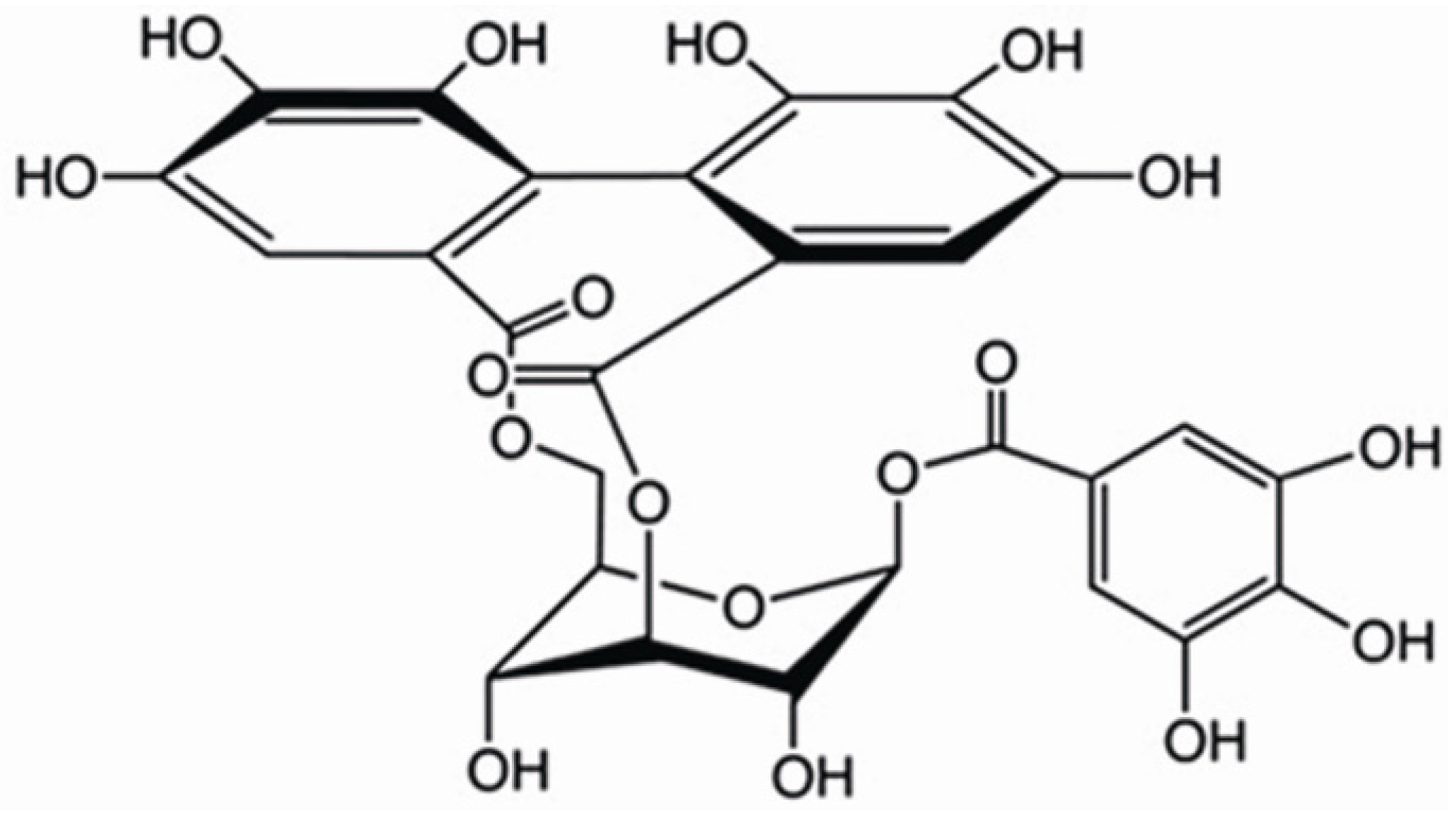
Corilagin Attenuates Aerosol Bleomycin-Induced Experimental Lung Injury

Abstract
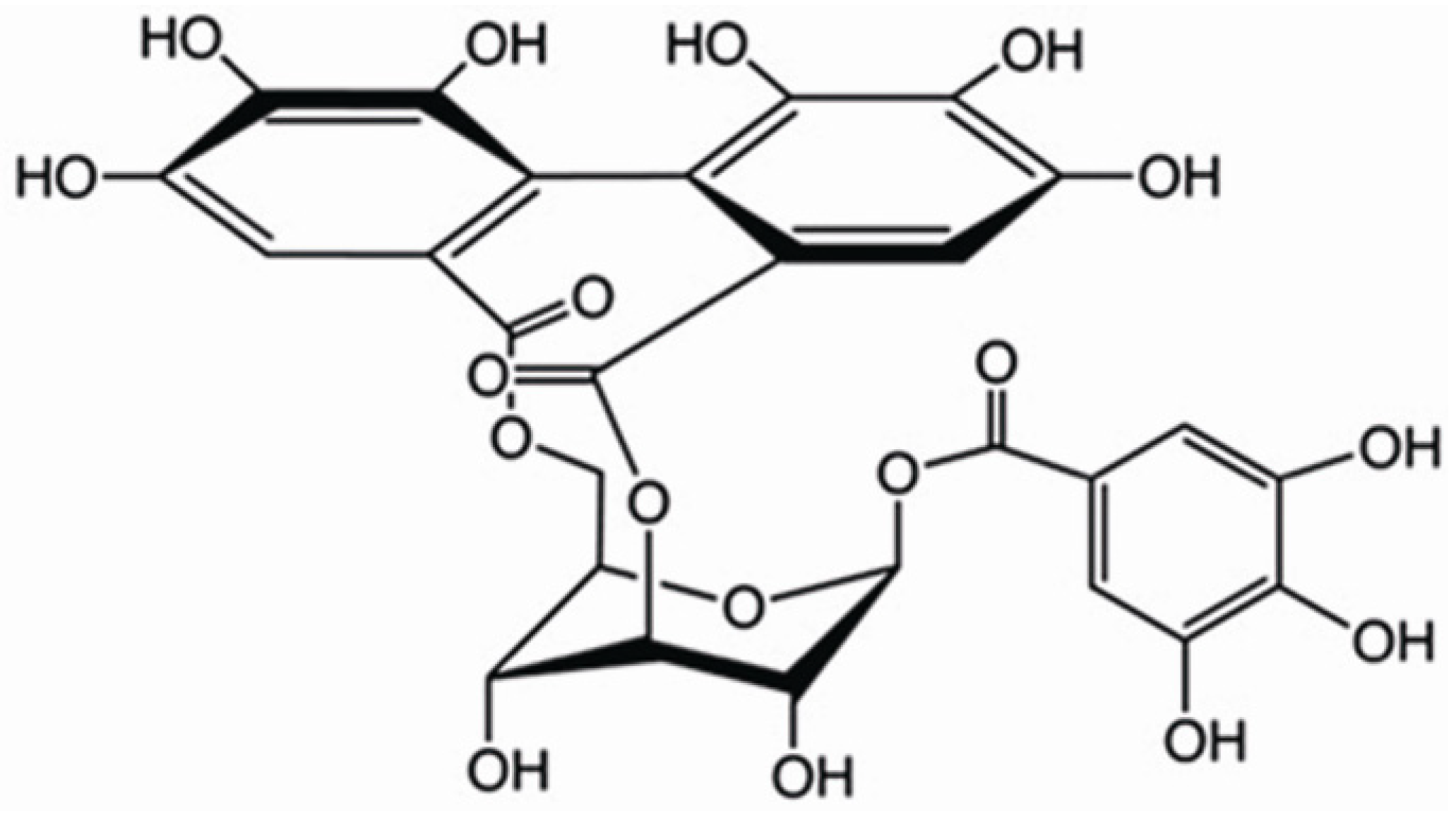
:
1. Introduction
2. Results and Discussion
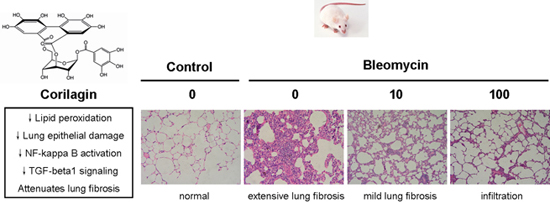
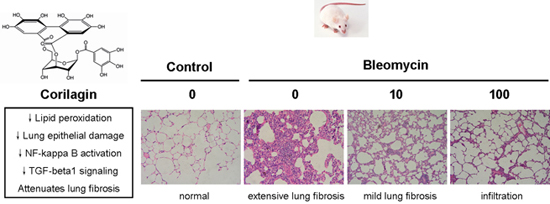
2.1. Macroscopic Observations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Group | F Value | p Value | |||
|---|---|---|---|---|---|---|
| ctrl | blm | l-cori | h-cori | |||
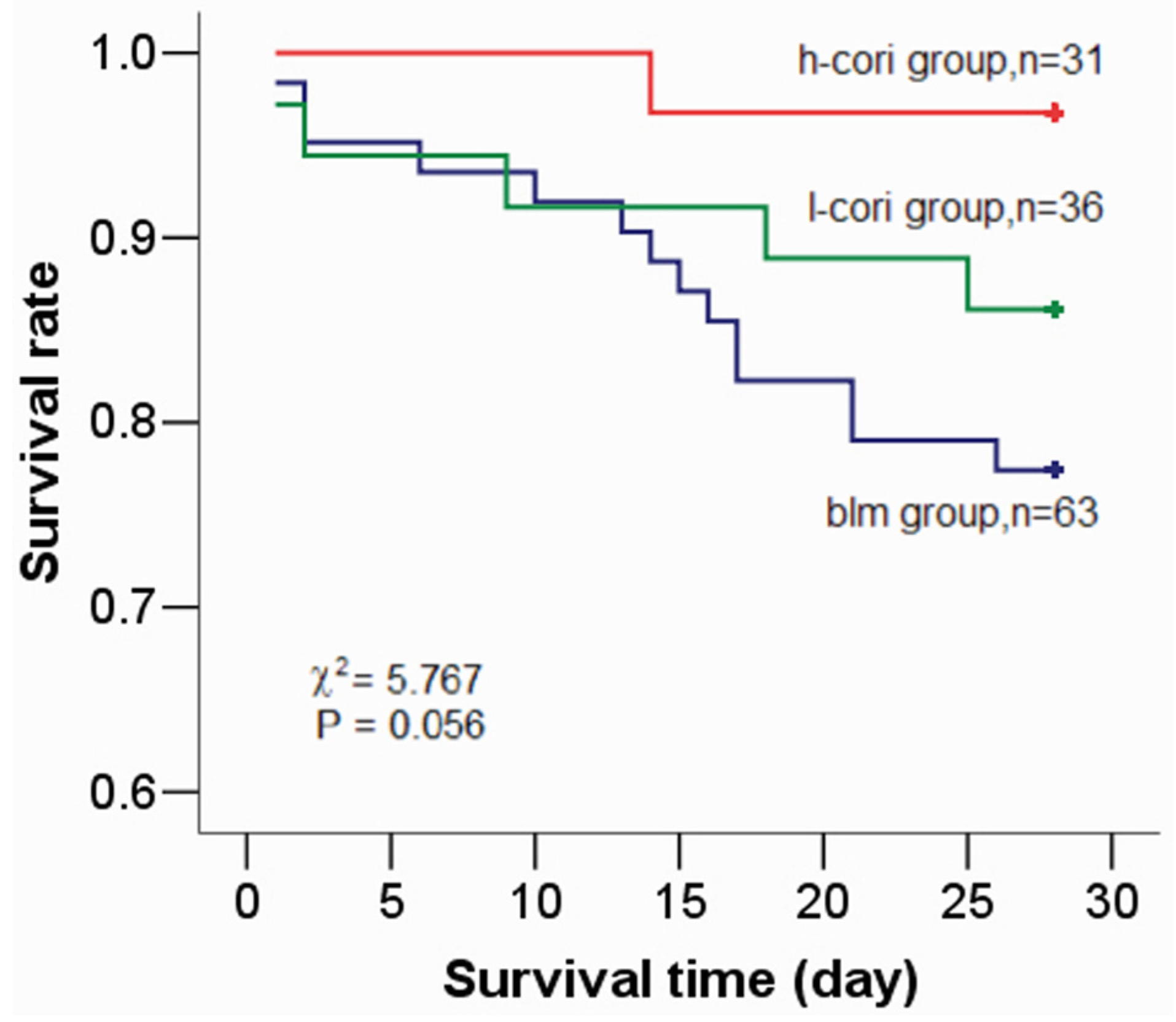
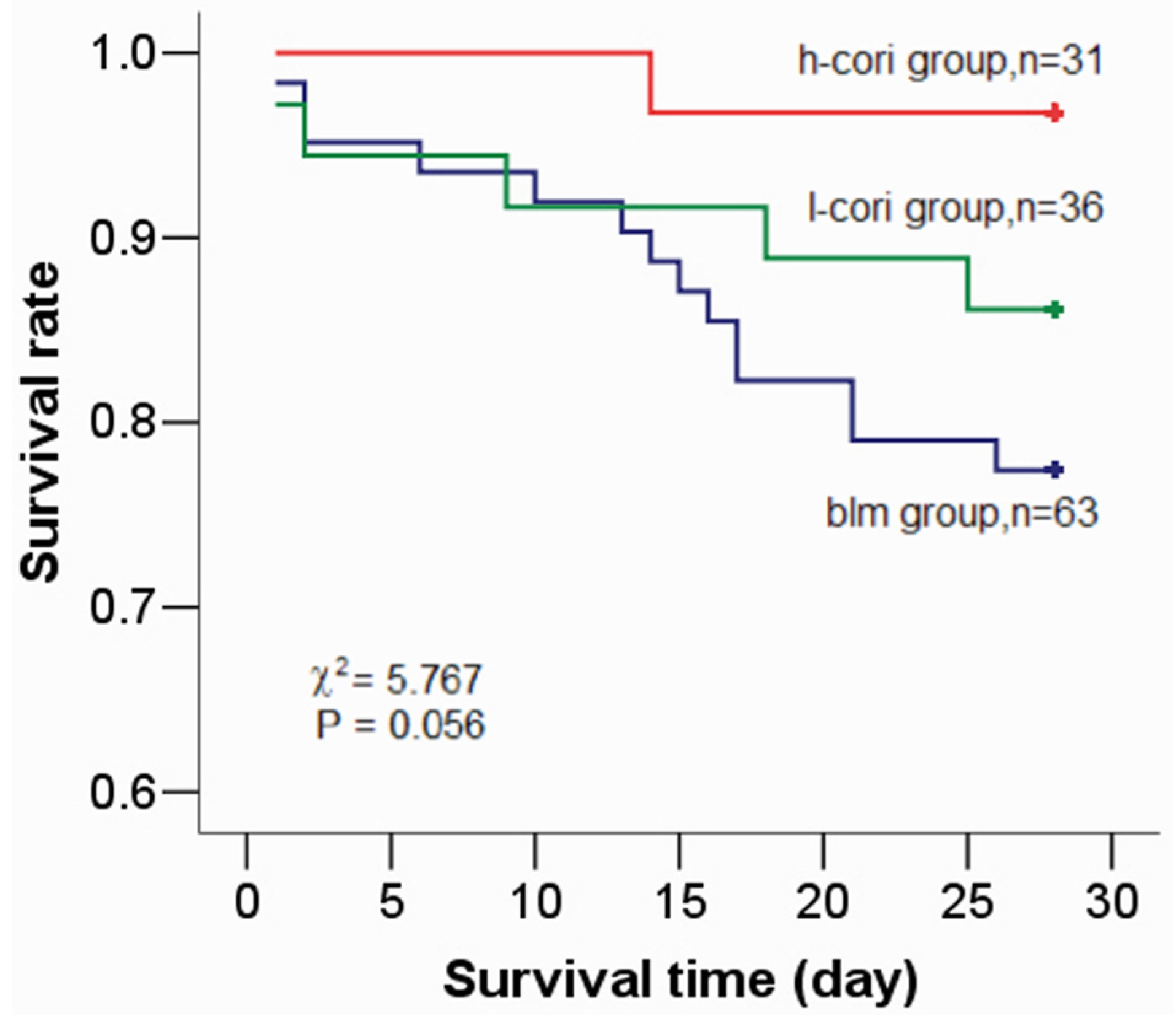
| Survival rate (%) | 100(14/14) | 77.4(48/62) | 86.1(31/36) | 96.8(30/31) | 9.07 † | 0.024 |
| Body weight (g) | ||||||
| d0 | 19.0 ± 0.7 | 19.2 ± 0.8 | 18.9 ± 0.7 | 19.0 ± 0.6 | 0.244 | 0.78 |
| d28 | 25.3 ± 2.6 | 18.6 ± 2.7 | 21.3 ± 3.2 *# | 24.4 ± 3.0 # | 1.764 | 0.033 |
| Ashcroft score | ||||||
| d7 | 0.4 ± 0.1 | 0.7 ± 0.2 | 1.0 ± 0.4 * | 0.9 ± 0.3 * | 6.533 | 0.002 |
| d15 | 0.8 ± 0.2 | 3.4 ± 0.7 * | 2.5 ± 0.6 * | 1.9 ± 0.7 *# | 24.145 | <0.001 |
| d28 | 1.4 ± 0.2 | 5.1 ± 1.0 * | 3.9 ± 0.8 * | 2.8 ± 0.8 *# | 30.076 | <0.001 |
| Hydroxyproline (mg/g) | ||||||
| d7 | 1.63 ± 0.11 | 1.85 ± 0.30 * | 1.83 ± 0.33 | 1.91 ± 0.24 * | 2.322 | 0.011 |
| d15 | 1.82 ± 0.27 | 3.76 ± 0.57 * | 2.88 ± 0.53 *# | 2.44 ± 0.41 *# | 18.675 | <0.001 |
| d28 | 2.20 ± 0.13 | 7.34 ± 0.92 *# | 5.10 ± 0.62 *# | 3.64 ± 0.58 *# | 85.053 | <0.001 |
| BALF cell count (104/mL) | ||||||
| d7 | 8.7 ± 1.6 | 22.3 ± 3.4 | 16.7 ± 1.2 | 13.4 ± 1.8 | 17.456 § | <0.001 |
| d15 | 9.0 ± 1.0 | 19.9 ± 2.0 * | 14.0 ± 2.0 *# | 12.7 ± 1.2 *# | 38.19 | <0.001 |
| d28 | 8.6 ± 1.4 | 17.9 ± 2.5 * | 12.4 ± 2.7 # | 13.0 ± 2.5 # | 13.244 | <0.001 |
| BALF differential cell proportion | ||||||
| Macrophages | ||||||
| d7 | 97.9 ± 0.5 | 57.7 ± 6.3 * | 63.2 ± 5.4 * | 69.5 ± 11.7 *# | 44.167 | <0.001 |
| d15 | 96.3 ± 0.4 | 56.1 ± 7.7 * | 57.8 ± 8.4 *# | 65.3 ± 7.7 *# | 35.945 | <0.001 |
| d28 | 96.1 ± 0.8 | 46.6 ± 8.2 *# | 73.7 ± 8.4 *# | 67.7 ± 7.5 *# | 42.586 | <0.001 |
| Neutrophils | ||||||
| d7 | 0.8 ± 0.2 | 10.0 ± 1.4 * | 14.2 ± 2.3 * | 15.2 ± 2.5 * | 64.645 | <0.001 |
| d15 | 2.4 ± 0.2 | 12.4 ± 2.3 * | 15.8 ± 3.5 * | 16.5 ± 5.2 * | 18.630 | <0.001 |
| d28 | 2.5 ± 0.1 | 7.7 ± 1.8 * | 11.3 ± 2.3 * | 21.8 ± 5.2 *# | 38.462 | <0.001 |
| Lymphocytes | ||||||
| d7 | 1.2 ± 0.2 | 33.4 ± 6.5 * | 22.3 ± 4.8 *# | 15.3 ± 3.6 *# | 47.488 | <0.001 |
| d15 | 1.1 ± 0.2 | 31.2 ± 5.2 | 24.7 ± 4.6 * | 17.3 ± 5.5 *# | 42.800 | <0.001 |
| d28 | 1.4 ± 0.3 | 24.9 ± 4.3 * | 15.4 ± 5.2 *# | 10.5 ± 4.3 *# | 29.966 | <0.001 |
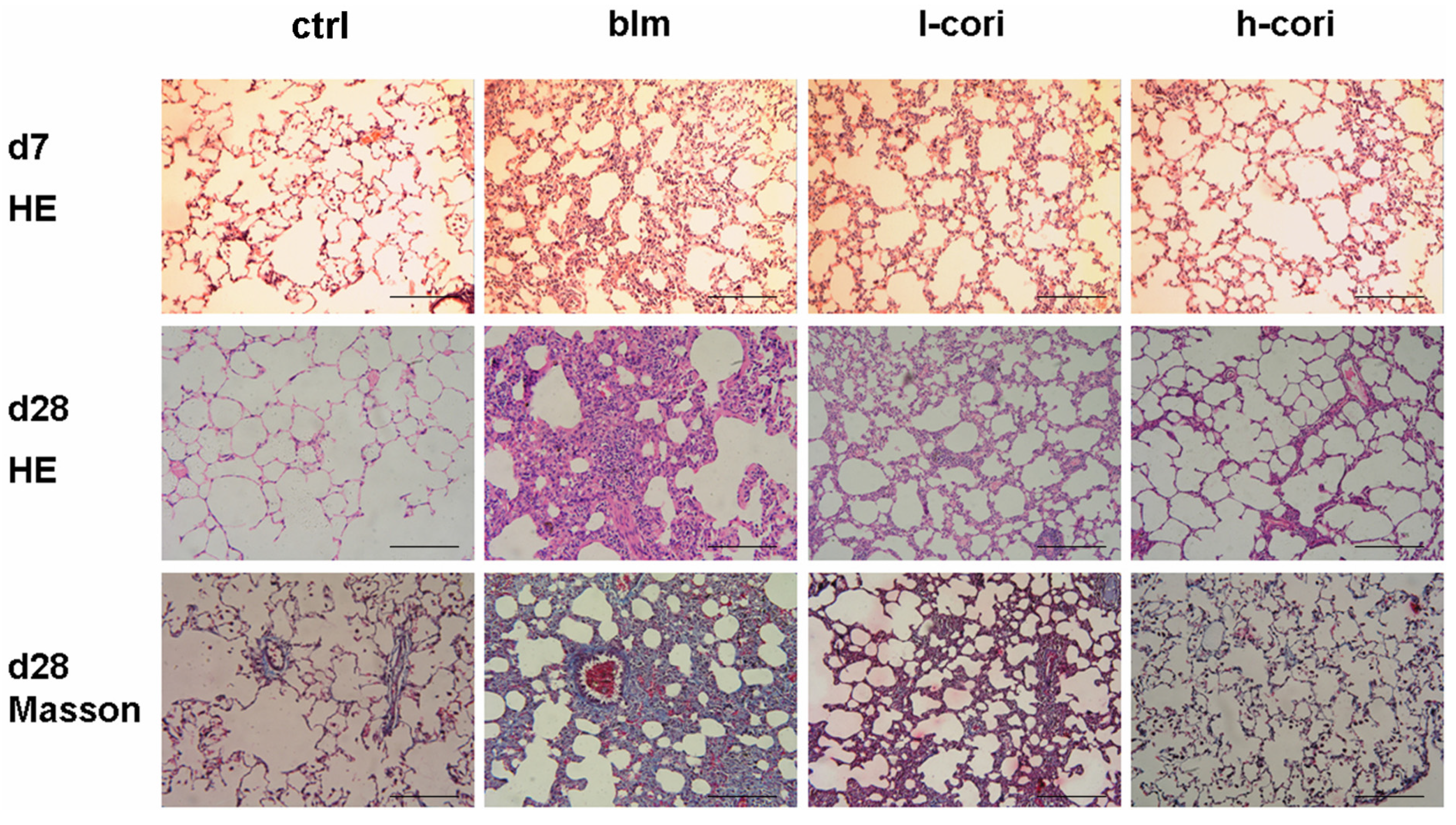
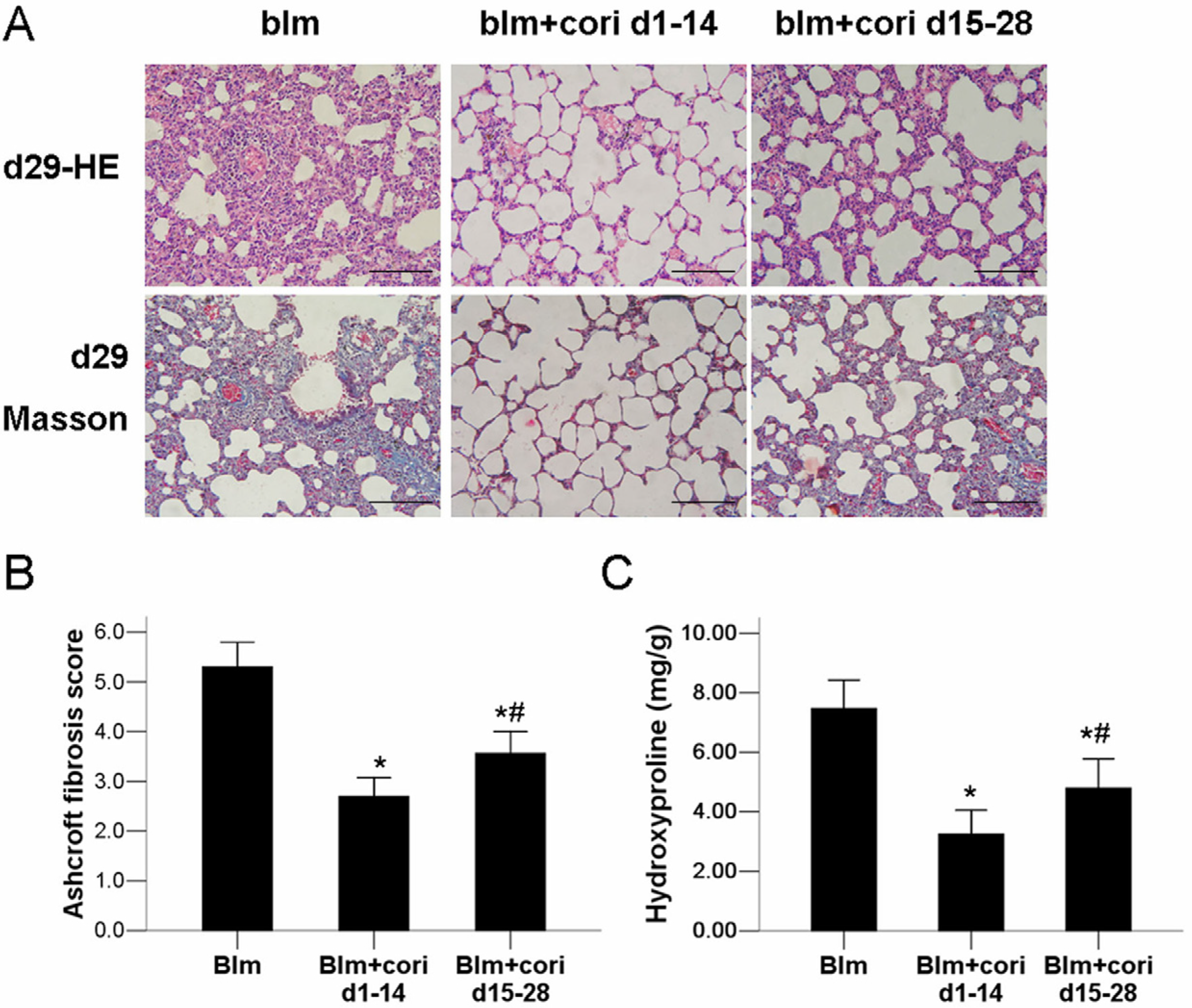
2.2. Corilagin Ameliorates Bleomycin-Induced Pulmonary Fibrosis
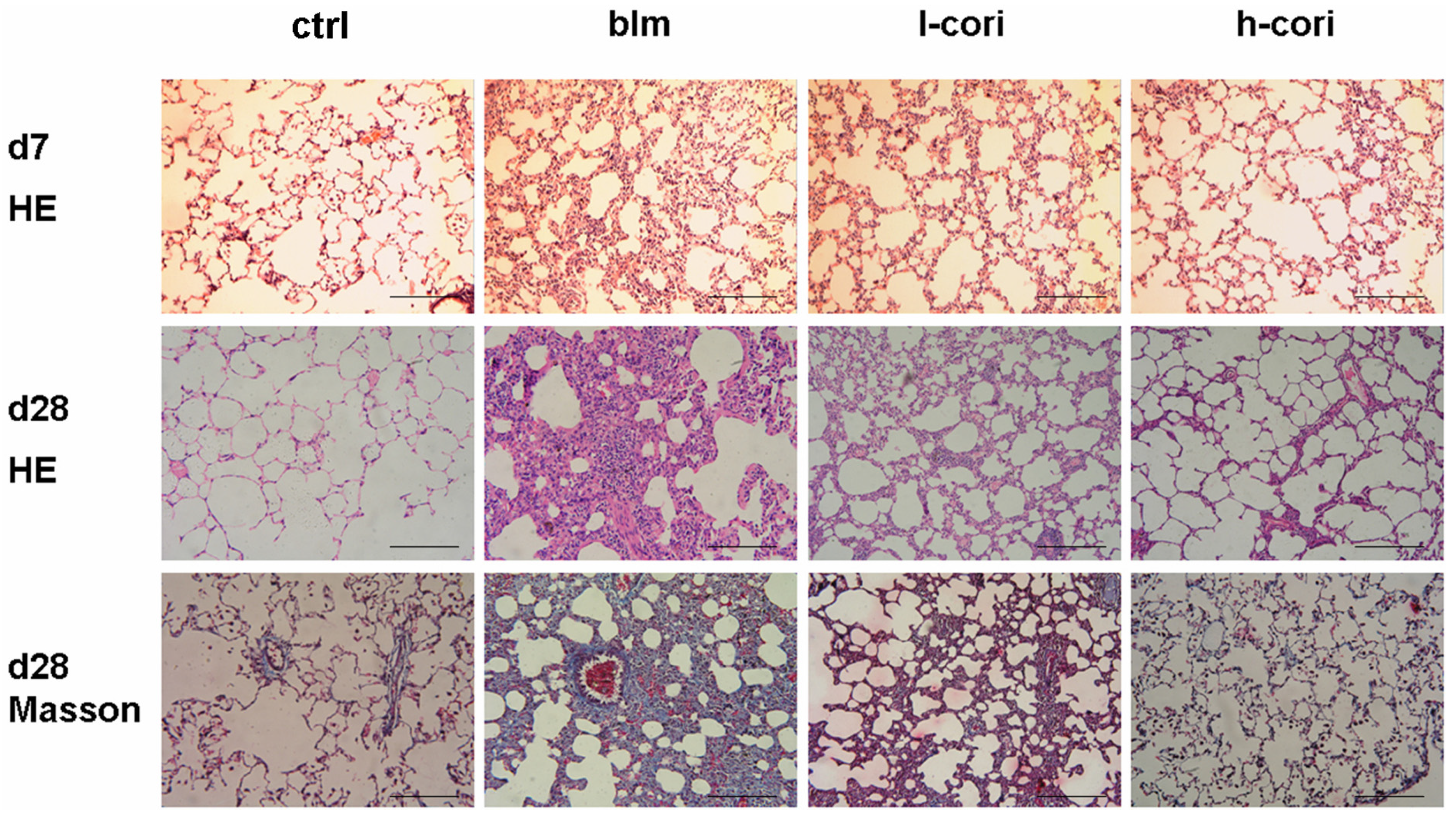
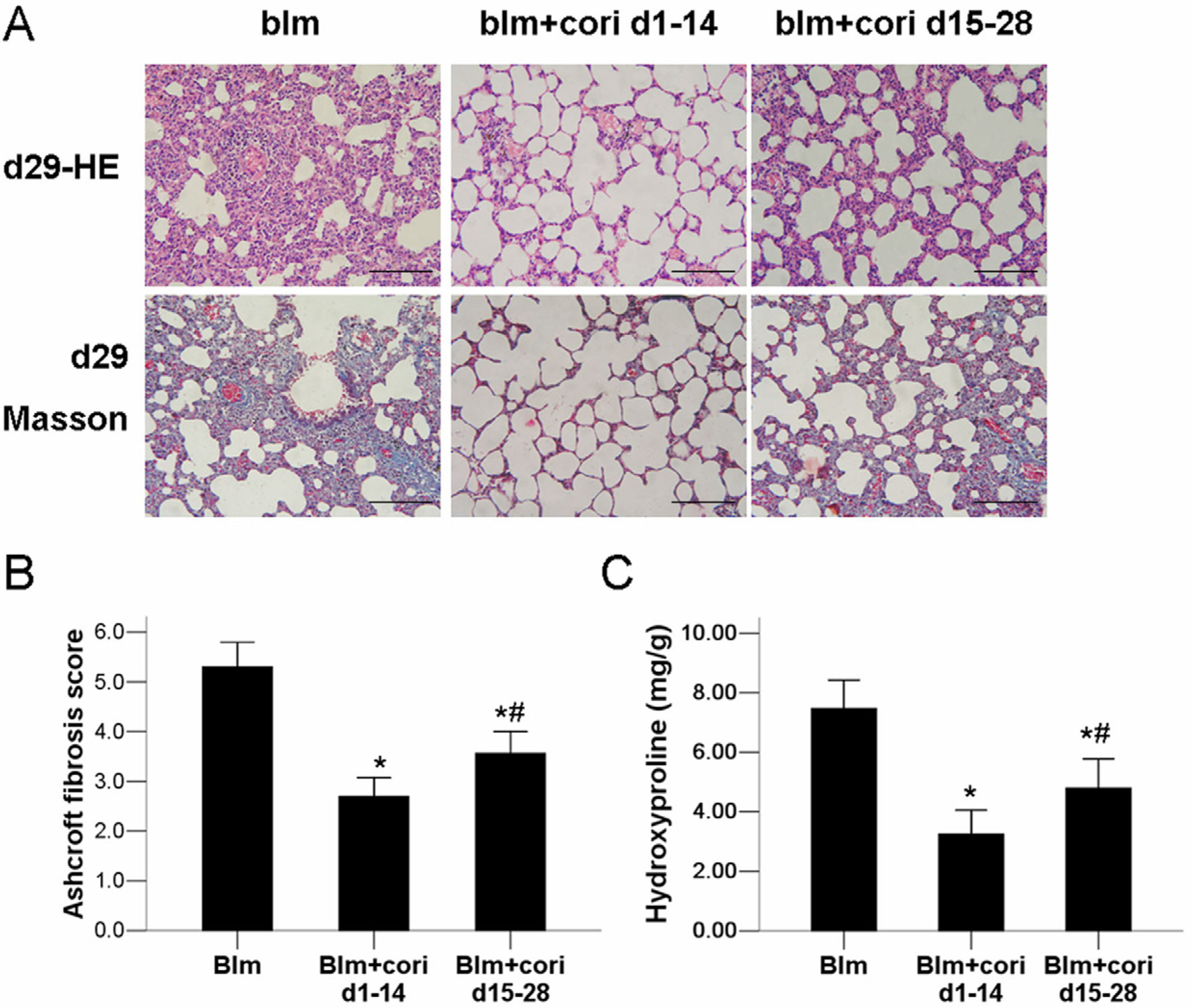
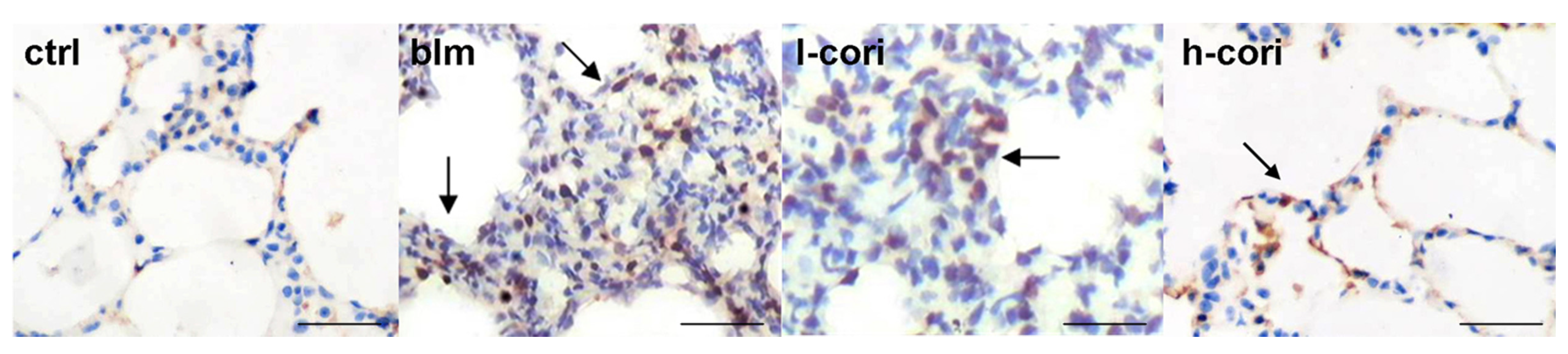
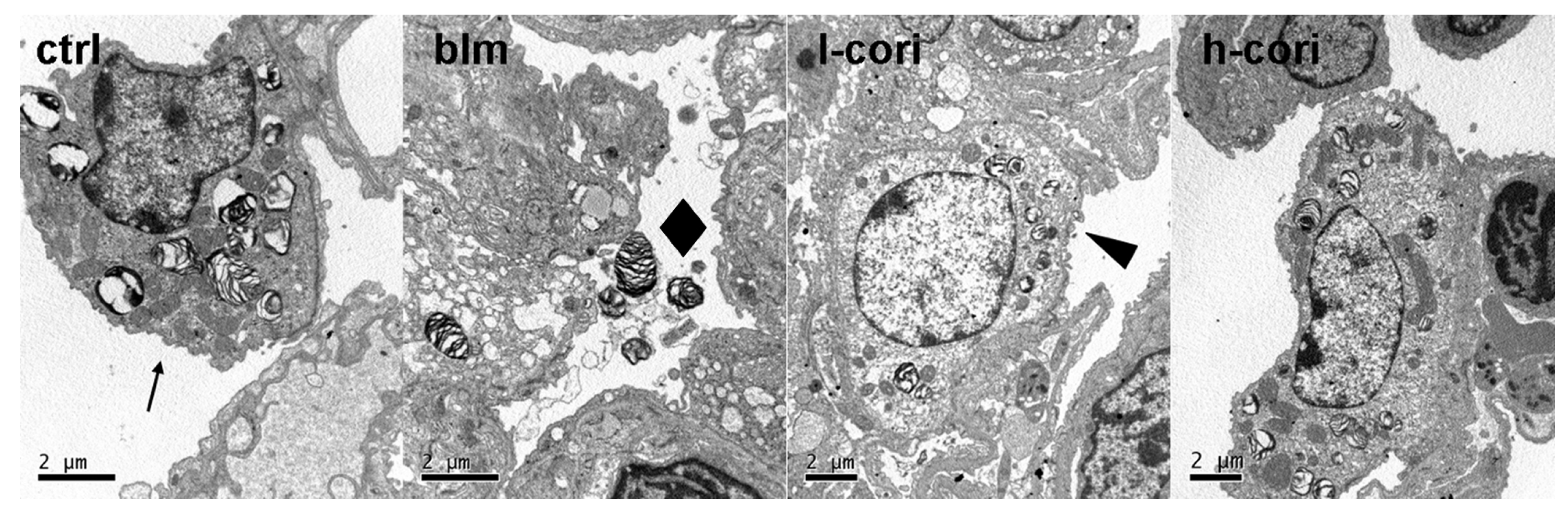
2.3. Corilagin Alleviates Bleomycin-Induced Lung Epithelial Injury
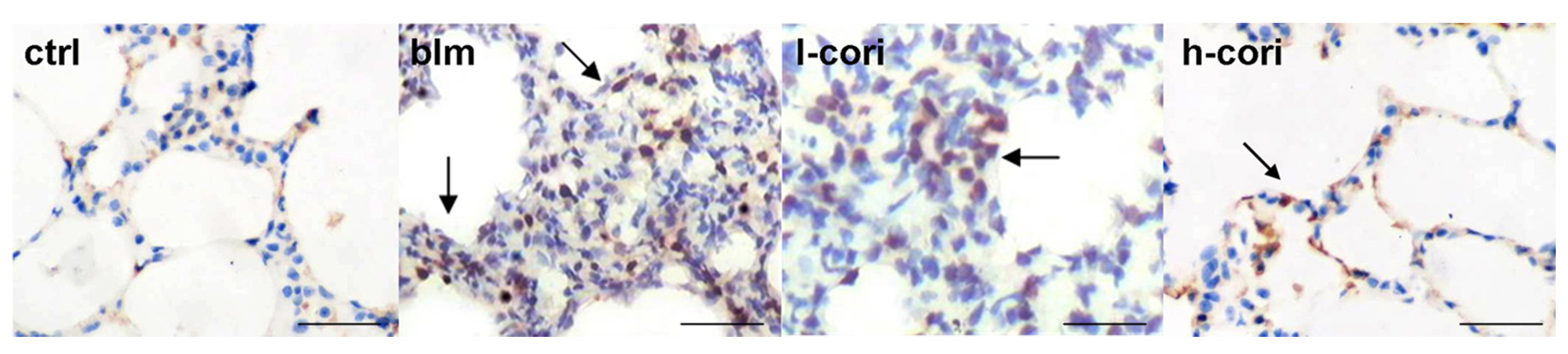
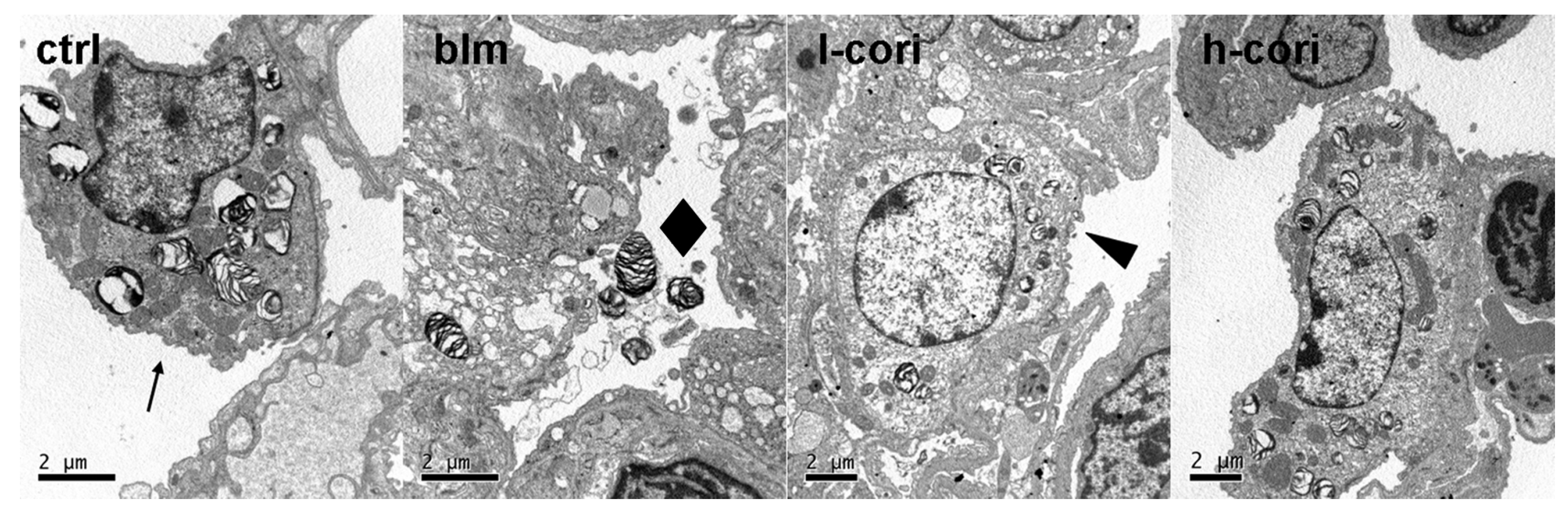
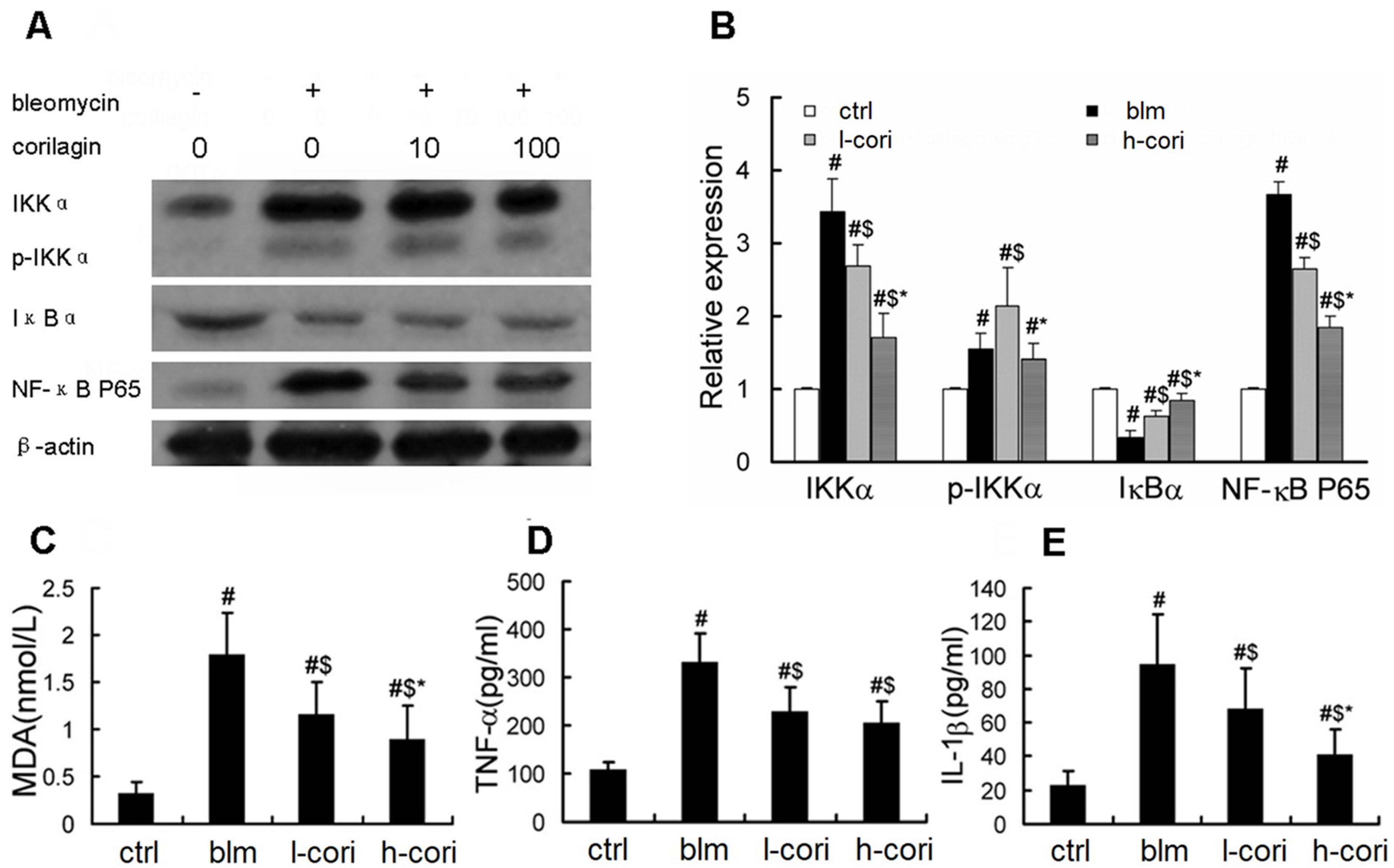
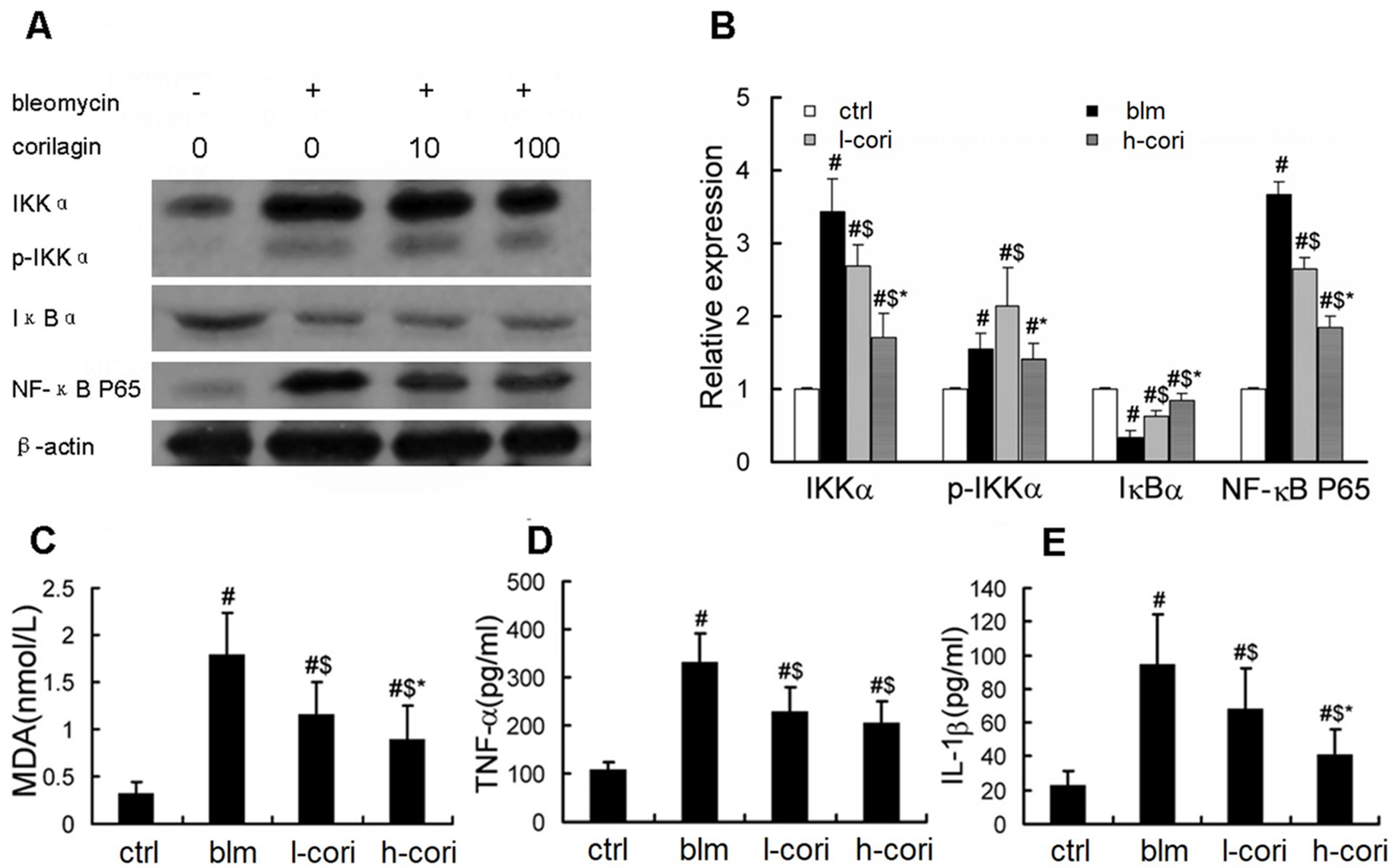
2.4. Corilagin Reduces Bleomycin-Induced Oxidative Responses and NF-κB Activation
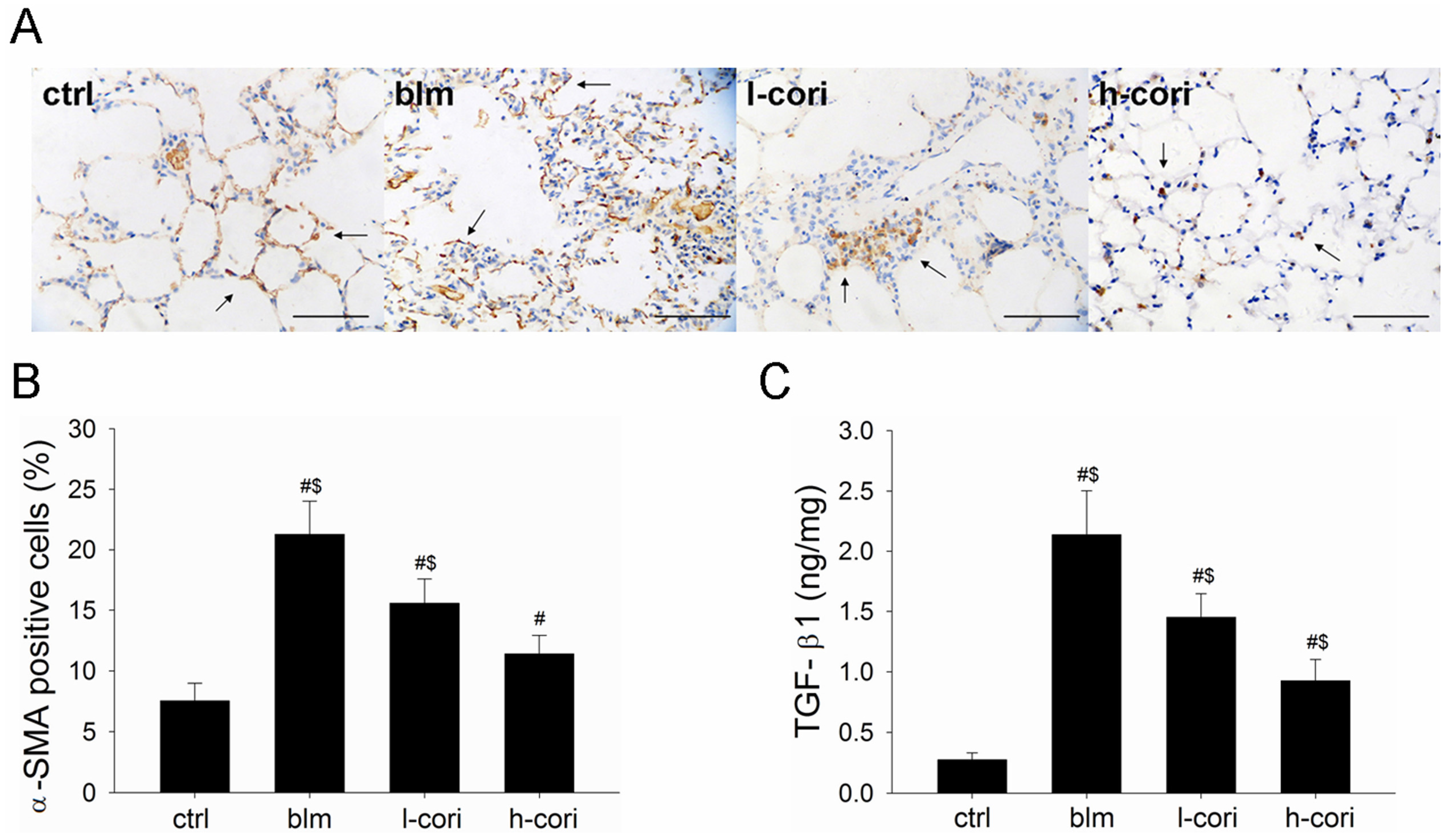
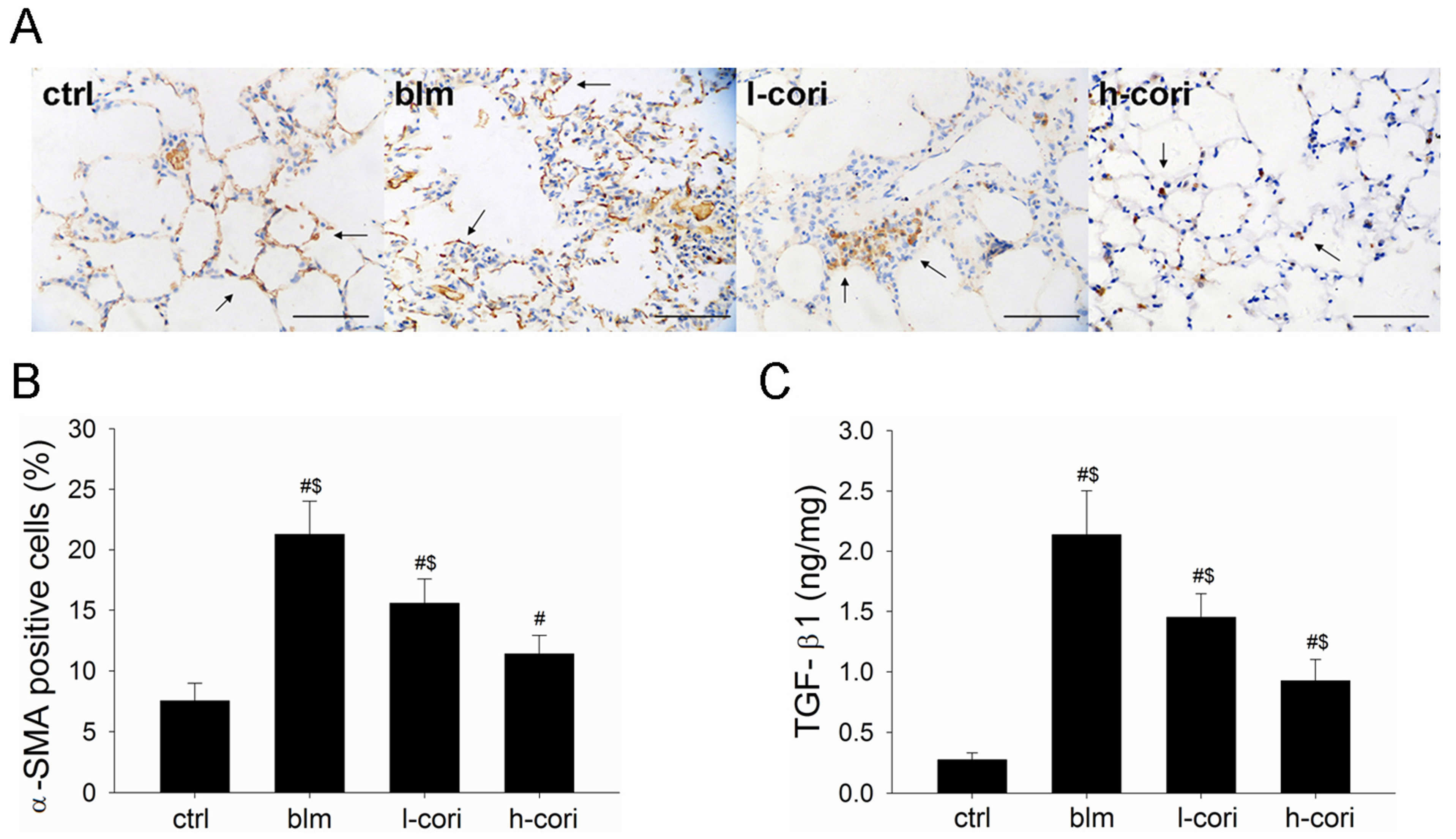
2.5. Corilagin Decreases TGF-β1 Production and α-SMA Expression
2.6. Discussion
3. Experimental Section
3.1. Animals
3.2. Inhalation of Bleomycin and Administration of Corilagin
3.3. Bronchoalveolar Lavage (BAL) and Sample Collection
3.4. Measurement of Oxidative Stress
3.5. Hydroxyproline Assay
3.6. Enzyme-Linked Immunosorbent Assay (ELISA)
3.7. Lung Morphology
3.8. Apoptosis Assay by Terminal Deoxynucleotidyl Transferase (TdT)-Mediated dUTP Nick end Labeling (TUNEL)
3.9. Immunohistochemistry
3.10. Western Blot Assay
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Chen, X.; Zhang, X.; Li, X.; Wang, Z. Who and what should we rely on in early diagnosis of idiopathic pulmonary fibrosis. Eur. Respir. J. 2013, 41, 249–250. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, X.; Kang, Y.; Zeng, Y.; Liu, H.; Chen, X.; Ma, L. Stem cell therapy for idiopathic pulmonary fibrosis: How far are we from the bench to the bedside. J. Biomed. Sci. Eng. 2013, 6, 24–31. [Google Scholar]
- Raghu, G.; Weycker, D.; Edelsberg, J.; Bradford, W.Z.; Oster, G. Incidence and prevalence of idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2006, 174, 810–816. [Google Scholar] [CrossRef]
- Raghu, G.; Collard, H.R.; Egan, J.J.; Martinez, F.J.; Behr, J.; Brown, K.K.; Colby, T.V.; Cordier, J.; Flaherty, K.R.; Lasky, J.A.; et al. An official ATS/ERS/JRS/ALAT statement: Idiopathic pulmonary fibrosis: Evidence-based guidelines for diagnosis and management. Am. J. Respir. Crit. Care Med. 2011, 183, 788–824. [Google Scholar] [CrossRef]
- Barbas-Filho, J.V.; Ferreira, M.A.; Sesso, A.; Kairalla, R.A.; Carvalho, C.R.; Capelozzi, V.L. Evidence of type II pneumocyte apoptosis in the pathogenesis of idiopathic pulmonary fibrosis (IFP)/usual interstitial pneumonia (UIP). J. Clin. Pathol. 2001, 54, 132–138. [Google Scholar] [CrossRef]
- Plataki, M.; Koutsopoulos, A.V.; Darivianaki, K.; Delides, G.; Siafakas, N.M.; Bouros, D. Expression of apoptotic and antiapoptotic markers in epithelial cells in idiopathic pulmonary fibrosis. Chest 2005, 127, 266–274. [Google Scholar] [CrossRef]
- Moore, B.B.; Hogaboam, C.M. Murine models of pulmonary fibrosis. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 294, L152–L160. [Google Scholar] [CrossRef]
- Li, W.; Hu, Y.; Yuan, W.; Li, L.; Huang, W. Comparison of two mouse models of lung fibrosis induced by intratracheal instillation and intratracheal aerosol administration of bleomycin. Nan Fang Yi Ke Da Xue Xue Bao 2012, 32, 221–225. [Google Scholar]
- El-Khouly, D.; El-Bakly, W.M.; Awad, A.S.; El-Mesallamy, H.O.; El-Demerdash, E. Thymoquinone blocks lung injury and fibrosis by attenuating bleomycin-induced oxidative stress and activation of nuclear factor Kappa-B in rats. Toxicology 2012, 302, 106–113. [Google Scholar] [CrossRef]
- Cavarra, E.; Carraro, F.; Fineschi, S.; Naldini, A.; Bartalesi, B.; Pucci, A.; Lungarella, G. Early response to bleomycin is characterized by different cytokine and cytokine receptor profiles in lungs. Am. J. Physiol. Lung Cell Mol. Physiol. 2004, 287, L1186–L1192. [Google Scholar]
- Kalayarasan, S.; Sriram, N.; Sudhandiran, G. Diallyl sulfide attenuates bleomycin-induced pulmonary fibrosis: Critical role of iNOS, NF-kappaB, TNF-alpha and IL-1beta. Life Sci. 2008, 82, 1142–1153. [Google Scholar]
- Degryse, A.L.; Tanjore, H.; Xu, X.C.; Polosukhin, V.V.; Jones, B.R.; Boomershine, C.S.; Ortiz, C.; Sherrill, T.P.; McMahon, F.B.; Gleaves, L.A.; et al. TGFbeta signaling in lung epithelium regulates bleomycin-induced alveolar injury and fibroblast recruitment. Am. J. Physiol. Lung Cell Mol. Physiol. 2011, 300, L887–L897. [Google Scholar] [CrossRef]
- Gharaee-Kermani, M.; Hu, B.; Phan, S.H.; Gyetko, M.R. Recent advances in molecular targets and treatment of idiopathic pulmonary fibrosis: Focus on TGFbeta signaling and the myofibroblast. Curr. Med. Chem. 2009, 16, 1400–1417. [Google Scholar] [CrossRef]
- Azuma, A.; Nukiwa, T.; Tsuboi, E.; Suga, M.; Abe, S.; Nakata, K.; Taguchi, Y.; Nagai, S.; Itoh, H.; Ohi, M.; et al. Double-blind, placebo-controlled trial of pirfenidone in patients with idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2005, 171, 1040–1047. [Google Scholar] [CrossRef]
- Demedts, M.; Behr, J.; Buhl, R.; Costabel, U.; Dekhuijzen, R.; Jansen, HM.; MacNee, W.; Thomeer, M.; Wallaert, B.; Laurent, F.; et al. High-dose acetylcysteine in idiopathic pulmonary fibrosis. N. Engl. J. Med. 2005, 353, 2229–2242. [Google Scholar] [CrossRef]
- Hunninghake, G.W. Antioxidant therapy for idiopathic pulmonary fibrosis. N. Engl. J. Med. 2005, 353, 2285–2287. [Google Scholar] [CrossRef]
- Fujimoto, H.; D’Alessandro-Gabazza, C.N.; Palanki, M.S.; Erdman, P.E.; Takagi, T.; Gabazza, E.C.; Bruno, N.E.; Yano, Y.; Hayashi, T.; Tamaki, S.; et al. Inhibition of nuclear factor-kappaB in T cells suppresses lung fibrosis. Am. J. Respir. Crit. Care Med. 2007, 176, 1251–1260. [Google Scholar] [CrossRef]
- Selman, M.; King, T.E.; Pardo, A. Idiopathic pulmonary fibrosis: Prevailing and evolving hypotheses about its pathogenesis and implications for therapy. Ann. Intern. Med. 2001, 134, 136–151. [Google Scholar] [CrossRef]
- Cheng, J.T.; Lin, T.C.; Hsu, F.L. Antihypertensive effect of corilagin in the rat. Can. J. Physiol. Pharmacol. 1995, 73, 1425–1429. [Google Scholar] [CrossRef]
- Shiota, S.; Shimizu, M.; Sugiyama, J.; Morita, Y.; Mizushima, T.; Tsuchiya, T. Mechanisms of action of corilagin and tellimagrandin I that remarkably potentiate the activity of beta-lactams against methicillin-resistant Staphylococcus aureus. Microbiol. Immunol. 2004, 48, 67–73. [Google Scholar] [CrossRef]
- Duan, W.; Yu, Y.; Zhang, L. Antiatherogenic effects of phyllanthus emblica associated with corilagin and its analogue. Yakugaku Zasshi 2005, 125, 587–591. [Google Scholar] [CrossRef]
- Hau, D.K.; Zhu, G.Y.; Leung, A.K.; Wong, R.S.; Cheng, G.Y.; Lai, P.B.; Tong, S.W.; Lau, F.Y.; Chan, K.W.; Wong, W.Y.; et al. In vivo anti-tumour activity of corilagin on Hep3B hepatocellular carcinoma. Phytomedicine 2010, 18, 11–15. [Google Scholar] [CrossRef]
- Kinoshita, S.; Inoue, Y.; Nakama, S.; Ichiba, T.; Aniya, Y. Antioxidant and hepatoprotective actions of medicinal herb, Terminalia catappa L. from Okinawa Island and its tannin corilagin. Phytomedicine 2007, 14, 755–762. [Google Scholar]
- Dong, X.R.; Luo, M.; Fan, L.; Zhang, T.; Liu, L.; Dong, J.H.; Wu, G. Corilagin inhibits the double strand break-triggered NF-kappaB pathway in irradiated microglial cells. Int. J. Mol. Med. 2010, 25, 531–536. [Google Scholar]
- Guo, Y.J.; Zhao, L.; Li, X.F.; Mei, Y.W.; Zhang, S.L.; Tao, J.Y.; Zhou, Y.; Dong, J.H. Effect of Corilagin on anti-inflammation in HSV-1 encephalitis and HSV-1 infected microglias. Eur. J. Pharmacol. 2010, 635, 79–86. [Google Scholar] [CrossRef]
- Gambari, R.; Borgatti, M.; Lampronti, I.; Fabbri, E.; Brognara, E.; Bianchi, N.; Piccagli, L.; Yuen, M.C.; Kan, C.W.; Hau, D.K.; et al. Corilagin is a potent inhibitor of NF-kappaB activity and downregulates TNF-alpha induced expression of IL-8 gene in cystic fibrosis IB3-1 cells. Int. Immunopharmacol. 2012, 13, 308–315. [Google Scholar] [CrossRef]
- Huang, Y.F.; Chen, S.D.; Wang, Y.F.; Shi, H.X.; Jin, H.H.; Zhang, Y.; Zhao, L. Effects of Corilagin on expression of IL-13 in schistosomiasis liver fibrosis of mice liver tissue and therapeutic effect of anti-fibrosis. Chin. J. Interg. Trad. West Med. Liver Dis. 2011, 21, 156–158. [Google Scholar]
- Sen, C.K.; Khanna, S.; Gordillo, G.; Bagchi, D.; Bagchi, M.; Roy, S. Oxygen, oxidants, and antioxidants in wound healing: An emerging paradigm. Ann. N. Y. Acad. Sci. 2002, 957, 239–249. [Google Scholar] [CrossRef]
- Serrano-Mollar, A.; Nacher, M.; Gay-Jordi, G.; Closa, D.; Xaubet, A.; Bulbena, O. Intratracheal transplantation of alveolar type II cells reverses bleomycin-induced lung fibrosis. Am. J. Respir. Crit. Care Med. 2007, 176, 1261–1268. [Google Scholar] [CrossRef]
- Mouratis, M.A.; Aidinis, V. Modeling pulmonary fibrosis with bleomycin. Curr. Opin. Pulm. Med. 2011, 17, 355–361. [Google Scholar] [CrossRef]
- Warshamana, G.S.; Corti, M.; Brody, A.R. TNF-alpha, PDGF, and TGF-beta(1) expression by primary mouse bronchiolar-alveolar epithelial and mesenchymal cells: Tnf-alpha induces TGF-beta(1). Exp. Mol. Pathol. 2001, 71, 13–33. [Google Scholar] [CrossRef]
- Zhang, K.; Gharaee-Kermani, M.; McGarry, B.; Remick, D.; Phan, S.H. TNF-alpha-mediated lung cytokine networking and eosinophil recruitment in pulmonary fibrosis. J. Immunol. 1997, 158, 954–959. [Google Scholar]
- Mantawy, E.M.; Tadros, M.G.; Awad, A.S.; Hassan, D.A.; El-Demerdash, E. Insights antifibrotic mechanism of methyl palmitate: Impact on nuclear factor kappa B and proinflammatory cytokines. Toxicol. Appl. Pharmacol. 2012, 258, 134–144. [Google Scholar] [CrossRef]
- Kolb, M.; Margetts, P.J.; Anthony, D.C.; Pitossi, F.; Gauldie, J. Transient expression of IL-1beta induces acute lung injury and chronic repair leading to pulmonary fibrosis. J. Clin. Investig. 2001, 107, 1529–1536. [Google Scholar] [CrossRef]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef]
- Cutroneo, K.R. Evidence for TGF-beta1 and bleomycin intracellular signaling through autocrine regulation of Smad 3 binding to the proximal promoter of the Smad 7 gene. J. Cell. Biochem. 2006, 97, 933–939. [Google Scholar] [CrossRef]
- Bors, W.; Michel, C. Chemistry of the antioxidant effect of polyphenols. Ann. N. Y. Acad. Sci. 2002, 957, 57–69. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, S.L.; Tao, J.Y.; Pang, R.; Jin, F.; Guo, Y.J.; Dong, J.H.; Ye, P.; Zhao, H.Y.; Zheng, G.H. Preliminary exploration on anti-inflammatory mechanism of Corilagin (beta-1-O-galloyl-3,6-(R)-hexahydroxydiphenoyl-d-glucose) in vitro. Int. Immunopharmacol. 2008, 8, 1059–1064. [Google Scholar] [CrossRef]
- Blenn, C.; Wyrsch, P.; Althaus, F.R. The ups and downs of tannins as inhibitors of poly(ADP-ribose)glycohydrolase. Molecules 2011, 16, 1854–1877. [Google Scholar] [CrossRef] [Green Version]
- Grzanna, R.; Lindmark, L.; Frondoza, C.G. Ginger—An herbal medicinal product with broad anti-inflammatory actions. J. Med. Food 2005, 8, 125–132. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.N.; Hu, X.F.; Chen, X.L.; Wang, X.R.; Zhao, T.T.; Peng, M.J.; Zou, P. Effects of pentoxifylline on Wnt/beta-catenin signaling in mice chronically exposed to cigarette smoke. Chin. Med. J. 2010, 123, 2688–2694. [Google Scholar]
- Ashcroft, T.; Simpson, J.M.; Timbrell, V. Simple method of estimating severity of pulmonary fibrosis on a numerical scale. J. Clin. Pathol. 1988, 41, 467–470. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, Z.; Guo, Q.-Y.; Zhang, X.-J.; Li, X.; Li, W.-T.; Ma, X.-T.; Ma, L.-J. Corilagin Attenuates Aerosol Bleomycin-Induced Experimental Lung Injury. Int. J. Mol. Sci. 2014, 15, 9762-9779. https://doi.org/10.3390/ijms15069762
Wang Z, Guo Q-Y, Zhang X-J, Li X, Li W-T, Ma X-T, Ma L-J. Corilagin Attenuates Aerosol Bleomycin-Induced Experimental Lung Injury. International Journal of Molecular Sciences. 2014; 15(6):9762-9779. https://doi.org/10.3390/ijms15069762
Chicago/Turabian StyleWang, Zheng, Qiong-Ya Guo, Xiao-Ju Zhang, Xiao Li, Wen-Ting Li, Xi-Tao Ma, and Li-Jun Ma. 2014. "Corilagin Attenuates Aerosol Bleomycin-Induced Experimental Lung Injury" International Journal of Molecular Sciences 15, no. 6: 9762-9779. https://doi.org/10.3390/ijms15069762