Human Mesenchymal Stem Cells Modulate Inflammatory Cytokines after Spinal Cord Injury in Rat


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Spinal Cord Injury and Cell Transplantation

2.2. Behavioral Testing
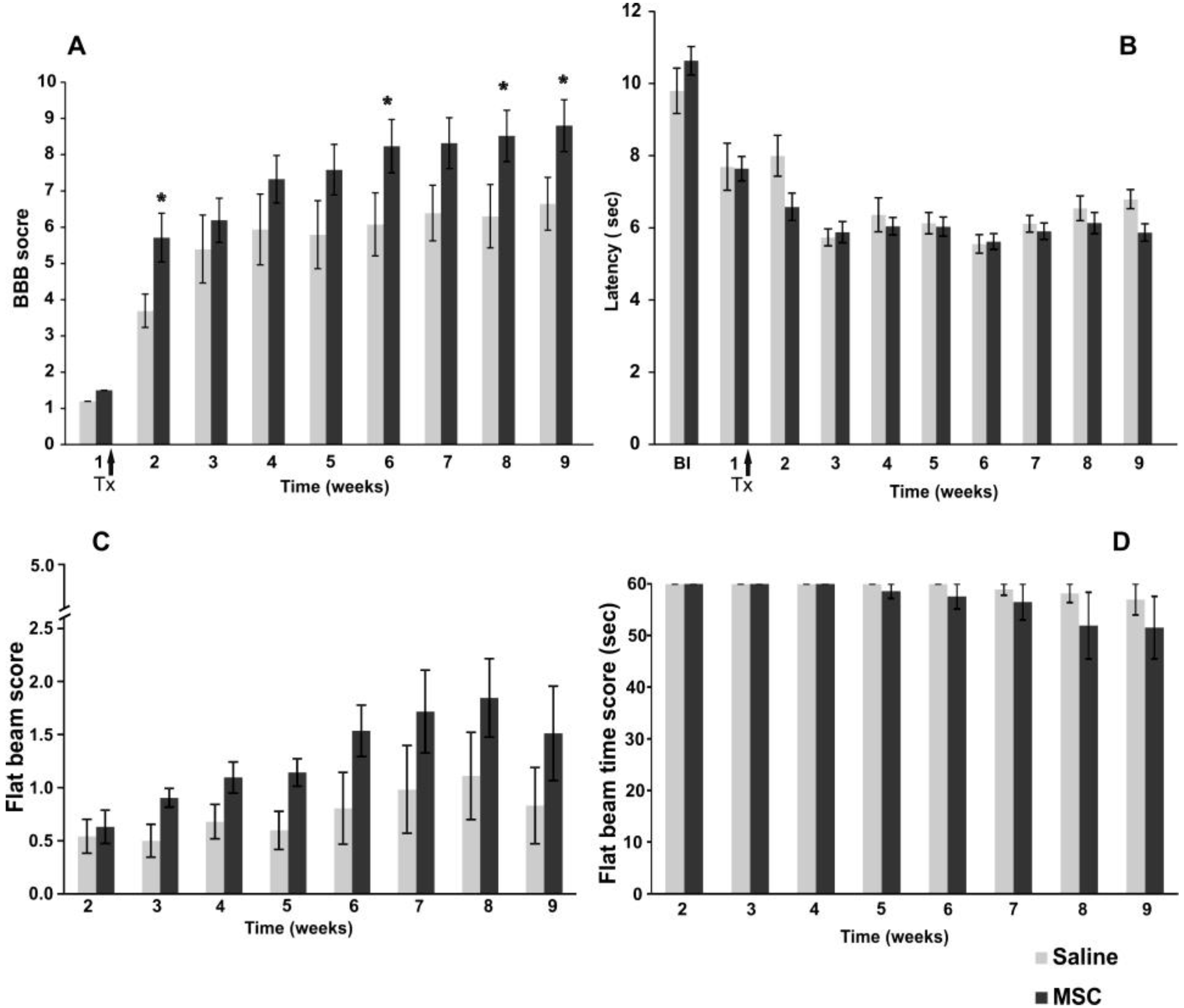
2.2.1. BBB Test
2.2.2. Plantar Test
2.2.3. Flat Beam Test

2.3. Histology and Immunohistochemistry
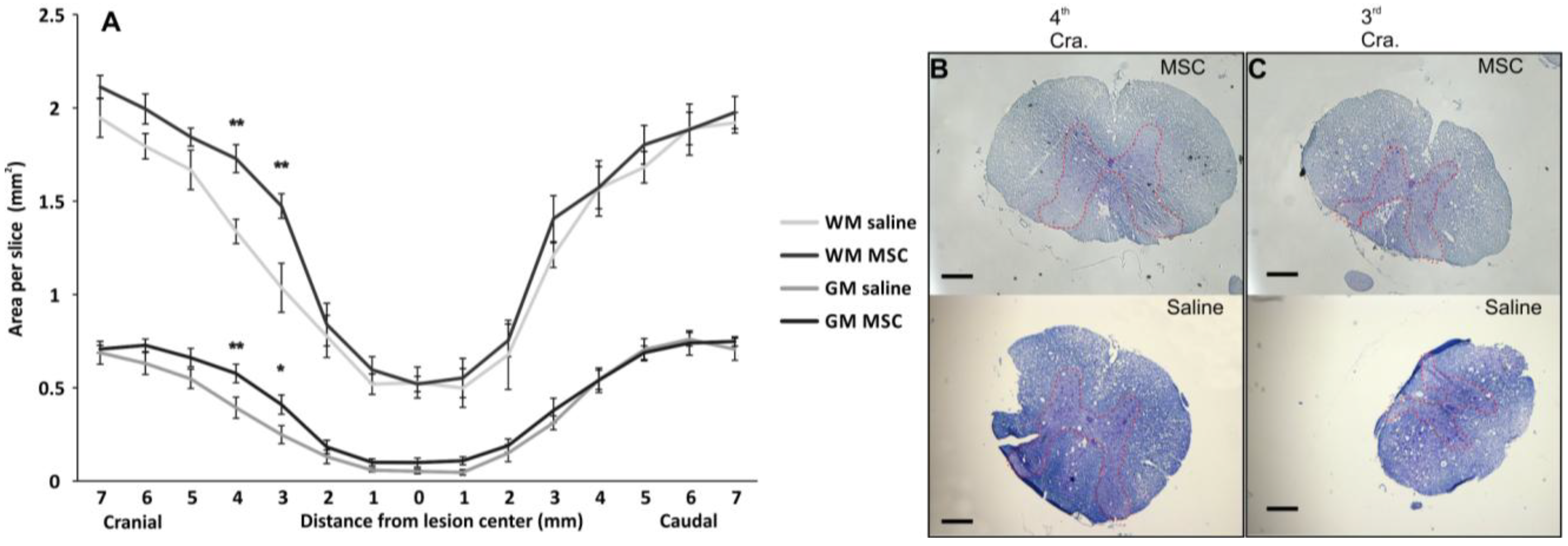
2.3.1. Grey/White Matter Sparing

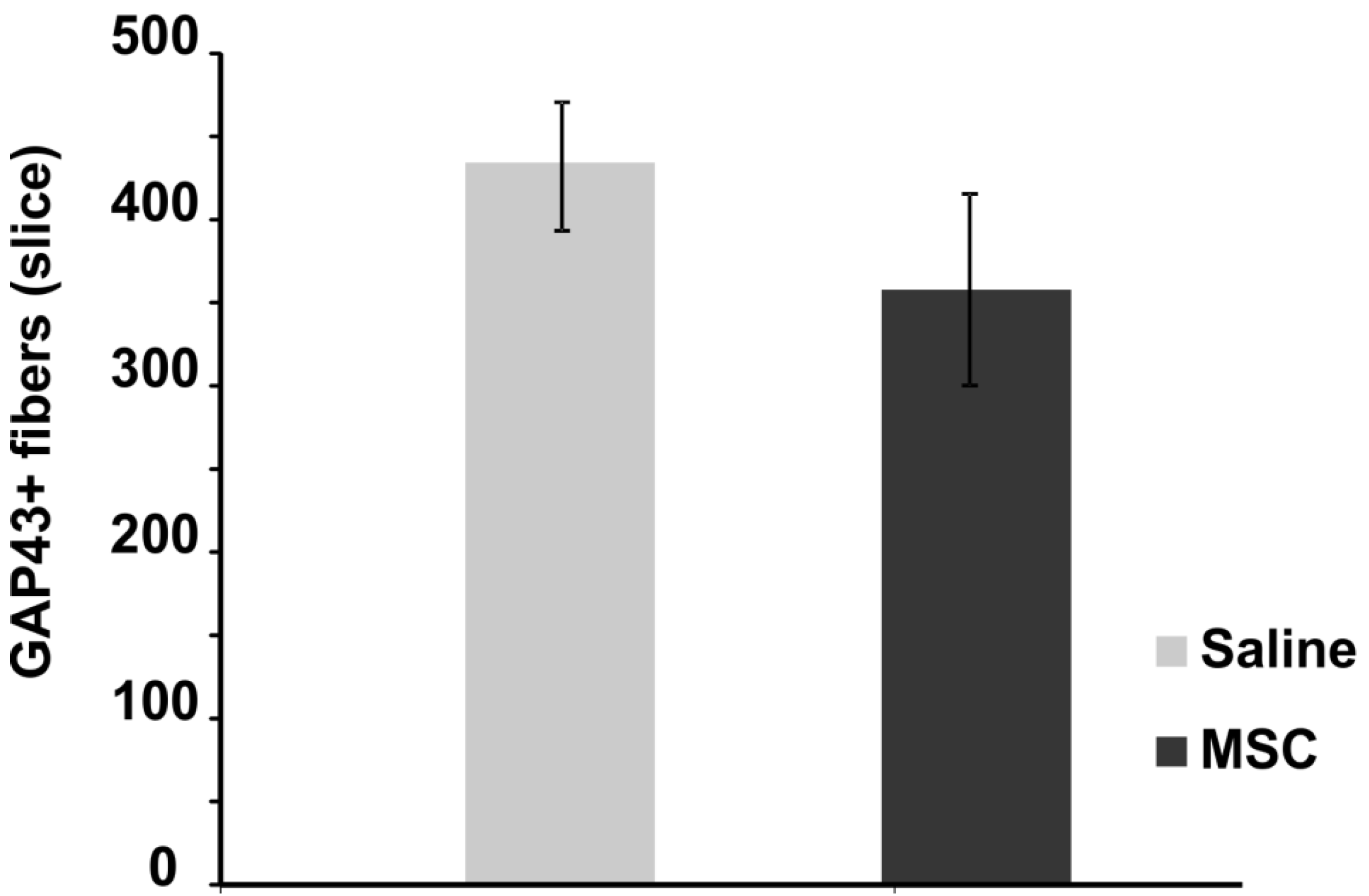
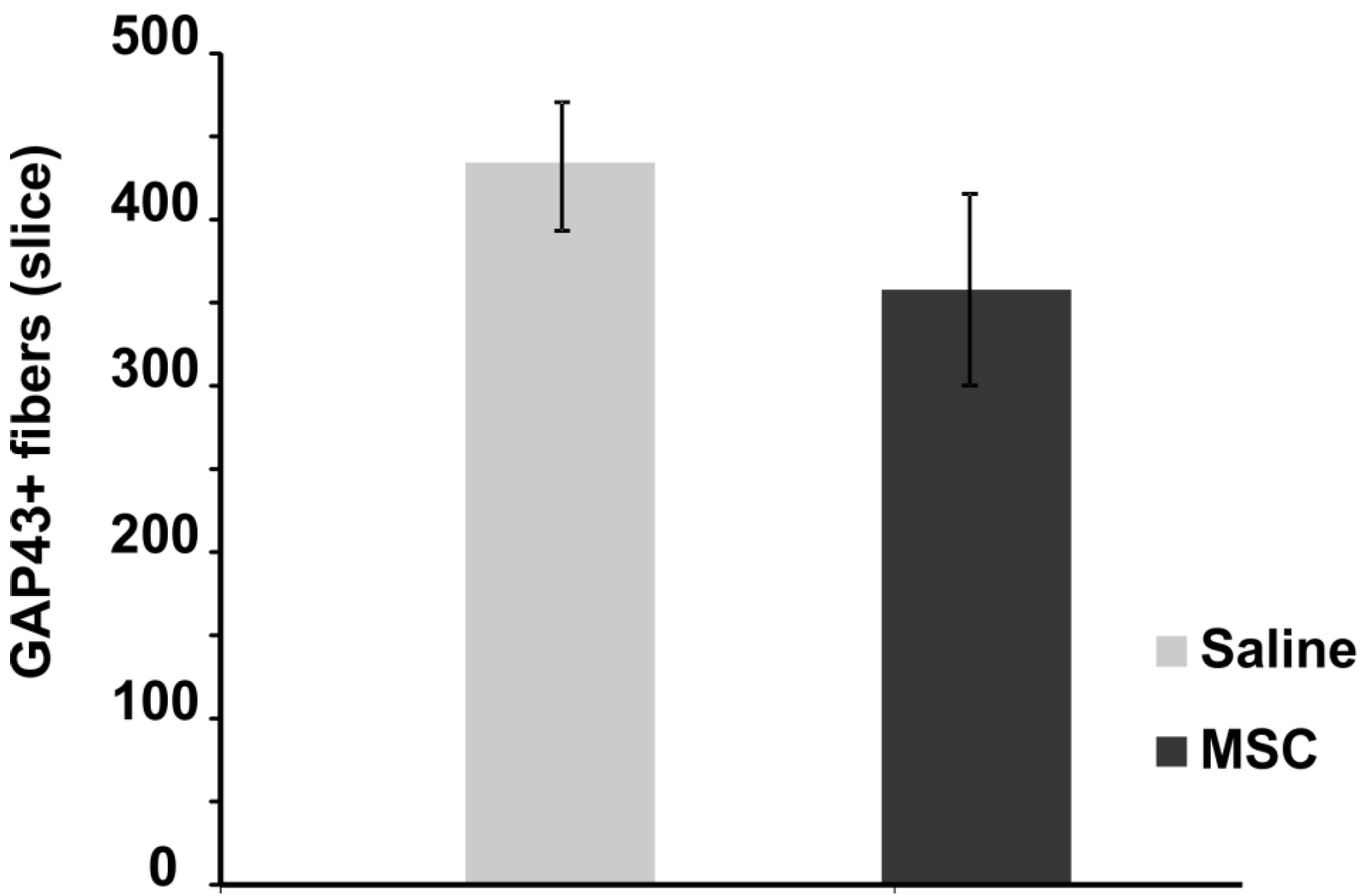
2.3.2. Axonal Sprouting
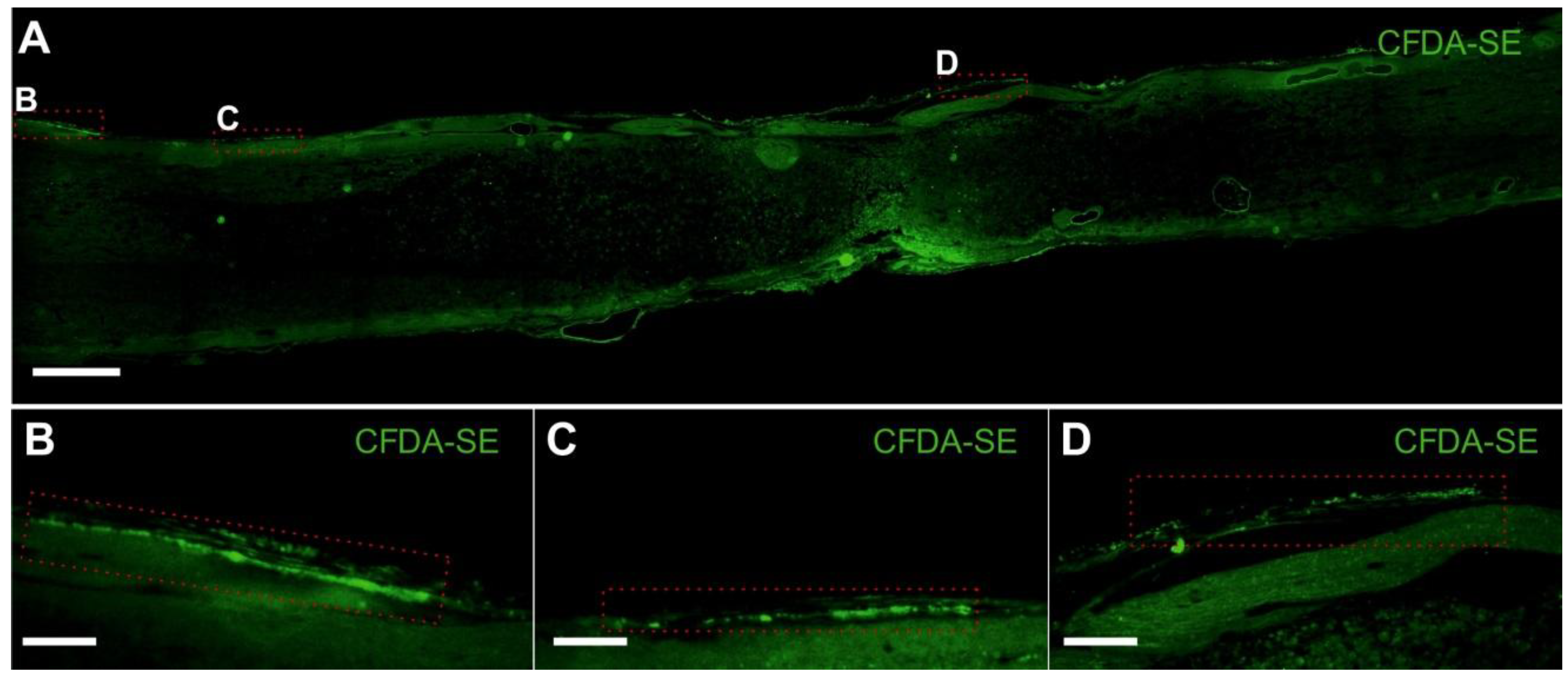
2.3.3. Blood Vessel and Axonal Support
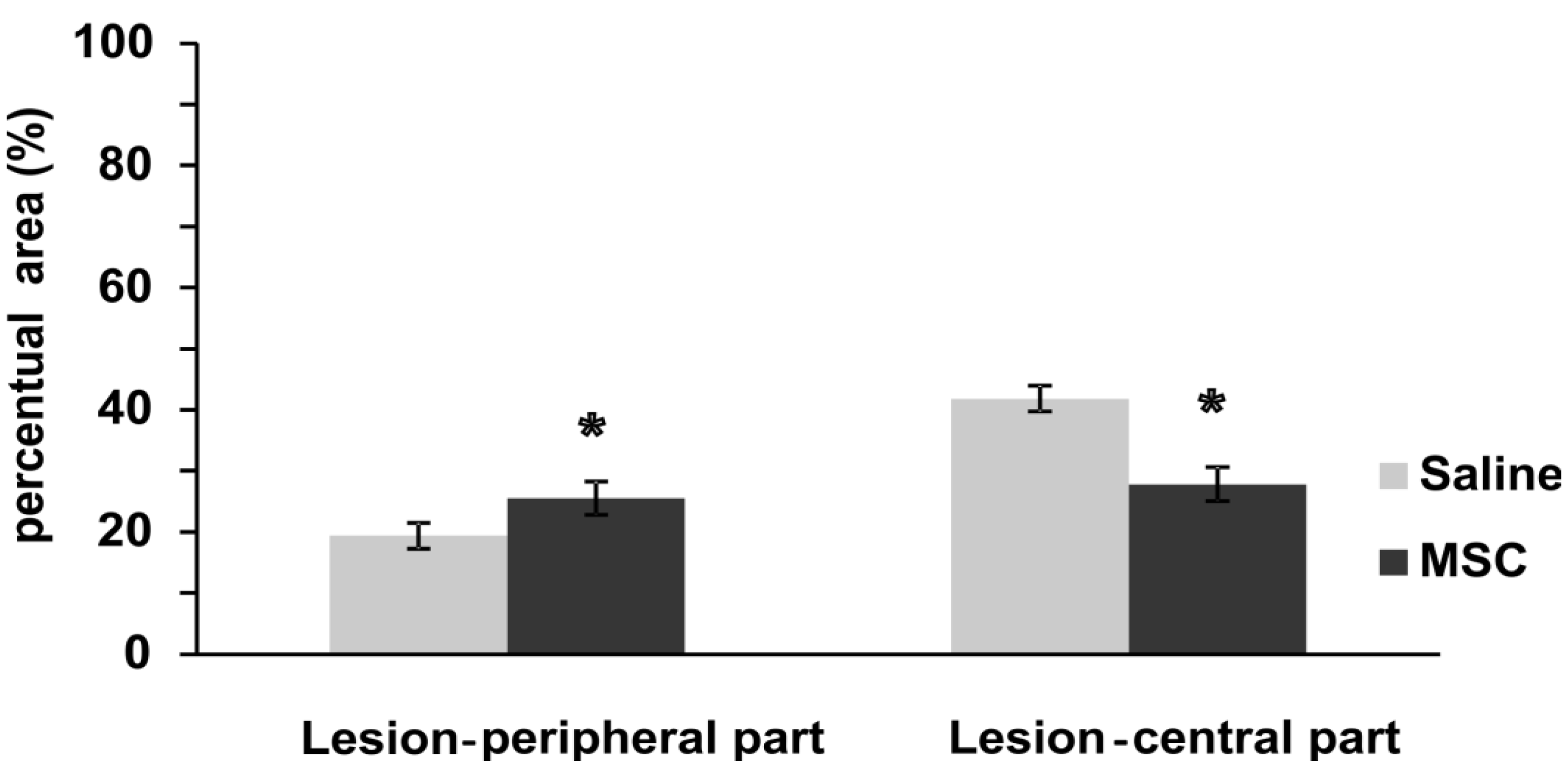
2.3.4. Glial Scar Re-Modulation

2.4. Gene Expression

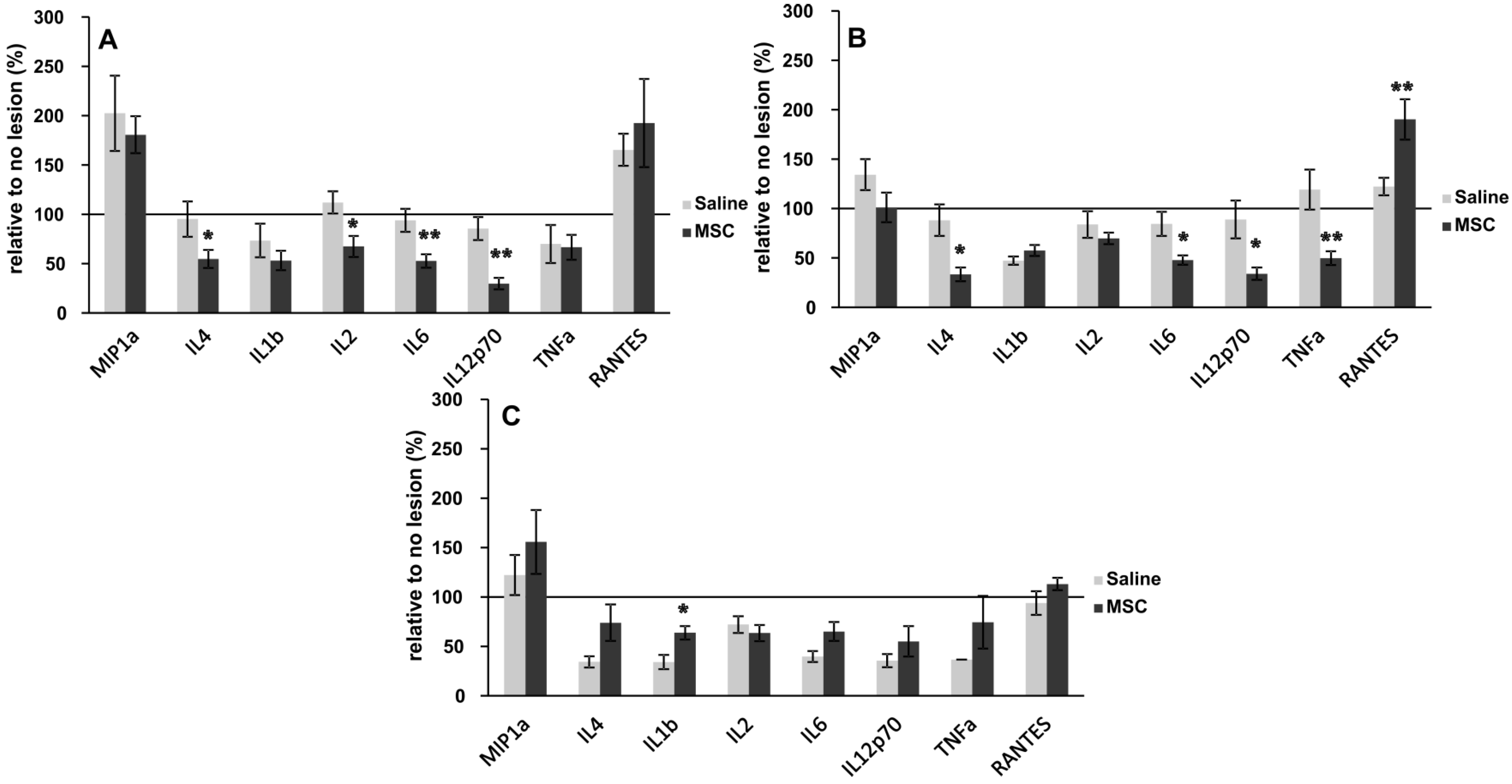
2.5. Inflammatory Cytokine

3. Experimental
3.1. Animals
3.1.1. Spinal Cord Injury and Cell Transplantation
3.2. Behavioral Testing
3.2.1. BBB Test
3.2.2. Plantar Test
3.2.3. Flat Beam Test
3.3. Histology and Immunohistochemistry
3.3.1. Grey/White Matter Sparing
3.3.2. Axonal Sprouting
3.3.3. Axonal Ingrowth
3.3.4. Blood Vessel Ingrowth
3.3.5. Glial Scar
3.4. Gene Expression
3.5. Cytokines
3.6. Statistical Evaluation
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Oliveri, R.S.; Bello, S.; Biering-Sorensen, F. Mesenchymal stem cells improve locomotor recovery in traumatic spinal cord injury: Systematic review with meta-analyses of rat models. Neurobiol. Dis. 2014, 62, 338–353. [Google Scholar] [CrossRef]
- Urdzikova, L.; Jendelova, P.; Glogarova, K.; Burian, M.; Hajek, M.; Sykova, E. Transplantation of bone marrow stem cells as well as mobilization by granulocyte-colony stimulating factor promotes recovery after spinal cord injury in rats. J. Neurotrauma 2006, 23, 1379–1391. [Google Scholar] [CrossRef]
- Coutts, M.; Keirstead, H.S. Stem cells for the treatment of spinal cord injury. Exp. Neurol. 2008, 209, 368–377. [Google Scholar] [CrossRef]
- Garbuzova-Davis, S.; Willing, A.E.; Saporta, S.; Bickford, P.C.; Gemma, C.; Chen, N.; Sanberg, C.D.; Klasko, S.K.; Borlongan, C.V.; Sanberg, P.R. Novel cell therapy approaches for brain repair. Prog. Brain Res. 2006, 157, 207–222. [Google Scholar] [CrossRef]
- Joyce, N.; Annett, G.; Wirthlin, L.; Olson, S.; Bauer, G.; Nolta, J.A. Mesenchymal stem cells for the treatment of neurodegenerative disease. Regen. Med. 2010, 5, 933–946. [Google Scholar] [CrossRef]
- Hardy, S.A.; Maltman, D.J.; Przyborski, S.A. Mesenchymal stem cells as mediators of neural differentiation. Curr. Stem Cell Res. Ther. 2008, 3, 43–52. [Google Scholar] [CrossRef]
- Nishio, Y.; Koda, M.; Kamada, T.; Someya, Y.; Yoshinaga, K.; Okada, S.; Harada, H.; Okawa, A.; Moriya, H.; Yamazaki, M. The use of hemopoietic stem cells derived from human umbilical cord blood to promote restoration of spinal cord tissue and recovery of hindlimb function in adult rats. J. Neurosurg. Spine 2006, 5, 424–433. [Google Scholar] [CrossRef]
- English, K. Mechanisms of mesenchymal stromal cell immunomodulation. Immunol. Cell Biol. 2013, 91, 19–26. [Google Scholar] [CrossRef]
- Stagg, J.; Galipeau, J. Mechanisms of immune modulation by mesenchymal stromal cells and clinical translation. Curr. Mol. Med. 2013, 13, 856–867. [Google Scholar] [CrossRef]
- Weiss, D.J.; Casaburi, R.; Flannery, R.; LeRoux-Williams, M.; Tashkin, D.P. A placebo-controlled, randomized trial of mesenchymal stem cells in copd. Chest 2013, 143, 1590–1598. [Google Scholar] [CrossRef]
- Karussis, D.; Karageorgiou, C.; Vaknin-Dembinsky, A.; Gowda-Kurkalli, B.; Gomori, J. M.; Kassis, I.; Bulte, J.W.; Petrou, P.; Ben-Hur, T.; Abramsky, O.; et al. Safety and immunological effects of mesenchymal stem cell transplantation in patients with multiple sclerosis and amyotrophic lateral sclerosis. Arch. Neurol. 2010, 67, 1187–1194. [Google Scholar]
- Vanecek, V.; Zablotskii, V.; Forostyak, S.; Ruzicka, J.; Herynek, V.; Babic, M.; Jendelova, P.; Kubinova, S.; Dejneka, A.; Sykova, E. Highly efficient magnetic targeting of mesenchymal stem cells in spinal cord injury. Int. J. Nanomed. 2012, 7, 3719–3730. [Google Scholar]
- Zhilai, Z.; Hui, Z.; Anmin, J.; Shaoxiong, M.; Bo, Y.; Yinhai, C. A combination of taxol infusion and human umbilical cord mesenchymal stem cells transplantation for the treatment of rat spinal cord injury. Brain Res. 2012, 1481, 79–89. [Google Scholar]
- Cizkova, D.; Novotna, I.; Slovinska, L.; Vanicky, I.; Jergova, S.; Rosocha, J.; Radonak, J. Repetitive intrathecal catheter delivery of bone marrow mesenchymal stromal cells improves functional recovery in a rat model of contusive spinal cord injury. J. Neurotrauma 2011, 28, 1951–1961. [Google Scholar] [CrossRef]
- Mothe, A.J.; Bozkurt, G.; Catapano, J.; Zabojova, J.; Wang, X.; Keating, A.; Tator, C.H. Intrathecal transplantation of stem cells by lumbar puncture for thoracic spinal cord injury in the rat. Spinal Cord 2011, 49, 967–973. [Google Scholar] [CrossRef]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A sensitive and reliable locomotor rating scale for open field testing in rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar]
- Hodgetts, S.I.; Simmons, P.J.; Plant, G.W. Human mesenchymal precursor cells (stro-1(+)) from spinal cord injury patients improve functional recovery and tissue sparing in an acute spinal cord injury rat model. Cell Transplant. 2013, 22, 393–412. [Google Scholar] [CrossRef]
- Ritfeld, G.J.; Nandoe Tewarie, R.D.; Vajn, K.; Rahiem, S.T.; Hurtado, A.; Wendell, D.F.; Roos, R.A.; Oudega, M. Bone marrow stromal cell-mediated tissue sparing enhances functional repair after spinal cord contusion in adult rats. Cell Transplant. 2012, 21, 1561–1575. [Google Scholar] [CrossRef]
- Urdzikova, L.; Vanicky, I. Post-traumatic moderate systemic hyperthermia worsens behavioural outcome after spinal cord injury in the rat. Spinal Cord 2006, 44, 113–119. [Google Scholar] [CrossRef]
- Amemori, T.; Jendelova, P.; Ruzickova, K.; Arboleda, D.; Sykova, E. Co-transplantation of olfactory ensheathing glia and mesenchymal stromal cells does not have synergistic effects after spinal cord injury in the rat. Cytotherapy 2010, 12, 212–225. [Google Scholar] [CrossRef]
- Arboleda, D.; Forostyak, S.; Jendelova, P.; Marekova, D.; Amemori, T.; Pivonkova, H.; Masinova, K.; Sykova, E. Transplantation of predifferentiated adipose-derived stromal cells for the treatment of spinal cord injury. Cell. Mol. Neurobiol. 2011, 31, 1113–1122. [Google Scholar] [CrossRef]
- Pourheydar, B.; Joghataei, M.T.; Bakhtiari, M.; Mehdizadeh, M.; Yekta, Z.; Najafzadeh, N. Co-transplantation of bone marrow stromal cells with schwann cells evokes mechanical allodynia in the contusion model of spinal cord injury in rats. Cell J. 2012, 13, 213–222. [Google Scholar]
- Paul, C.; Samdani, A.F.; Betz, R.R.; Fischer, I.; Neuhuber, B. Grafting of human bone marrow stromal cells into spinal cord injury: A comparison of delivery methods. Spine 2009, 34, 328–334. [Google Scholar] [CrossRef]
- Hejcl, A.; Ruzicka, J.; Kapcalova, M.; Turnovcova, K.; Krumbholcova, E.; Pradny, M.; Michalek, J.; Cihlar, J.; Jendelova, P.; Sykova, E. Adjusting the chemical and physical properties of hydrogels leads to improved stem cell survival and tissue ingrowth in spinal cord injury reconstruction: A comparative study of four methacrylate hydrogels. Stem Cells Dev. 2013, 22, 2794–2805. [Google Scholar] [CrossRef]
- Hejcl, A.; Sedy, J.; Kapcalova, M.; Toro, D.A.; Amemori, T.; Lesny, P.; Likavcanova-Masinova, K.; Krumbholcova, E.; Pradny, M.; Michalek, J.; et al. Hpma-rgd hydrogels seeded with mesenchymal stem cells improve functional outcome in chronic spinal cord injury. Stem Cells Dev. 2010, 19, 1535–1546. [Google Scholar] [CrossRef]
- Renault-Mihara, F.; Okada, S.; Shibata, S.; Nakamura, M.; Toyama, Y.; Okano, H. Spinal cord injury: Emerging beneficial role of reactive astrocytes' migration. Int. J. Biochem. Cell Biol. 2008, 40, 1649–1653. [Google Scholar] [CrossRef]
- Lukovic, D.; Moreno Manzano, V.; Stojkovic, M.; Bhattacharya, S.S.; Erceg, S. Concise review: Human pluripotent stem cells in the treatment of spinal cord injury. Stem Cells 2012, 30, 1787–1792. [Google Scholar] [CrossRef]
- Hausmann, O.N. Post-traumatic inflammation following spinal cord injury. Spinal Cord 2003, 41, 369–378. [Google Scholar] [CrossRef]
- Cusimano, M.; Biziato, D.; Brambilla, E.; Donega, M.; Alfaro-Cervello, C.; Snider, S.; Salani, G.; Pucci, F.; Comi, G.; Garcia-Verdugo, J.M.; et al. Transplanted neural stem/precursor cells instruct phagocytes and reduce secondary tissue damage in the injured spinal cord. Brain 2012, 135, 447–460. [Google Scholar] [CrossRef]
- Ren, Y.; Young, W. Managing inflammation after spinal cord injury through manipulation of macrophage function. Neural Plast. 2013, 2013, 945034. [Google Scholar]
- Hirai, T.; Uchida, K.; Nakajima, H.; Guerrero, A.R.; Takeura, N.; Watanabe, S.; Sugita, D.; Yoshida, A.; Johnson, W.E.; Baba, H. The prevalence and phenotype of activated microglia/macrophages within the spinal cord of the hyperostotic mouse (twy/twy) changes in response to chronic progressive spinal cord compression: Implications for human cervical compressive myelopathy. PLoS One 2013, 8, e64528. [Google Scholar] [CrossRef]
- Krausgruber, T.; Blazek, K.; Smallie, T.; Alzabin, S.; Lockstone, H.; Sahgal, N.; Hussell, T.; Feldmann, M.; Udalova, I.A. Irf5 promotes inflammatory macrophage polarization and th1-th17 responses. Nat. Immunol. 2011, 12, 231–238. [Google Scholar] [CrossRef]
- Nakajima, H.; Uchida, K.; Rodriguez Guerrero, A.; Watanabe, S.; Sugita, D.; Takeura, N.; Yoshida, A.; Long, G.; Wright, K.; Johnson, E.; et al. Transplantation of mesenchymal stem cells promotes the alternative pathway of macrophage activation and functional recovery after spinal cord injury. J. Neurotrauma 2012, 29, 1614–1625. [Google Scholar]
- Greish, S.; Abogresha, N.; Abdel-Hady, Z.; Zakaria, E.; Ghaly, M.; Hefny, M. Human umbilical cord mesenchymal stem cells as treatment of adjuvant rheumatoid arthritis in a rat model. World J. Stem Cells 2012, 4, 101–109. [Google Scholar]
- Kim, J.; Hematti, P. Mesenchymal stem cell-educated macrophages: A novel type of alternatively activated macrophages. Exp. Hematol. 2009, 37, 1445–1453. [Google Scholar]
- Li, M.; Ikehara, S. Bone-marrow-derived mesenchymal stem cells for organ repair. Stem Cells Int. 2013, 2013, 1–8. [Google Scholar]
- De la Calle, J.L.; Paino, C.L. A procedure for direct lumbar puncture in rats. Brain Res. Bull. 2002, 59, 245–250. [Google Scholar]
- Machova Urdzikova, L.; Sedlacek, R.; Suchy, T.; Amemori, T.; Ruzicka, J.; Lesny, P.; Havlas, V.; Sykova, E.; Jendelova, P. Human multipotent mesenchymal stem cells improve healing after collagenase tendon injury in the rat. Biomed. Eng. Online 2014, 13, 42. [Google Scholar] [CrossRef]
- Goldstein, B.; Little, J.W.; Harris, R.M. Axonal sprouting following incomplete spinal cord injury: An experimental model. J. Spinal Cord Med. 1997, 20, 200–206. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Urdzíková, L.M.; Růžička, J.; LaBagnara, M.; Kárová, K.; Kubinová, Š.; Jiráková, K.; Murali, R.; Syková, E.; Jhanwar-Uniyal, M.; Jendelová, P. Human Mesenchymal Stem Cells Modulate Inflammatory Cytokines after Spinal Cord Injury in Rat. Int. J. Mol. Sci. 2014, 15, 11275-11293. https://doi.org/10.3390/ijms150711275
Urdzíková LM, Růžička J, LaBagnara M, Kárová K, Kubinová Š, Jiráková K, Murali R, Syková E, Jhanwar-Uniyal M, Jendelová P. Human Mesenchymal Stem Cells Modulate Inflammatory Cytokines after Spinal Cord Injury in Rat. International Journal of Molecular Sciences. 2014; 15(7):11275-11293. https://doi.org/10.3390/ijms150711275
Chicago/Turabian StyleUrdzíková, Lucia Machová, Jiří Růžička, Michael LaBagnara, Kristýna Kárová, Šárka Kubinová, Klára Jiráková, Raj Murali, Eva Syková, Meena Jhanwar-Uniyal, and Pavla Jendelová. 2014. "Human Mesenchymal Stem Cells Modulate Inflammatory Cytokines after Spinal Cord Injury in Rat" International Journal of Molecular Sciences 15, no. 7: 11275-11293. https://doi.org/10.3390/ijms150711275