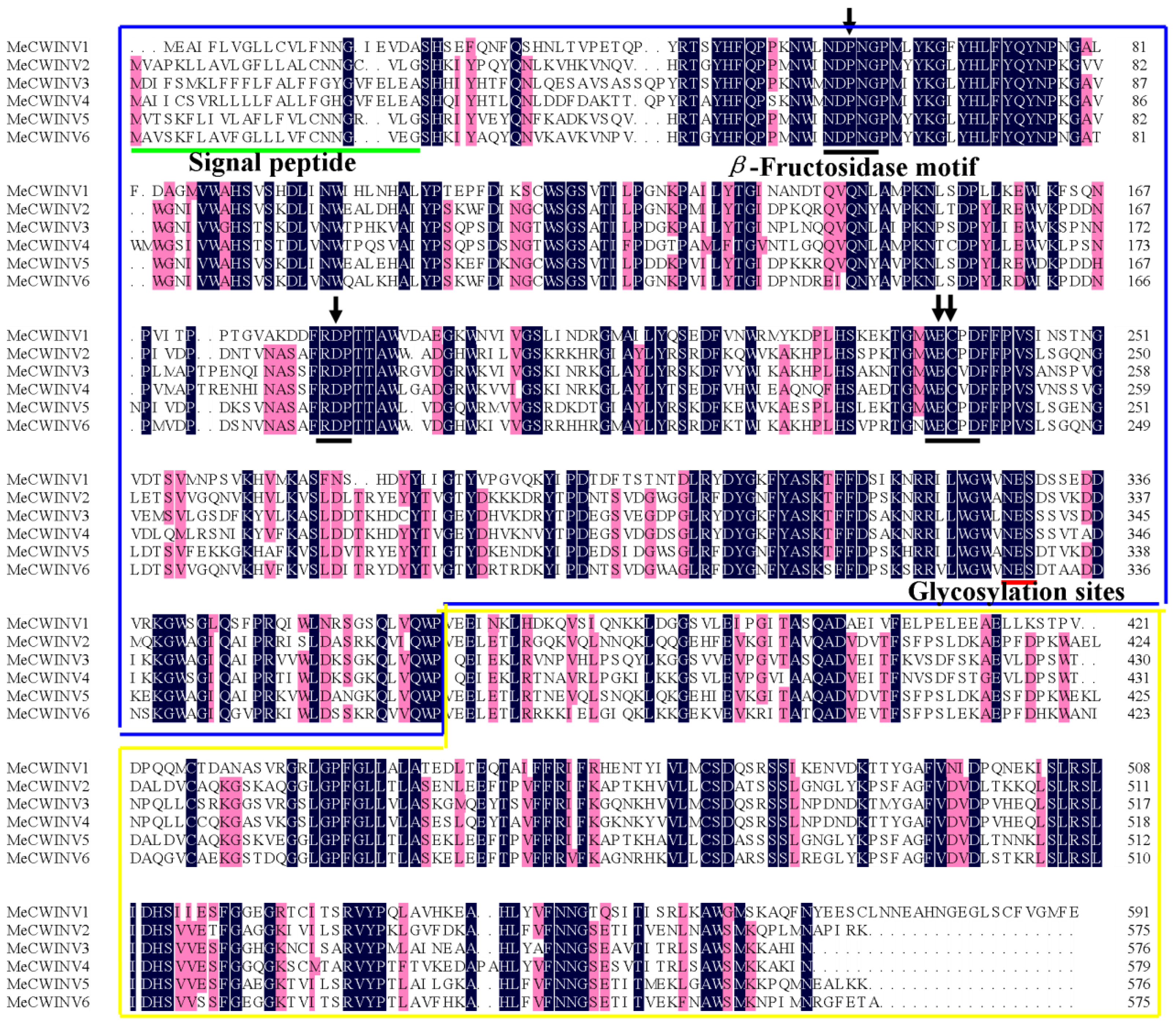
2.1. Identification and Characterization of the MeCWINV Genes in Cassava
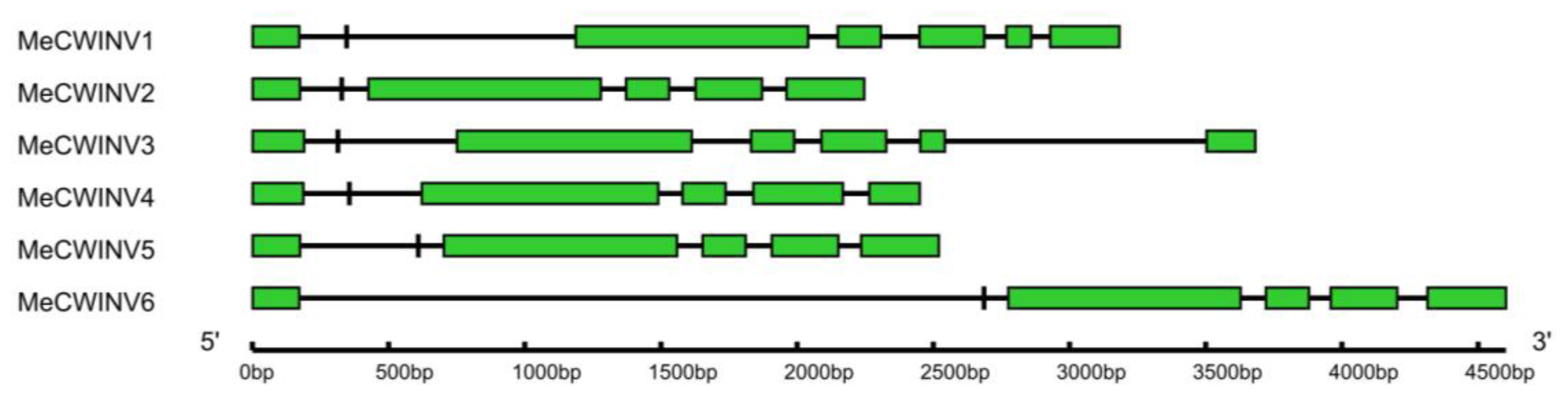
2.2. Gene Structure Analysis of the MeCWINV Genes
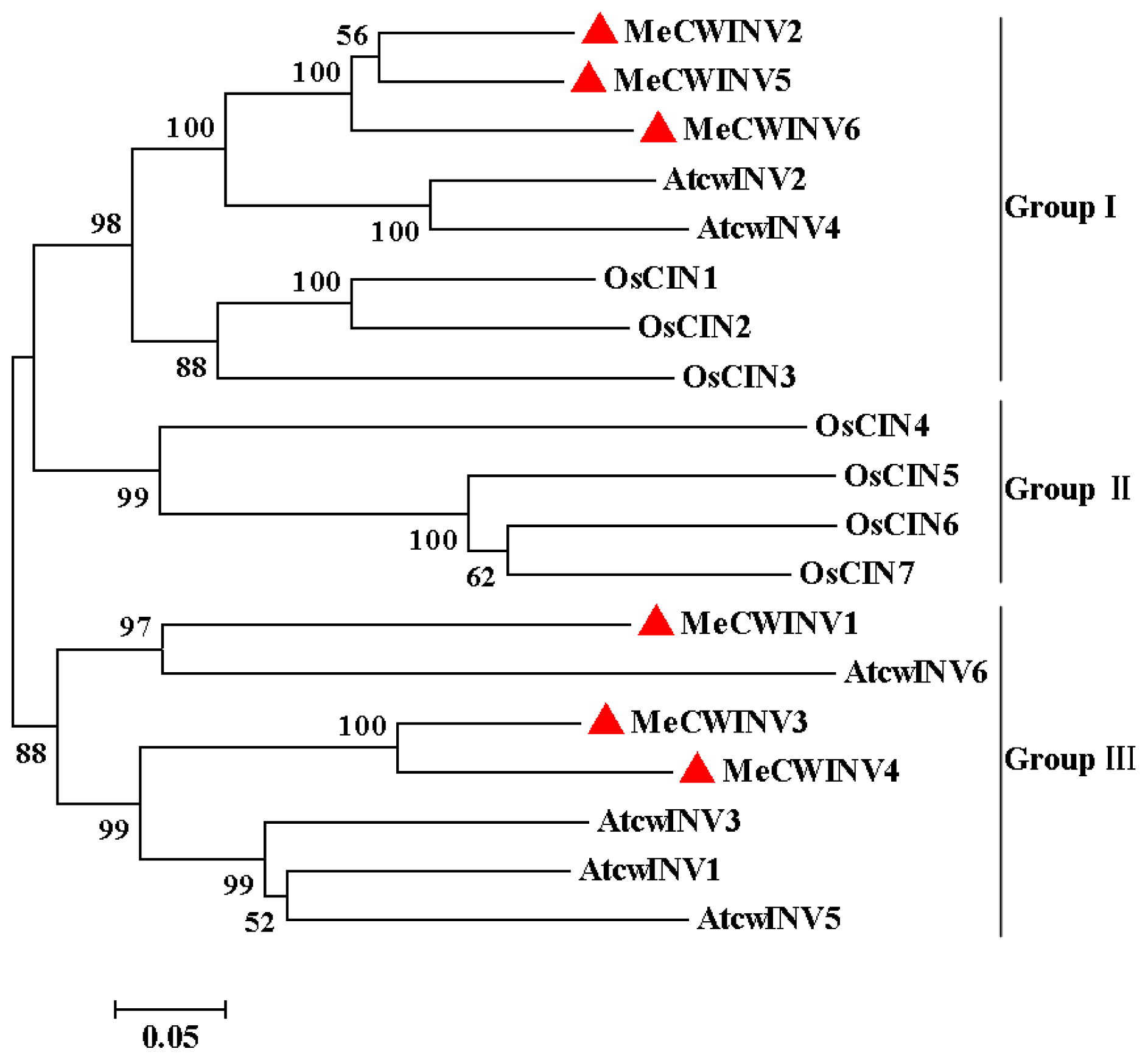
2.3. Phylogenetic Analysis of the MeCWINV Genes
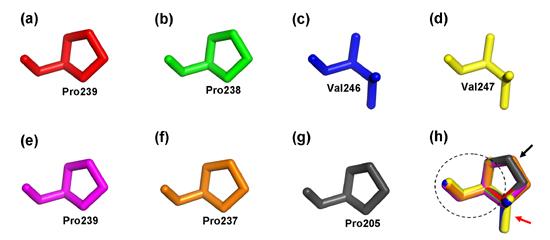
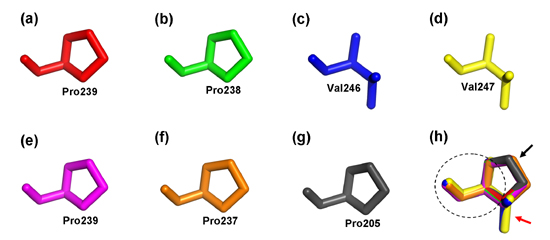
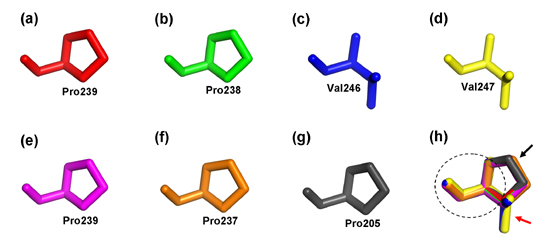
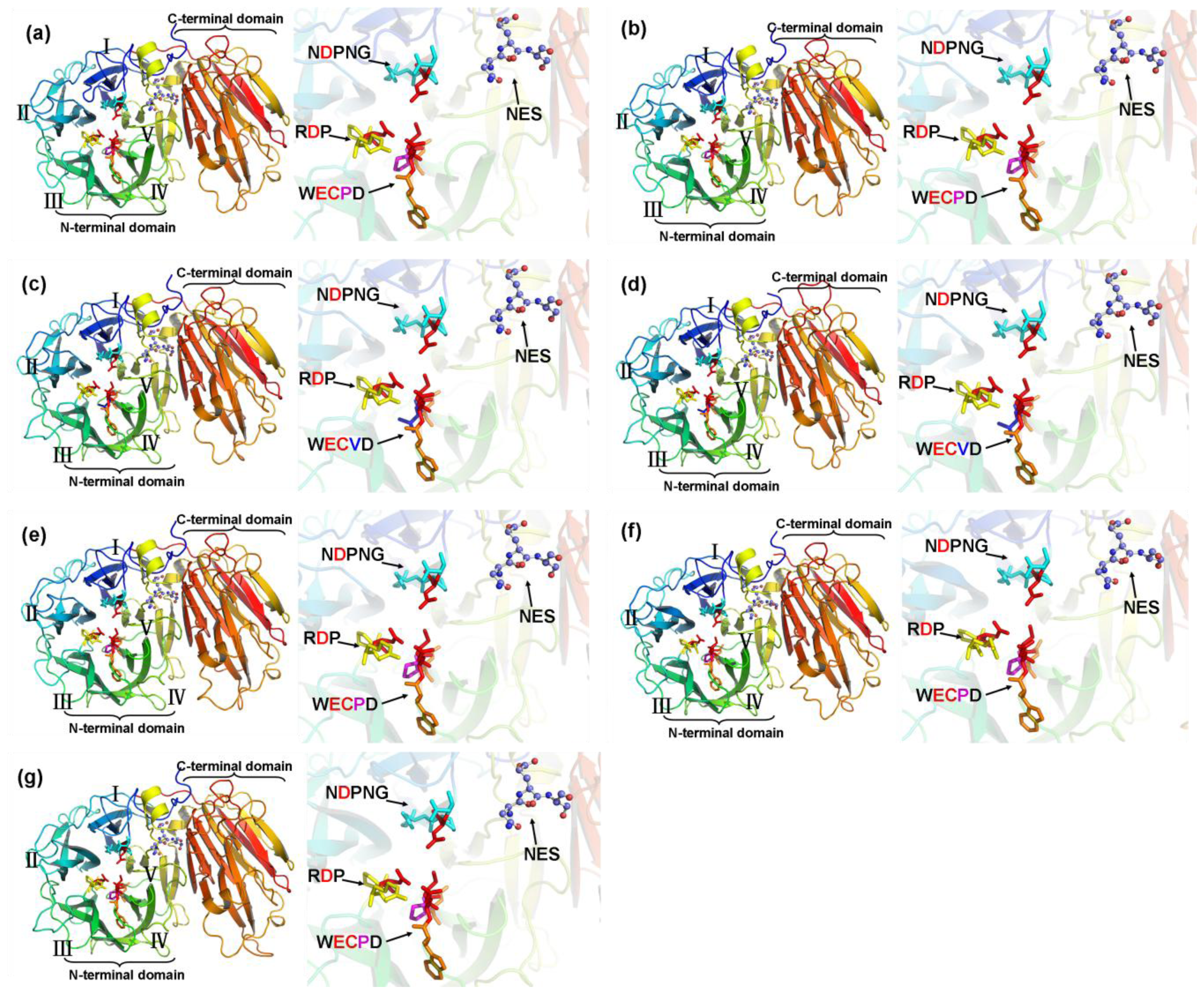
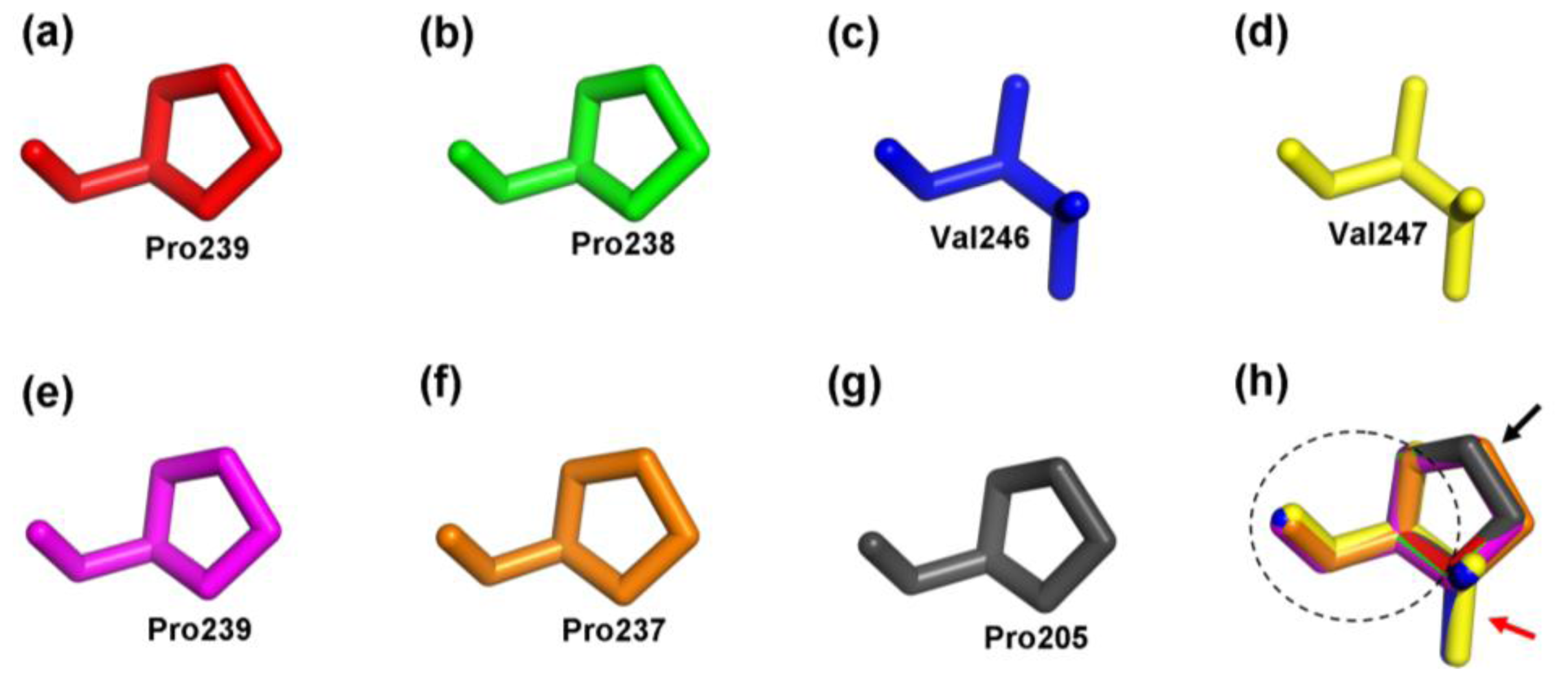
2.4. Structure Prediction and Homology Modeling of the MeCWINV Genes
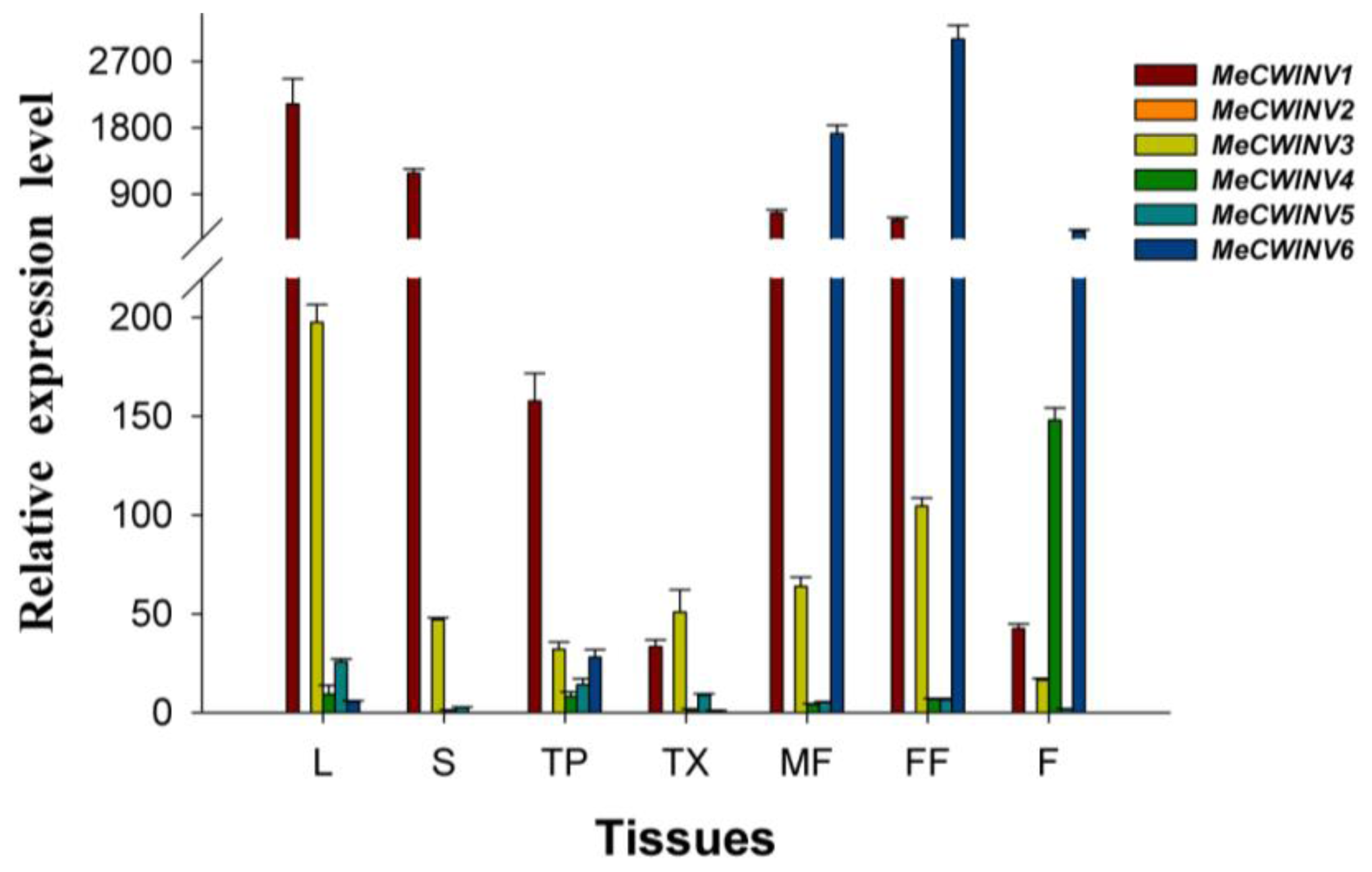
2.5. The Differential Expression Analysis of the MeCWINV Genes in Cassava Organs or Tissues
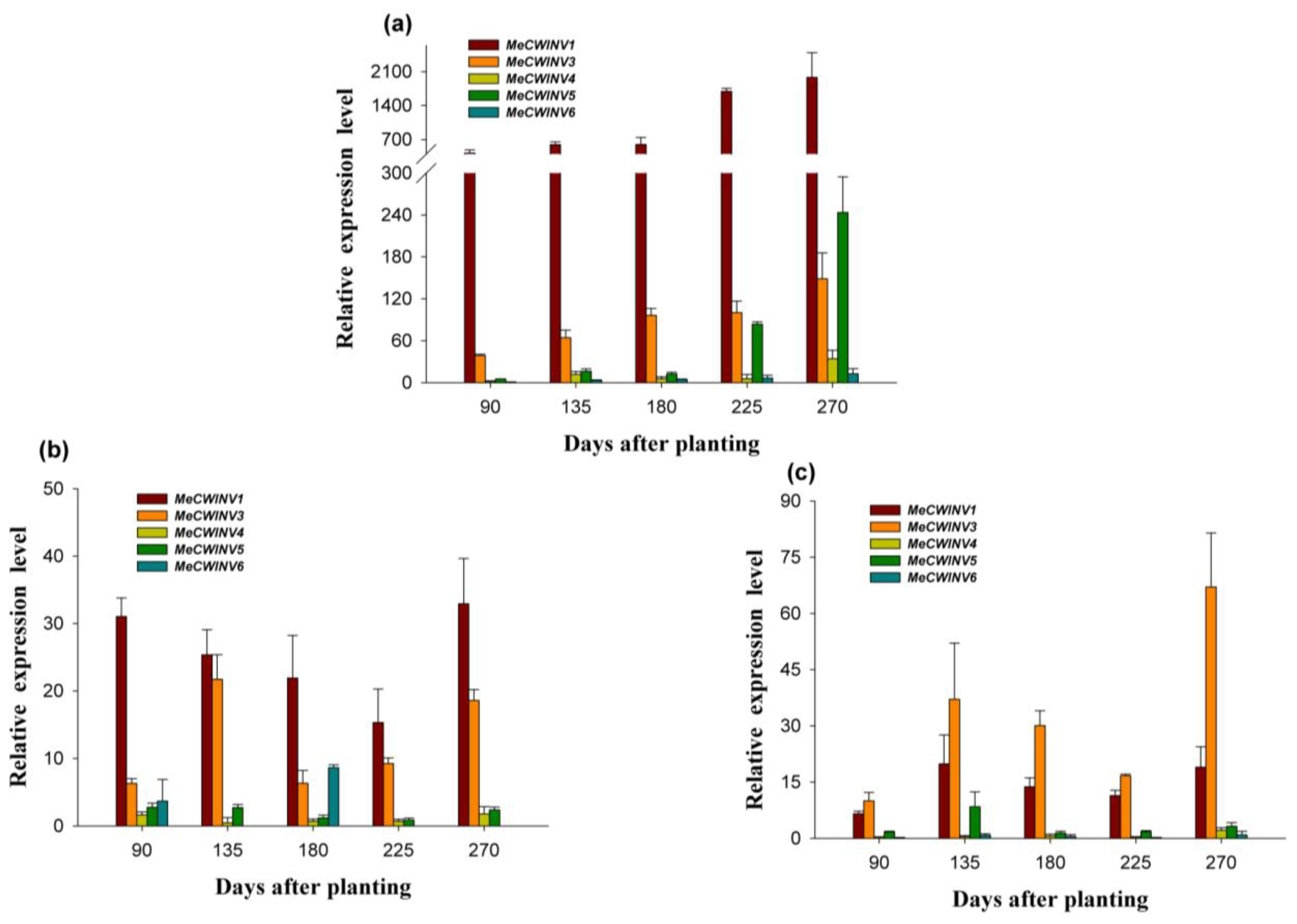
2.6. The Differential Expression of the MeCWINV Genes during Cassava Tuber Root Development
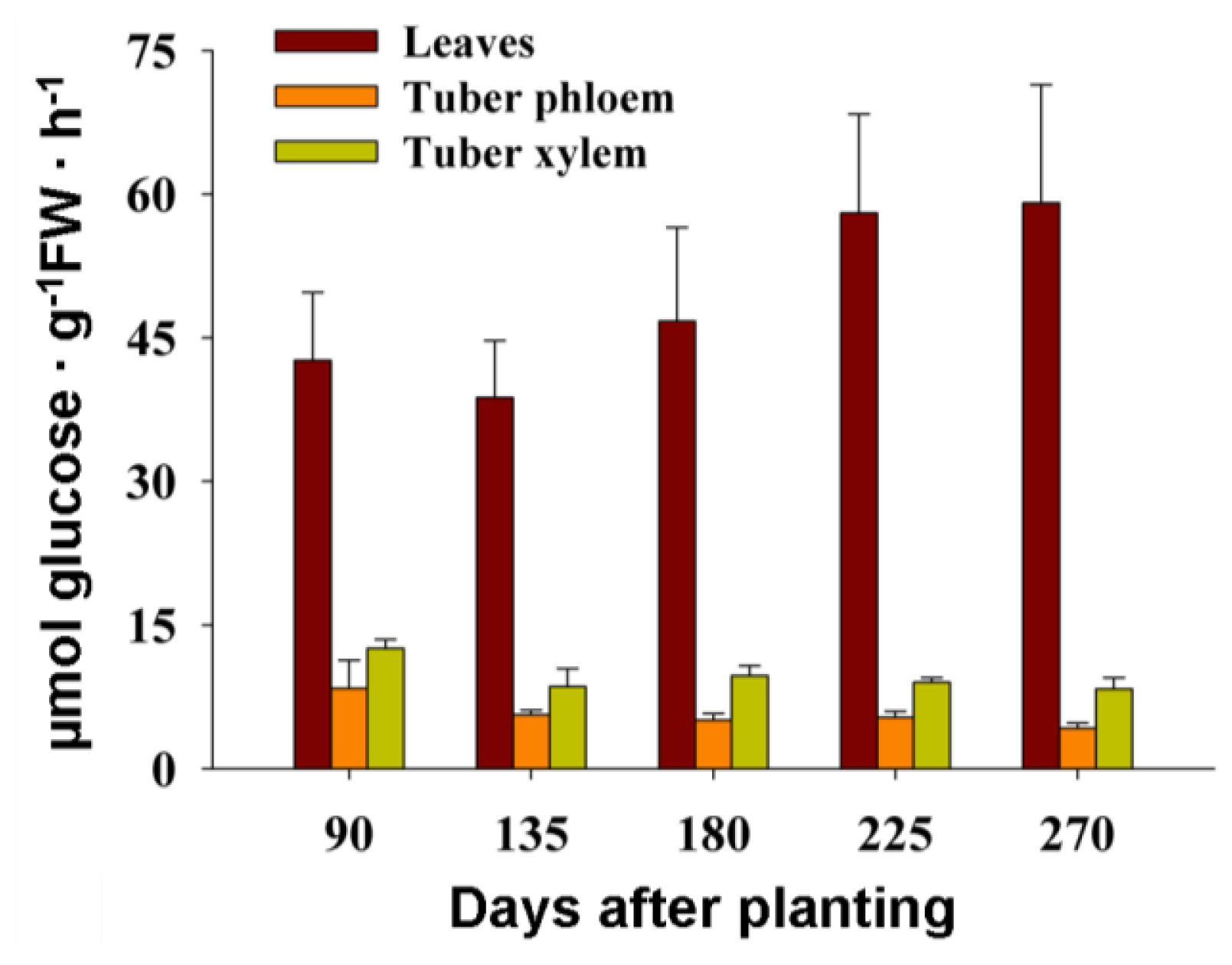
2.7. The Activity of Cell Wall Invertase in Cassava Leaves and Tubers during Tuber Root Development
3. Discussion
3.1. Identification and Characterization of the MeCWINV Genes
3.2. Protein Structural Features of the MeCWINVs in Cassava
3.3. Differential Expression and Enzymatic Activity Analysis of the MeCWINV Genes
4. Experimental Section
4.1. Plant Materials
4.2. Cloning of the Full-Length MeCWINV Genes
4.3. Sequence Alignment and Phylogenetic Analysis
4.4. Signal Peptide and Subcellular Localization Prediction
4.5. Exon-Intron Structure Analysis
4.6. Homology Modeling and Structure Prediction
4.7. Real-Time RT-PCR Analysis
4.8. Activity Analysis of the Cell Wall Invertase
5. Conclusions
Supplementary Information
ijms-15-07313-s001.pdfAcknowledgments
Conflicts of Interest
Abbreviations
| a.a | amino acids |
| bp | base pair |
| BLAST | basic local alignment search tool |
| ORF | open reading frame |
| RACE | rapid amplification of cDNA ends |
| RT-PCR | reverse transcription polymerase chain reaction |
| qRT-PCR | real-time quantitative PCR |
- Author ContributionsYuan Yao and Mengting Geng were responsible for all aspects of the research, including experimental design, data acquisition and analysis, and manuscript preparation. Xiaohui Wu worked on the preparation of the studied materials and gene cloning. Jiao Liu and Ruimei Li worked on primer design, technical and informatics’ analyses of these genes. Xinwen Hu and Jianchun Guo were responsibility for the programs and all experiments, and they critically revised the manuscript and provided the final approval of the article.
References
- Ihemere, U.; Arias-Garzon, D.; Lawrence, S.; Sayre, R. Genetic modification of cassava for enhanced starch production. Plant Biotechnol. J 2006, 4, 453–465. [Google Scholar]
- Zhang, Y.; Zhang, A.; Jiang, J. Gene expression patterns of invertase gene families and modulation of the inhibitor gene in tomato sucrose metabolism. Genet. Mol. Res 2013, 12, 3412–3420. [Google Scholar]
- Chandra, A.; Jain, R.; Solomon, S. Complexities of invertases controlling sucrose accumulation and retention in sugarcane. Curr. Sci 2012, 102, 857–866. [Google Scholar]
- Roitsch, T.; González, M.C. Function and regulation of plant invertases: Sweet sensations. Trends Plant Sci 2004, 9, 606–613. [Google Scholar]
- Lammens, W.; Le Roy, K.; van Laere, A.; Rabijns, A.; van den Ende, W. Crystal structures of Arabidopsis thaliana cell-wall invertase mutants in complex with sucrose. J. Mol. Biol 2008, 377, 378–385. [Google Scholar]
- Cho, J.I.; Lee, S.K.; Ko, S.; Kim, H.K.; Jun, S.H.; Lee, Y.H.; Bhoo, S.H.; Lee, K.W.; An, G.; Hahn, T.R. Molecular cloning and expression analysis of the cell-wall invertase gene family in rice (Oryza sativa L.). Plant Cell Rep 2005, 24, 225–236. [Google Scholar]
- Chen, T.-H.; Huang, Y.-C.; Yang, C.-S.; Yang, C.-C.; Wang, A.-Y.; Sung, H.-Y. Insights into the catalytic properties of bamboo vacuolar invertase through mutational analysis of active site residues. Phytochemistry 2009, 70, 25–31. [Google Scholar]
- Goetz, M.; Roitsch, T. The different pH optima and substrate specificities of extracellular and vacuolar invertases from plants are determined by a single amino-acid substitution. Plant J 1999, 20, 707–711. [Google Scholar]
- Yu, X.; Wang, X.; Zhang, W.; Qian, T.; Tang, G.; Guo, Y.; Zheng, C. Antisense suppression of an acid invertase gene (MAI1) in muskmelon alters plant growth and fruit development. J. Exp. Bot 2008, 59, 2969–2977. [Google Scholar]
- Qi, X.; Wu, Z.; Li, J.; Mo, X.; Wu, S.; Chu, J.; Wu, P. AtCYT-INV1, a neutral invertase, is involved in osmotic stress-induced inhibition on lateral root growth in Arabidopsis. Plant Mol. Biol 2007, 64, 575–587. [Google Scholar]
- Kim, J.-Y.; Mahé, A.; Brangeon, J.; Prioul, J.-L. A maize vacuolar invertase, IVR2, is induced by water stress. Organ/tissue specificity and diurnal modulation of expression. Plant Physiol 2000, 124, 71–84. [Google Scholar]
- Tang, G.Q.; Luscher, M.; Sturm, A. Antisense repression of vacuolar and cell wall invertase in transgenic carrot alters early plant development and sucrose partitioning. Plant Cell 1999, 11, 177–189. [Google Scholar]
- Schaarschmidt, S.; Roitsch, T.; Hause, B. Arbuscular mycorrhiza induces gene expression of the apoplastic invertase LIN6 in tomato (Lycopersicon esculentum) roots. J. Exp. Bot 2006, 57, 4015–4023. [Google Scholar]
- Jain, M.; Chourey, P.S.; Boote, K.J.; Allen, L.H., Jr. Short-term high temperature growth conditions during vegetative-to-reproductive phase transition irreversibly compromise cell wall invertase-mediated sucrose catalysis and microspore meiosis in grain sorghum (Sorghum bicolor). J. Plant Physiol 2010, 167, 578–582. [Google Scholar]
- Kocal, N.; Sonnewald, U.; Sonnewald, S. Cell wall-bound invertase limits sucrose export and is involved in symptom development and inhibition of photosynthesis during compatible interaction between tomato and Xanthomonas campestris pv. vesicatoria. Plant Physiol 2008, 148, 1523–1536. [Google Scholar]
- Chourey, P.; Jain, M.; Li, Q.-B.; Carlson, S. Genetic control of cell wall invertases in developing endosperm of maize. Planta 2006, 223, 159–167. [Google Scholar]
- Phytozome. Available online: http://www.phytozome.net/cassava accessed on 15 April, 2011.
- Vargas, W.; Cumino, A.; Salerno, G.L. Cyanobacterial alkaline/neutral invertases. Origin of sucrose hydrolysis in the plant cytosol? Planta 2003, 216, 951–960. [Google Scholar]
- Bocock, P.; Morse, A.; Dervinis, C.; Davis, J. Evolution and diversity of invertase genes in Populus trichocarpa. Planta 2008, 227, 565–576. [Google Scholar]
- Fridman, E.; Zamir, D. Functional divergence of a syntenic invertase gene family in tomato, potato, and Arabidopsis. Plant Physiol 2003, 131, 603–609. [Google Scholar]
- Ji, X.; van den Ende, W.; van Laere, A.; Cheng, S.; Bennett, J. Structure, evolution, and expression of the two invertase gene families of rice. J. Mol. Evol 2005, 60, 615–634. [Google Scholar]
- Thelander, M.; Nilsson, A.; Ronne, H. Annual Plant Reviews; Wiley: New York, NY, USA, 2009; Volume 36, pp. 211–245. [Google Scholar]
- Alberto, F.; Bignon, C.; Sulzenbacher, G.; Henrissat, B.; Czjzek, M. The three-dimensional structure of invertase (β-fructosidase) from Thermotoga maritima reveals a bimodular arrangement and an evolutionary relationship between retaining and inverting glycosidases. J. Biol. Chem 2004, 279, 18903–18910. [Google Scholar]
- Verhaest, M.; Lammens, W.; Le Roy, K.; de Coninck, B.; de Ranter, C.J.; van Laere, A.; van den Ende, W.; Rabijns, A. X-ray diffraction structure of a cell-wall invertase from Arabidopsis thaliana. Acta Cryst. D 2006, 62, 1555–1563. [Google Scholar]
- Li, Z.; Palmer, W.M.; Martin, A.P.; Wang, R.; Rainsford, F.; Jin, Y.; Patrick, J.W.; Yang, Y.; Ruan, Y.L. High invertase activity in tomato reproductive organs correlates with enhanced sucrose import into, and heat tolerance of, young fruit. J. Exp. Bot 2012, 63, 1155–1166. [Google Scholar]
- Schneidereit, A.; Scholz-Starke, J.; Büttner, M. Functional characterization and expression analyses of the glucose-specific ATSTP9 monosaccharide transporter in pollen of Arabidopsis. Plant Physiol 2003, 133, 182–190. [Google Scholar]
- Schneidereit, A.; Scholz-Starke, J.; Sauer, N.; Büttner, M. AtSTP11, a pollen tube-specific monosaccharide transporter in Arabidopsis. Planta 2005, 221, 48–55. [Google Scholar]
- Chourey, P.S.; Li, Q.-B.; Cevallos-Cevallos, J. Pleiotropy and its dissection through a metabolic gene Miniature1 (Mn1) that encodes a cell wall invertase in developing seeds of maize. Plant Sci 2012, 184, 45–53. [Google Scholar]
- Sturm, A.; Tang, G.Q. The sucrose-cleaving enzymes of plants are crucial for development, growth and carbon partitioning. Trends Plant Sci 1999, 4, 401–407. [Google Scholar]
- Kühn, C.; Hajirezaei, M.R.; Fernie, A.R.; Roessner-Tunali, U.; Czechowski, T.; Hirner, B.; Frommer, W.B. The sucrose transporter StSUT1 localizes to sieve elements in potato tuber phloem and influences tuber physiology and development. Plant Physiol 2003, 131, 102–113. [Google Scholar]
- Viola, R.; Roberts, A.G.; Haupt, S.; Gazzani, S.; Hancock, R.D.; Marmiroli, N.; Machray, G.C.; Oparka, K.J. Tuberization in potato involves a switch from apoplastic to symplastic phloem unloading. Plant Cell 2001, 13, 385–398. [Google Scholar]
- Hothorn, M.; van den Ende, W.; Lammens, W.; Rybin, V.; Scheffzek, K. Structural insights into the pH-controlled targeting of plant cell-wall invertase by a specific inhibitor protein. Proc. Natl. Acad. Sci. USA 2010, 107, 17427–17432. [Google Scholar]
- Bonfig, K.B.; Gabler, A.; Simon, U.K.; Luschin-Ebengreuth, N.; Hatz, M.; Berger, S.; Muhammad, N.; Zeier, J.; Sinha, A.K.; Roitsch, T. Post-translational derepression of invertase activity in source leaves via down-regulation of invertase inhibitor expression is part of the plant defense response. Mol. Plant 2010, 3, 1037–1048. [Google Scholar]
- SignalP. Available online: http://www.cbs.dtu.dk/services/SignalP/ accessed on 15 April, 2013.
- TargetP. Available online: http://www.cbs.dtu.dk/services/TargetP/ accessed on 17 April 2013.
- Gene Structure Display Server. Available online: http://gsds.cbi.pku.edu.cn/index.php accessed on 25 April 2013.
- SWISS-MODEL. Available online: http://swissmodel.expasy.org accessed on 20 January 2014.
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar]
- Albertson, P.L.; Peters, K.F.; Grof, C.P.L. An improved method for the measurement of cell wall invertase activity in sugarcane tissue. Funct. Plant Biol 2001, 28, 323–328. [Google Scholar]
- Mišić, D.; Dragićević, M.; Šiler, B.; Nestorović Živković, J.; Maksimović, V.; Momčilović, I.; Nikolic, M. Sugars and acid invertase mediate the physiological response of Schenkia spicata root cultures to salt stress. J. Plant Physiol 2012, 169, 1281–1289. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession numbers | Genomic location | ORF length | Length (a.a) |
|---|---|---|---|---|
| MeCWINV1 | JQ339929 | scaffold00206: 207290–10385 | 1779 | 592 |
| MeCWINV2 | JX291160 | scaffold06550: 791589–793834 | 1728 | 575 |
| MeCWINV3 | JN801147 | scaffold12525: 531768–539616 | 1731 | 576 |
| MeCWINV4 | JQ792172 | scaffold05875: 522938–525387 | 1731 | 576 |
| MeCWINV5 | JX291159 | scaffold06550: 798389–800908 | 1731 | 576 |
| MeCWINV6 | JQ339930 | scaffold02892: 67903–72504 | 1728 | 575 |
| Gene | Forward primer (5′ to 3′) | Reverse primer (5′ to 3′) |
|---|---|---|
| MeCWINV1 | CGCGGATCCAAAATAGAGATGGAAGCAAT | CGCGTCGACATCATTTCTCAAACATACCC |
| MeCWINV2 | CATGCCATGGTCTTCTTCATCTTCAGTC | CGGGTTACCTTTTATCTTTAGCTCAGC |
| MeCWINV3 | TCAATCAAAGGAGCTATGGACA | GCACTGGCTTCTTTTATTTCATC |
| MeCWINV4 | TATGGATCCCTCACCAGCATGGCTATCA | CATGTCGACTGTCAACCCTGGCTATTTCTCA |
| MeCWINV5 | TCATCCCGCAAATTCAACATT | GCTTACTTCTTTTCCACCTTATTTTTT |
| MeCWINV6 | CTCATTATCCCAAACAGATCAACC | ATTCTCACTTCTTTAAGCAGTCTCA |
| Gene | Forward primer (5′ to 3′) | Reverse primer (5′ to 3′) |
|---|---|---|
| MeCWINV1 | CAATGGAACTCAGAGCATAACC | TCTCAAACATACCCACAAAACA |
| MeCWINV2 | ATAGGAGGATTTTGTGGGGTTG | TGACTTGTTTTCTACTGGCATCT |
| MeCWINV3 | GCTTCGTGTGAATCCAGTCC | TCTGCCTGTGATGCTGTGA |
| MeCWINV4 | ATTTACTGTGAAGGAAGATGCG | TCTGTAGTTGTCAACCCTGGCT |
| MeCWINV5 | TTGGTAAGGCTCACTTGTTTGTAT | CAAGTATGGACATGTTAGACAGAATG |
| MeCWINV6 | GGAAGATTGTGGTGGGGAGTAG | TGCCCAACAACTGAAGTATCC |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yao, Y.; Geng, M.-T.; Wu, X.-H.; Liu, J.; Li, R.-M.; Hu, X.-W.; Guo, J.-C. Genome-Wide Identification, 3D Modeling, Expression and Enzymatic Activity Analysis of Cell Wall Invertase Gene Family from Cassava (Manihot esculenta Crantz). Int. J. Mol. Sci. 2014, 15, 7313-7331. https://doi.org/10.3390/ijms15057313
Yao Y, Geng M-T, Wu X-H, Liu J, Li R-M, Hu X-W, Guo J-C. Genome-Wide Identification, 3D Modeling, Expression and Enzymatic Activity Analysis of Cell Wall Invertase Gene Family from Cassava (Manihot esculenta Crantz). International Journal of Molecular Sciences. 2014; 15(5):7313-7331. https://doi.org/10.3390/ijms15057313
Chicago/Turabian StyleYao, Yuan, Meng-Ting Geng, Xiao-Hui Wu, Jiao Liu, Rui-Mei Li, Xin-Wen Hu, and Jian-Chun Guo. 2014. "Genome-Wide Identification, 3D Modeling, Expression and Enzymatic Activity Analysis of Cell Wall Invertase Gene Family from Cassava (Manihot esculenta Crantz)" International Journal of Molecular Sciences 15, no. 5: 7313-7331. https://doi.org/10.3390/ijms15057313