Cloning and Expression Characteristics of the Pig Stra8 Gene



Abstract
:1. Introduction
2. Results and Discussion
2.1. Cloning of Pig Stra8


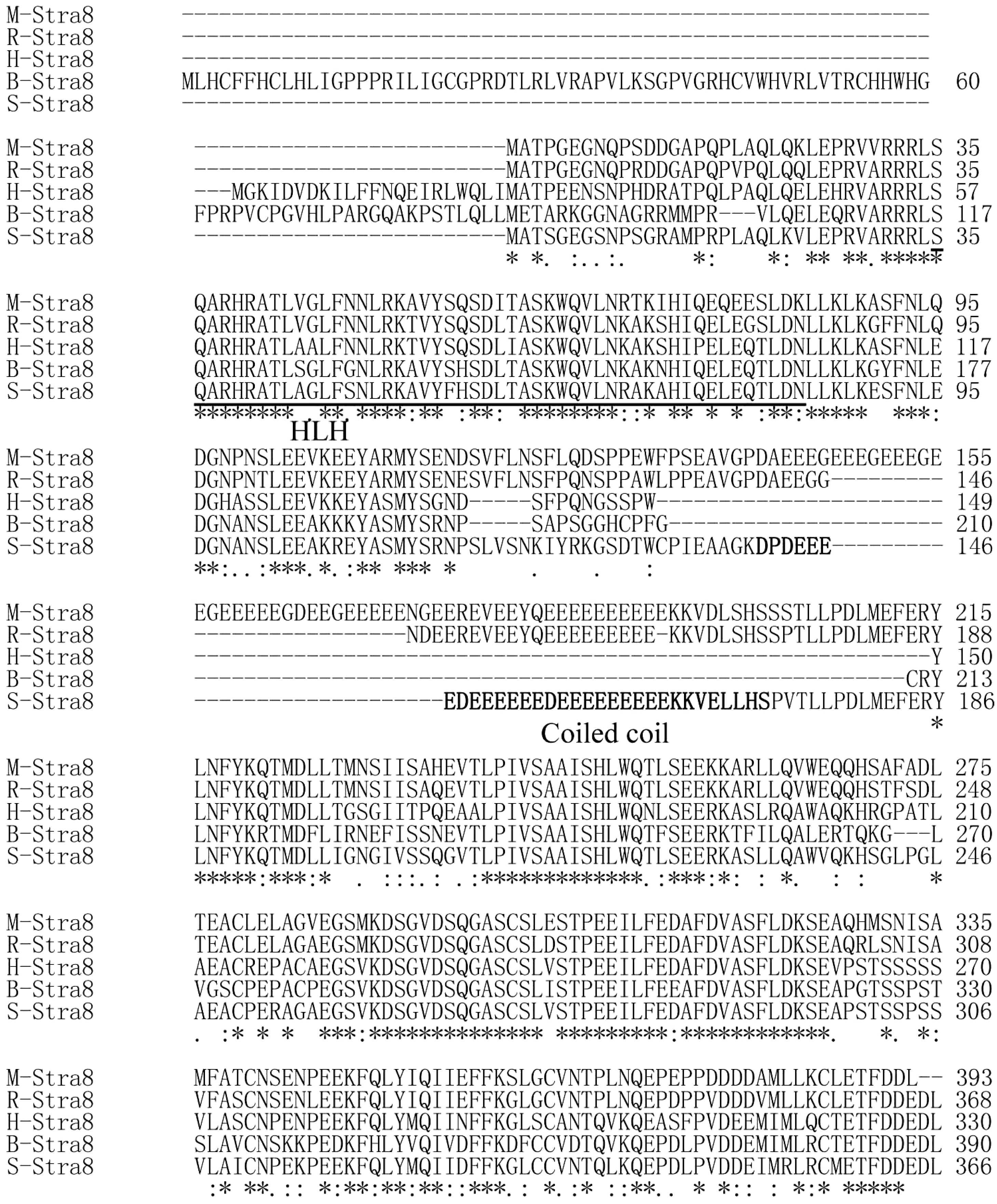
2.2. Multiple Sequence Alignments and Phylogenetic Relationships

{kind=link}
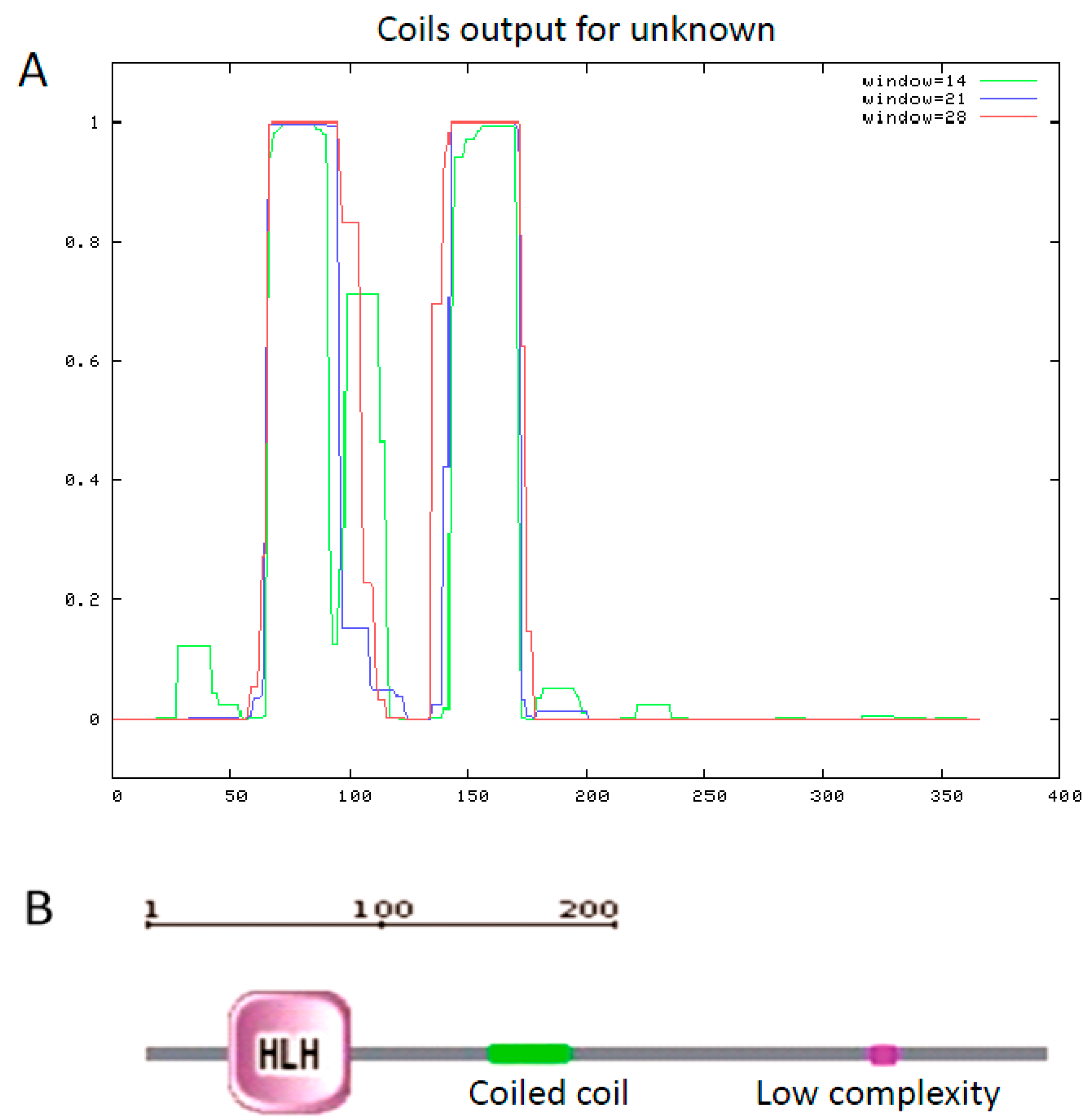{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Similarity | GenBank Accession Number |
|---|---|---|
| Homo sapiens | 66.5% | NM_182489.1 |
| Rattus norvegicus | 72.1% | XM_575429.2 |
| Mus musculus | 78.4% | NM_009292.1 |
| Bos taurus | 57.8% | XM_001253649.1 |
| Canis lupus familiaris | 66.4% | XM_847727.2 |
| Gallus gallus | 45.4% | XM_416179.3 |
| Equus caballus | 75.0% | XM_001914766.1 |
| Pongo abelii | 66.8% | XM_002818472.1 |
| Pan troglodytes | 66.6% | XM_001144872.2 |


2.3. Stra8 mRNA Distribution

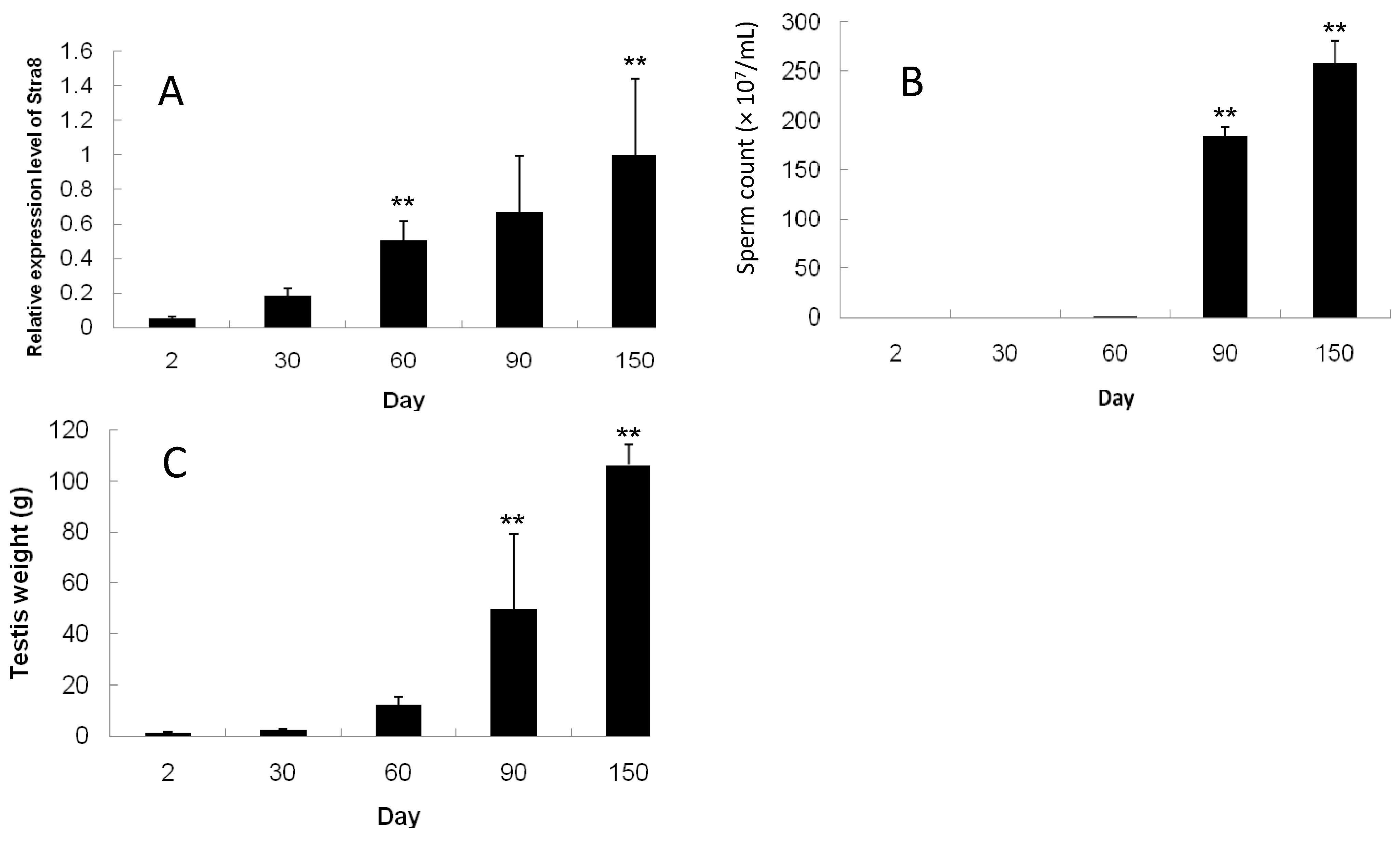
2.4. Developmental Expression Pattern of Stra8, Testes Weight, and Sperm Count
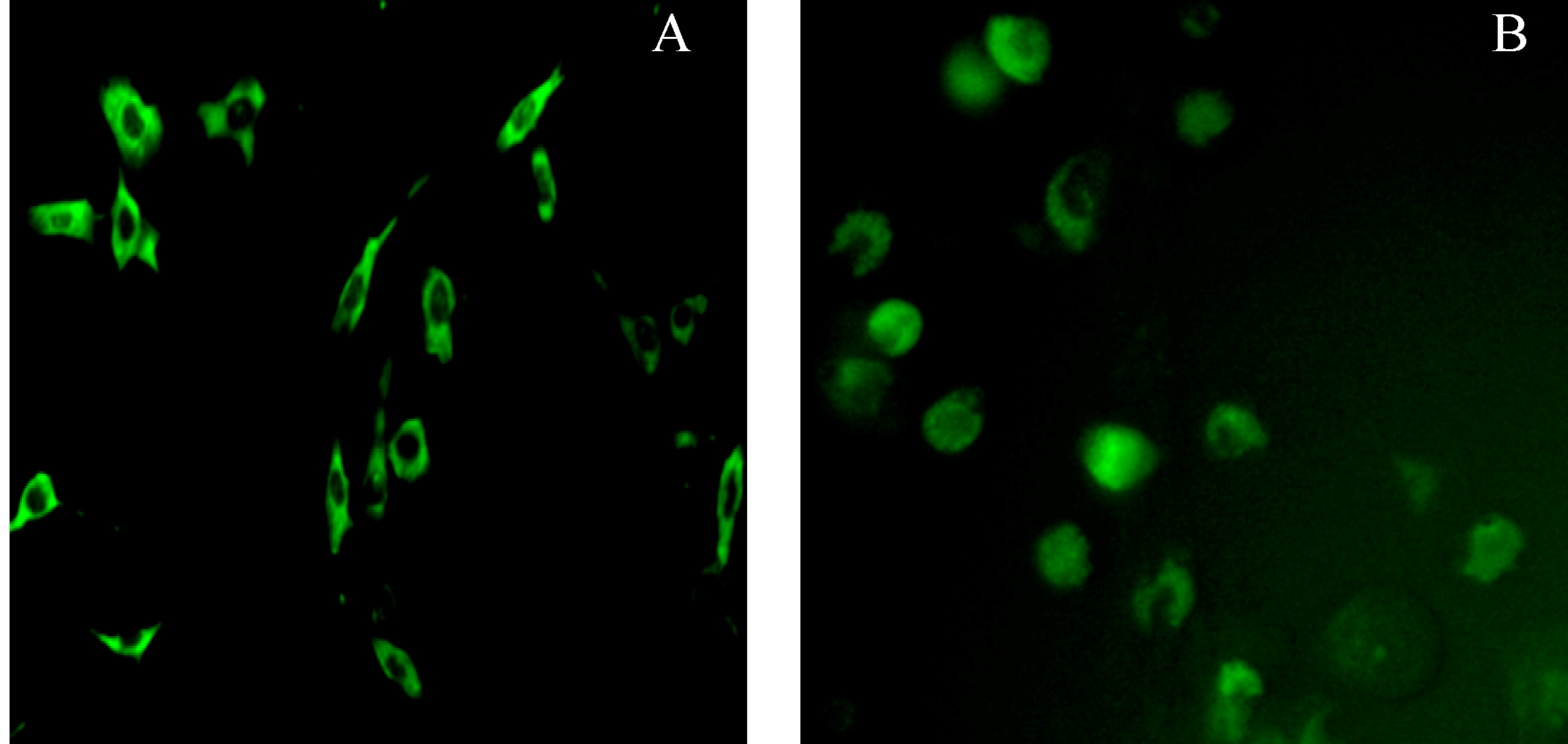
2.5. Subcellular Localization of Pig Stra8 in NIH-3T3 Cells and Pig SSCs (Spermatogonial Stem Cells)


2.6. Discussion
3. Experimental Section
3.1. Animals and Samples
3.2. Total RNA Isolation and cDNA Synthesis
3.3. Cloning of Pig Stra8 Gene
3.4. 5'-RACE (Rapid Amplification of cDNA Ends) and 3'-RACE
3.5. Sequence Analyses
3.6. Semi-Quantitative RT-PCR
3.7. Quantitative Real-Time RT-PCR Assays
3.8. Construction of Expression Vectors
3.9. Cell Culture and Subcellular Localization
3.10. Statistical Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Borum, K. Oogenesis in the mouse: A study of the meiotic prophase. Exp. Cell. Res. 1961, 24, 495–507. [Google Scholar] [CrossRef]
- Hilscher, B.; Hilscher, W.; Bülthoff-Ohnolz, B.; Krämer, U.; Birke, A.; Pelzer, H.; Gauss, G. Kinetics of gametogenesis. Cell. Tissue Res. 1974, 154, 443–470. [Google Scholar]
- McLaren, A. Meiosis and differentiation of mouse germ cells. Symp. Soc. Exp. Biol. 1984, 38, 7–23. [Google Scholar]
- Speed, R.M. Meiosis in the fetal mouse ovary. Chromosoma 1982, 85, 427–437. [Google Scholar] [CrossRef]
- Koubova, J.; Menke, D.B.; Zhou, Q.; Capel, B.; Griswold, M.D.; Page, D.C. Retinoic acid regulates sex-specific timing of meiotic initiation in mice. Proc. Natl. Acad. Sci. USA 2006, 103, 2474–2479. [Google Scholar] [CrossRef]
- Bowles, J.; Knight, D.; Smith, C.; Wilhelm, D.; Richman, J.; Mamiya, S.; Yashiro, K.; Chawengsaksophak, K.; Wilson, M.J.; Rossant, J.; et al. Retinoid signaling determines germ cell fate in mice. Science 2006, 312, 596–600. [Google Scholar] [CrossRef]
- Li, H.; Palczewski, K.; Baehr, W.; Clagett-Dame, M. Vitamin A deficiency results in meiotic failure and accumulation of undifferentiated spermatogonia in prepubertal mouse testis. Biol. Reprod. 2011, 84, 336–341. [Google Scholar] [CrossRef]
- Clagett-Dame, M.; Knutson, D. Vitamin A in reproduction and development. Nutrients 2011, 3, 385–428. [Google Scholar] [CrossRef]
- Raverdeau, M.; Gely-Pernot, A.; Féret, B.; Dennefeld, C.; Benoit, G.; Davidson, I.; Chambon, P.; Mark, M.; Ghyselinck, N.B. Retinoic acid induces Sertoli cell paracrine signals for spermatogonia differentiation but cell autonomously drives spermatocyte meiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 16582–16587. [Google Scholar]
- Oulad-Abdelghani, M.; Bouillet, P.; Décimo, D.; Gansmuller, A.; Heyberger, S.; Dollé, P.; Bronner, S.; Lutz, Y.; Chambon, P. Characterization of a premeiotic germ cell-specific cytoplasmic protein encoded by Stra8, a novel retinoic acid-responsive gene. J. Cell. Biol. 1996, 135, 469–477. [Google Scholar] [CrossRef]
- Bouillet, P.; Oulad-Abdelghani, M.; Vicaire, S.; Garnier, J.M.; Schuhbaur, B.; Dollé, P.; Chambon, P. Efficient cloning of cDNAs of retinoic acid-responsive genes in P19 embryonal carcinoma cells and characterization of a novel mouse gene, Stra1 (mouse LERK-2/Eplg2). Dev. Biol. 1995, 170, 420–433. [Google Scholar] [CrossRef]
- Menke, D.B.; Koubova, J.; Page, D.C. Sexual differentiation of germ cells in XX mouse gonads occurs in an anterior-to-posterior wave. Dev. Biol. 2003, 262, 303–312. [Google Scholar] [CrossRef]
- Zhou, Q.; Li, Y.; Nie, R.; Friel, P.; Mitchell, D.; Evanoff, R.M.; Pouchnik, D.; Banasik, B.; McCarrey, J.R.; Small, C.; et al. Expression of stimulated by retinoic acid gene 8 (Stra8) and maturation of murine gonocytes and spermatogonia induced by retinoic acid in vitro. Biol. Reprod. 2008, 78, 537–545. [Google Scholar] [CrossRef]
- Silva, C.; Wood, J.R.; Salvador, L.; Zhang, Z.; Kostetskii, I.; Williams, C.J.; Strauss, J.F., 3rd. Expression profile of male germ cell-associated genes in mouse embryonic stem cell cultures treated with all-trans retinoic acid and testosterone. Mol. Reprod. Dev. 2009, 76, 11–21. [Google Scholar] [CrossRef]
- Ohta, K.; Lin, Y.; Hogg, N.; Yamamoto, M.; Yamazaki, Y. Direct effects of retinoic acid on entry of fetal male germ cells into meiosis in mice. Biol. Reprod. 2010, 83, 1056–1063. [Google Scholar] [CrossRef]
- Childs, A.J.; Cowan, G.; Kinnell, H.L.; Anderson, R.A.; Saunders, P.T. Retinoic Acid signalling and the control of meiotic entry in the human fetal gonad. PLoS One 2011, 6, e20249. [Google Scholar]
- Ge, C.T.; Zhou, W.X.; Wang, W.; Li, C.Y.; Gao, Y.L.; Qian, G.Y. Retinoic acid directly regulates the initiation and completion of meiosis in chicken primordial germ cells. Sci. Sin. Vitae 2011, 41, 1167–1176. [Google Scholar]
- Baltus, A.E.; Menke, D.B.; Hu, Y.C.; Goodheart, M.L.; Carpenter, A.E.; de Rooij, D.G.; Page, D.C. In germ cells of mouse embryonic ovaries, the decision to enter meiosis precedes premeiotic DNA replication. Nat. Genet. 2006, 38, 1430–1434. [Google Scholar] [CrossRef]
- Anderson, E.L.; Baltus, A.E.; Roepers-Gajadien, H.L.; Hassold, T.J.; de Rooij, D.G.; van Pelt, A.M.; Page, D.C. Stra8 and its inducer, retinoic acid, regulate meiotic initiation in both spermatogenesis and oogenesis in mice. Proc. Natl. Acad. Sci. USA 2008, 105, 14976–14980. [Google Scholar] [CrossRef]
- Feng, C.W.; Bowles, J.; Koopman, P. Control of mammalian germ cell entry into meiosis. Mol. Cell Endocrinol. 2014, 382, 488–497. [Google Scholar] [CrossRef]
- Snyder, E.M.; Small, C.; Griswold, M.D. Retinoic acid availability drives the asynchronous initiation of spermatogonial differentiation in the mouse. Biol. Reprod. 2010, 83, 783–790. [Google Scholar] [CrossRef]
- Lu, C.; Xu, M.; Wang, Y.; Qin, Y.; Du, G.; Wu, W.; Han, X.; Ji, C.; Yang, Y.; Gu, A.; et al. Genetic variants in meiotic program initiation pathway genes are associated with spermatogenic impairment in a Han Chinese population. PLoS One 2013, 8, e53443. [Google Scholar]
- Bonache, S.; Algaba, F.; Franco, E.; Bassas, L.; Larriba, S. Altered gene expression signature of early stages of the germ line supports the pre-meiotic origin of human spermatogenic failure. Andrology 2014, 2, 596–606. [Google Scholar] [CrossRef]
- Cheng, P.L. A highly prolific pig breed of China- the Taihu breed. Parts I and II. Pig News Inf. 1983, 4, 407–425. [Google Scholar]
- Lunstra, D.D.; Ford, J.J.; Klindt, J.; Wise, T.H. Physiology of the Meishan boar. J. Reprod. Fertil. Suppl. 1997, 52, 181–193. [Google Scholar]
- Miyamoto, T.; Sengoku, K.; Takuma, N.; Hasuike, S.; Hayashi, H.; Yamauchi, T.; Yamashita, T.; Ishikawa, M. Isolation and expression analysis of the testis-specific gene, Stra8, stimulated by retinoic acid gene 8. J. Assist. Reprod. Genet. 2002, 19, 531–535. [Google Scholar] [CrossRef]
- Liu, J.; Zheng, Q.; Deng, Y.; Cheng, C.S.; Kallenbach, N.R.; Lu, M. A seven-helix coiled coil. Proc. Natl. Acad. Sci. USA 2006, 103, 15457–15462. [Google Scholar]
- Murre, C.; Bain, G.; Dijk, M.A.V.; Engel, I.; Furnari, B.A.; Massari, M.E.; Matthews, J.R.; Quong, M.W.; Rivera, R.R.; Stuiver, M.H. Structure and function of helix-loop-helix proteins. Biochim. Biophys. Acta 1994, 1218, 129–135. [Google Scholar] [CrossRef]
- Zhou, Q.; Nie, R.; Li, Y.; Friel, P.; Mitchel, l.D.; Hess, R.A.; Small, C.; Griswold, M.D. Expression of stimulated by retinoic acid gene 8 (Stra8) in spermatogenic cells induced by retinoic acid: An in vivo study in vitamin A-sufficient postnatal murine testes. Biol. Reprod. 2008, 79, 35–42. [Google Scholar] [CrossRef]
- Xiongpin, J.; Jiatong, D. Research on maturity of testis and puberty of Erhualian boar. (in Chinese). J. Sichuan Agric. Univ. 1998, 2, 234–236. [Google Scholar]
- Biqin, F.; Jiafeng, H.; Duoxian, H.; Deshun, P.; Junliang, T.; Baoshen, Z. Research on reproductive characters of Erhualian Pig—Boar sex behavior and development of reproductive function. Anim. Husb. Vet. Med. 1980, 2, 4–8. [Google Scholar]
- Stouffs, K.; Vandermaelen, D.; Tournaye, H.; Liebaers, I.; van Steirteghem, A.; Lissens, W. Genetics and male infertility. Verh. K. Acad. Geneeskd. Belg. 2009, 71, 115–139. [Google Scholar]
- Choi, Y.J.; Yoon, J.W.; Pyo, C.W.; Kim, J.A.; Bae, S.H.; Park, S.S. A possible role of STRA8 as a transcriptional factor. Genes Genomics 2010, 32, 521–526. [Google Scholar] [CrossRef]
- Engel, K.B. The Regulation and Toxicology of Spermatogenesis. Ph.D. Thesis, Boston University, Boston, MA, USA, 2008. [Google Scholar]
- Untergrasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40. [Google Scholar] [CrossRef]
- Song, C.Y. Molecular Cloning of Several Pig Sperm Functional Genes and Characterization of Their mRNA Expression Profiles in the Genital Tracts and Ejaculated Sperm. Ph.D. Thesis, Yangzhou University, Yangzhou, China, 2010. [Google Scholar]
- Scotto-Lavino, E.; Du, G.; Frohman, M.A. 5' end cDNA amplification using classic RACE. Nat. Protoc. 2006, 1, 2555–2562. [Google Scholar]
- Scotto-Lavino, E.; Du, G.; Frohman, M.A. 3' end cDNA amplification using classic RACE. Nat. Protoc. 2006, 6, 2742–2745. [Google Scholar]
- Yachdav, G.; Kloppmann, E.; Kajan, L.; Hecht, M.; Goldberg, T.; Hamp, T.; Hönigschmid, P.; Schafferhans, A.; Roos, M.; Bernhofer, M.; et al. PredictProtein—an open resource for online prediction of protein structural and functional features. Nucleic Acids Res. 2014. [Google Scholar] [CrossRef]
- Lupas, A.; van Dyke, M.; Stock, J. Predicting coiled coils from protein sequences. Science 1991, 252, 1162–1164. [Google Scholar] [CrossRef]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2012, 40, 302–305. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Kuijk, E.W.; Colenbrander, B.; Roelen, B.A. The effects of growth factors on in vitro-cultured porcine testicular cells. Reproduction 2009, 138, 721–731. [Google Scholar] [CrossRef]
- Jia, Y.H. The Study of Isolation and Culture of Spermatcgonial Stem Cells of WZSP. Master’s Thesis, Liaoning Normal University, Dalian, China, 2009. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, X.; Chen, T.; Song, C.; Gao, B.; Zhang, Y. Cloning and Expression Characteristics of the Pig Stra8 Gene. Int. J. Mol. Sci. 2014, 15, 12480-12494. https://doi.org/10.3390/ijms150712480
Wang X, Chen T, Song C, Gao B, Zhang Y. Cloning and Expression Characteristics of the Pig Stra8 Gene. International Journal of Molecular Sciences. 2014; 15(7):12480-12494. https://doi.org/10.3390/ijms150712480
Chicago/Turabian StyleWang, Xiaoyan, Tingfeng Chen, Chengyi Song, Bo Gao, and Yani Zhang. 2014. "Cloning and Expression Characteristics of the Pig Stra8 Gene" International Journal of Molecular Sciences 15, no. 7: 12480-12494. https://doi.org/10.3390/ijms150712480