Chikusetsu Saponin V Attenuates MPP+-Induced Neurotoxicity in SH-SY5Y Cells via Regulation of Sirt1/Mn-SOD and GRP78/Caspase-12 Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
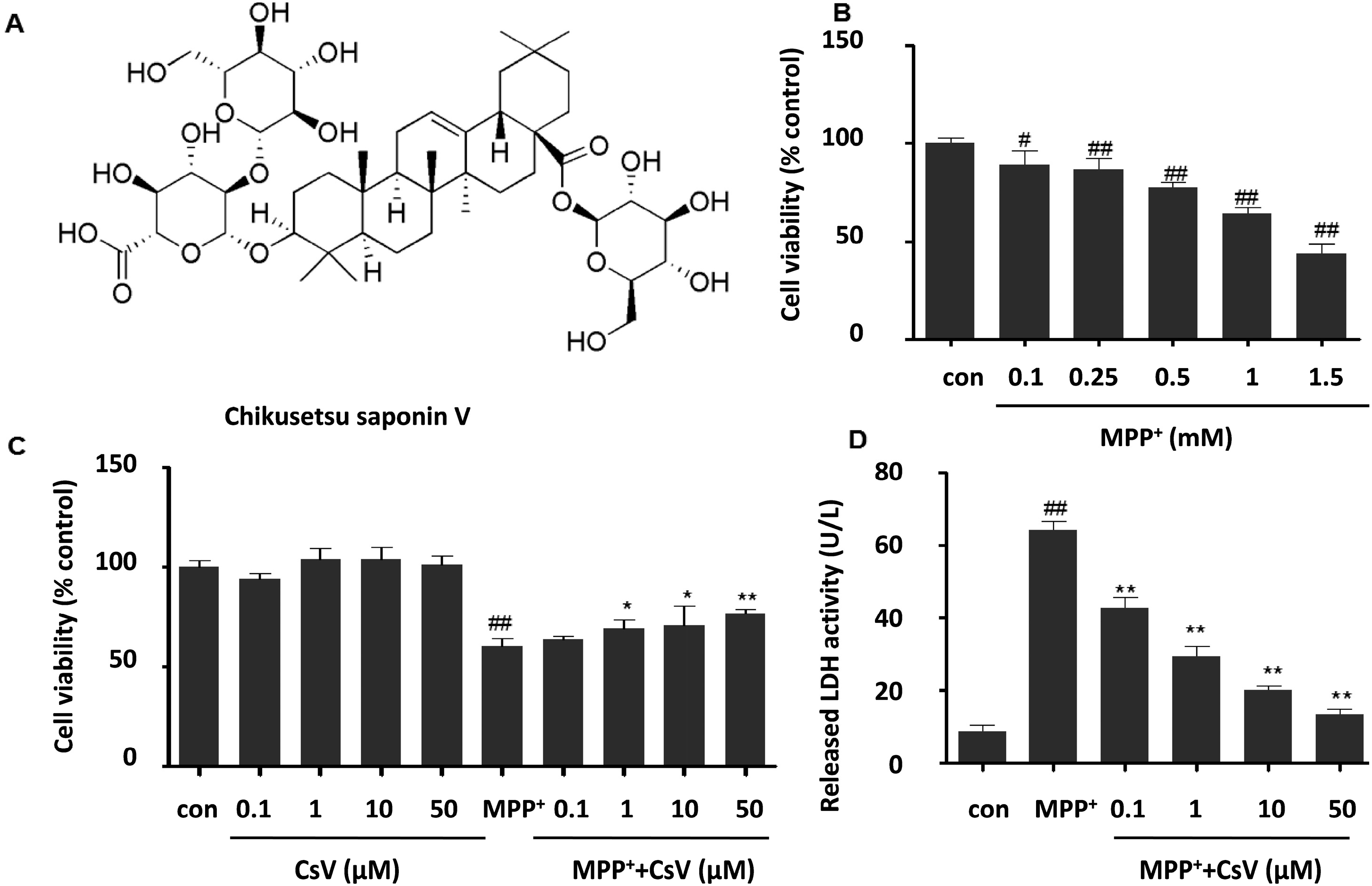
2.1. Chikusetsu Saponin V (CsV) Attenuated 1-Methyl-4-phenylpyridinium Ion (MPP+)-Induced Cytotoxicity in SH-SY5Y Cells

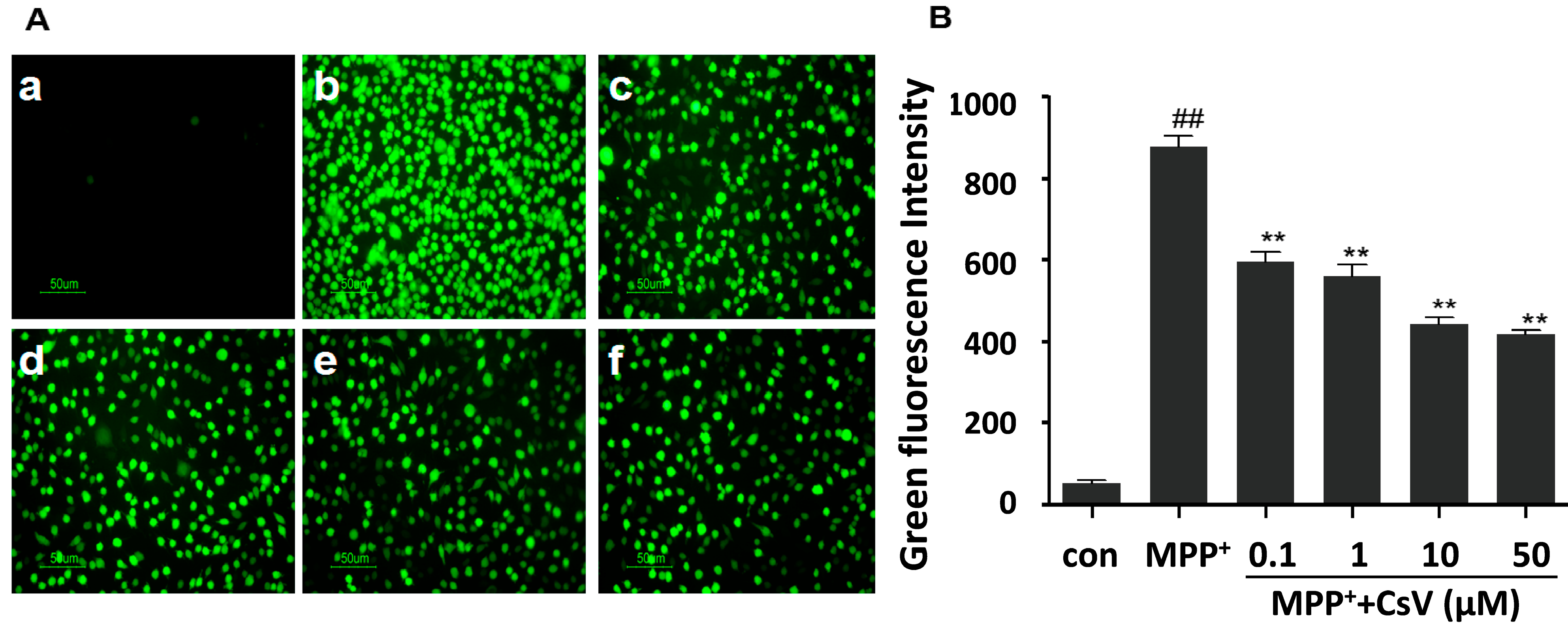
2.2. CsV Suppressed MPP+-Induced ROS Accumulation in SH-SY5Y Cells

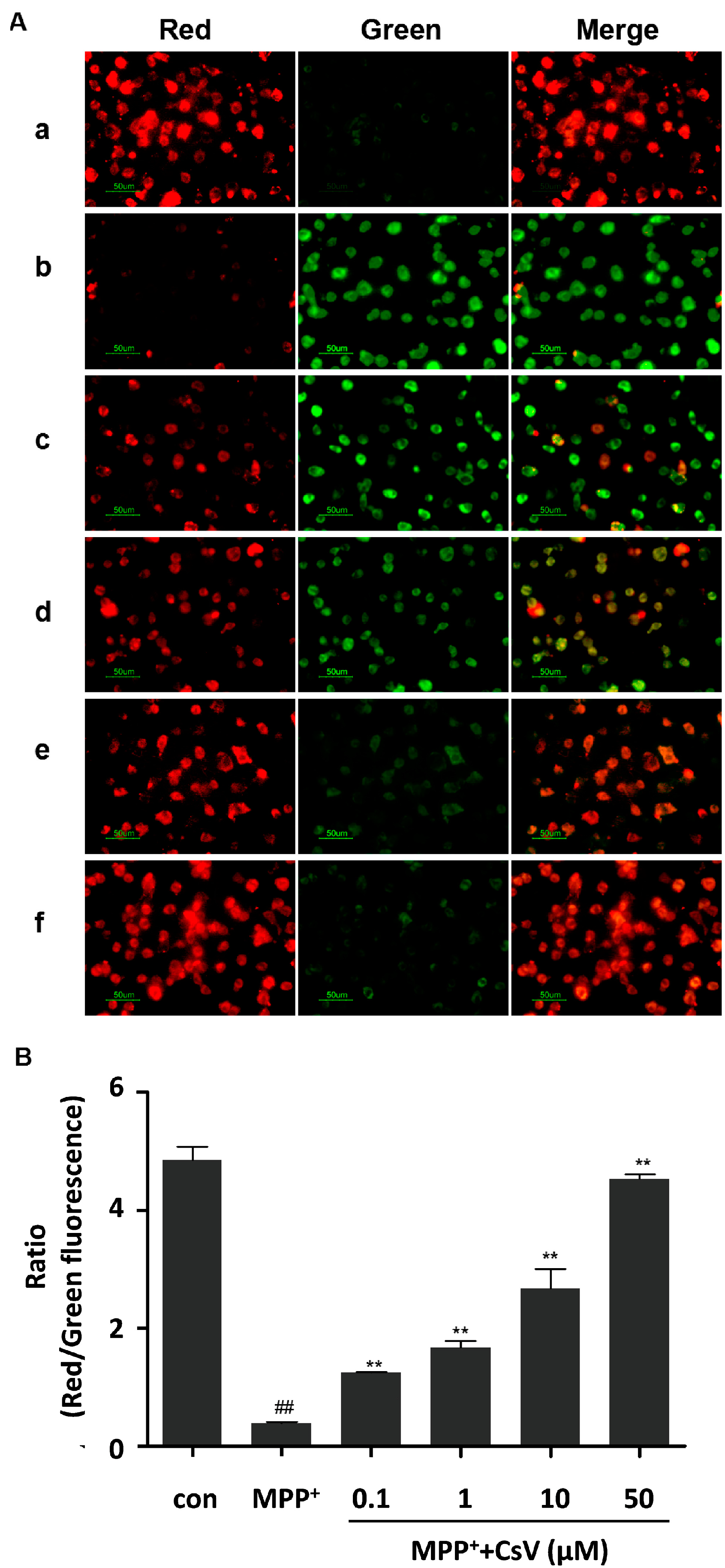
2.3. CsV Restored Mitochondrial Membrane Potential of SH-SY5Y Cells Treated with MPP+

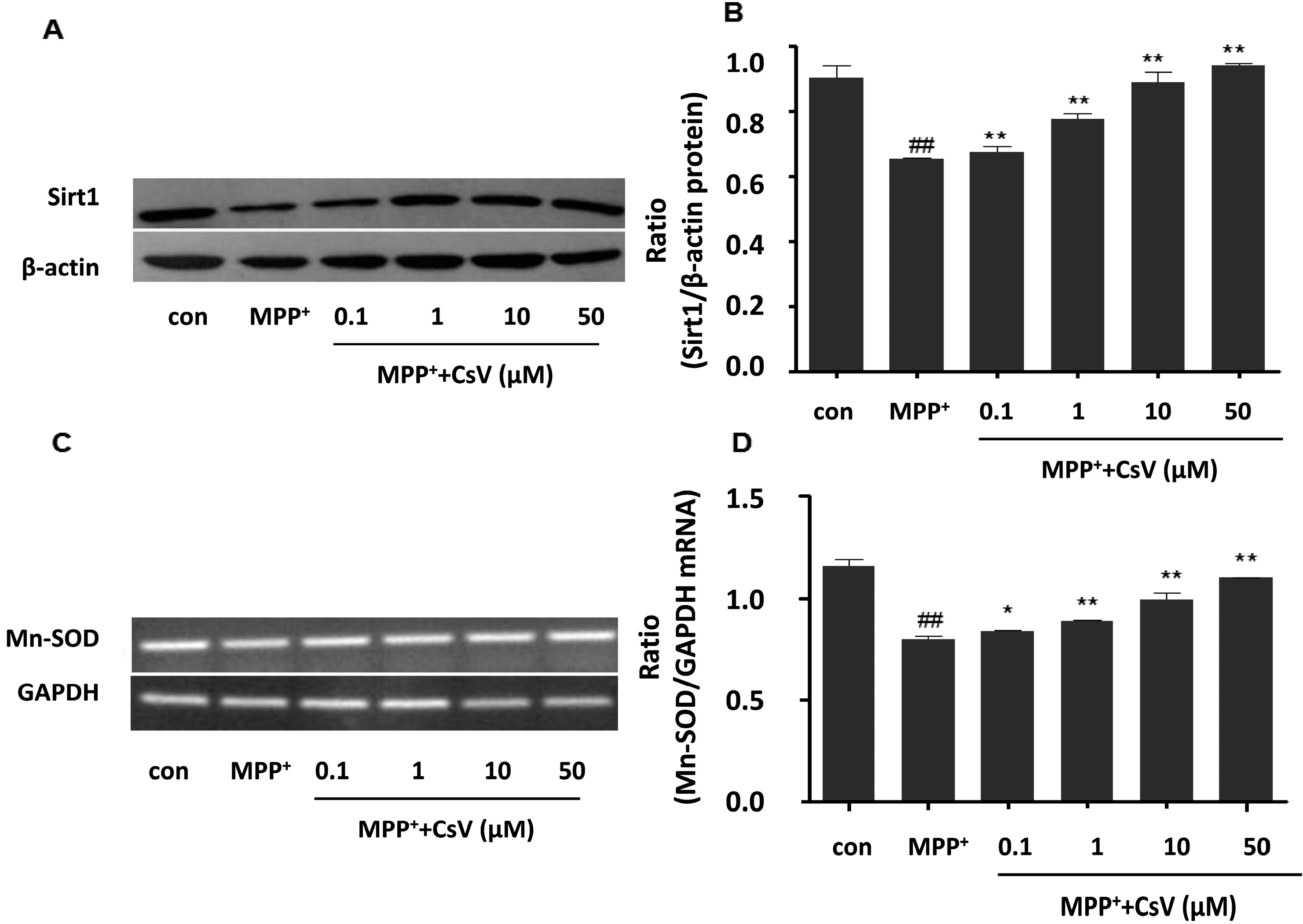
2.4. CsV Up-Regulated Sirt1 Protein and Mn-SOD mRNA Expressions in MPP+-Treated SH-SY5Y Cells

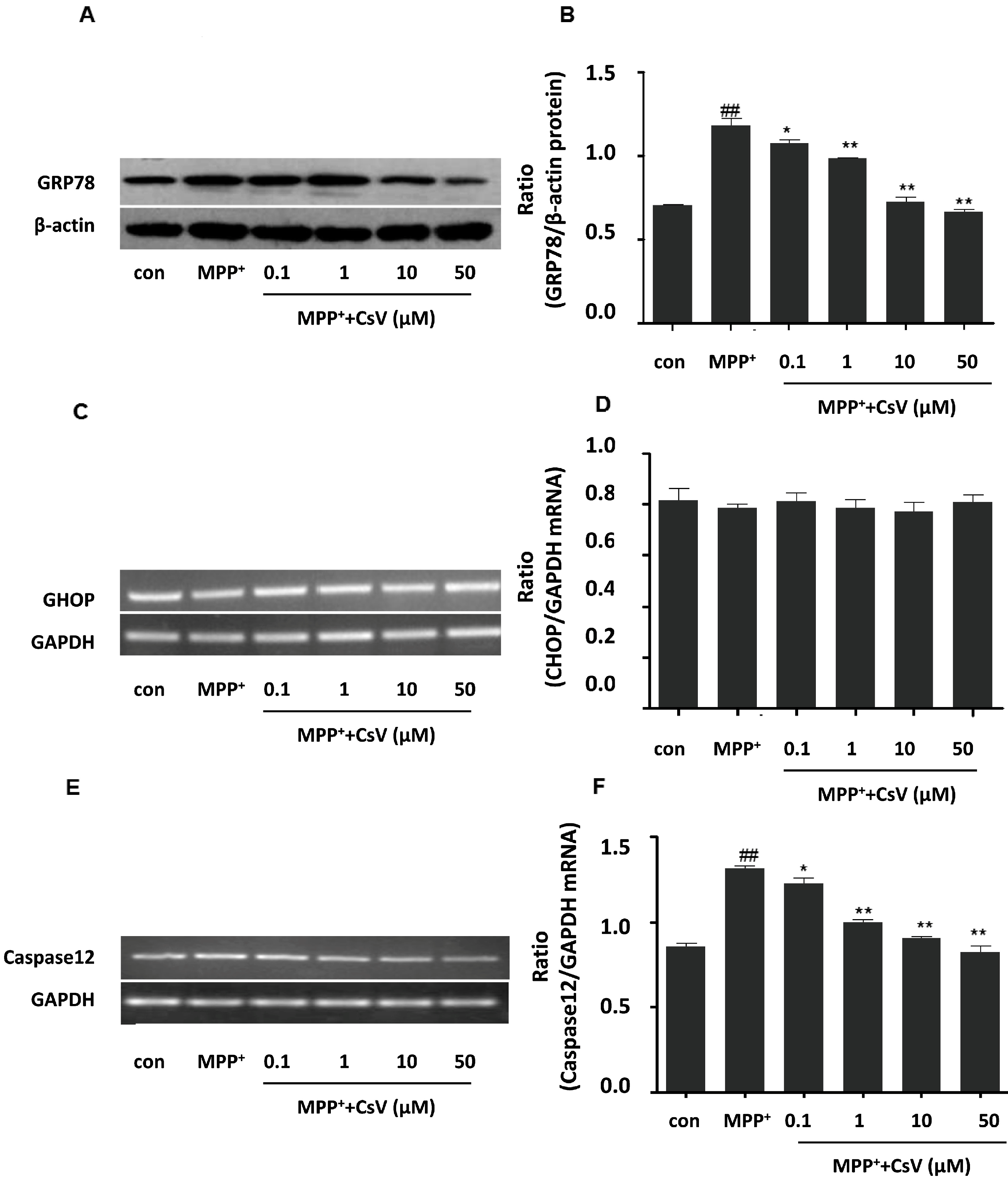
2.5. CsV Ameliorated MPP+-Induced Endoplasmic Reticulum (ER) Stress in SH-SY5Y Cells

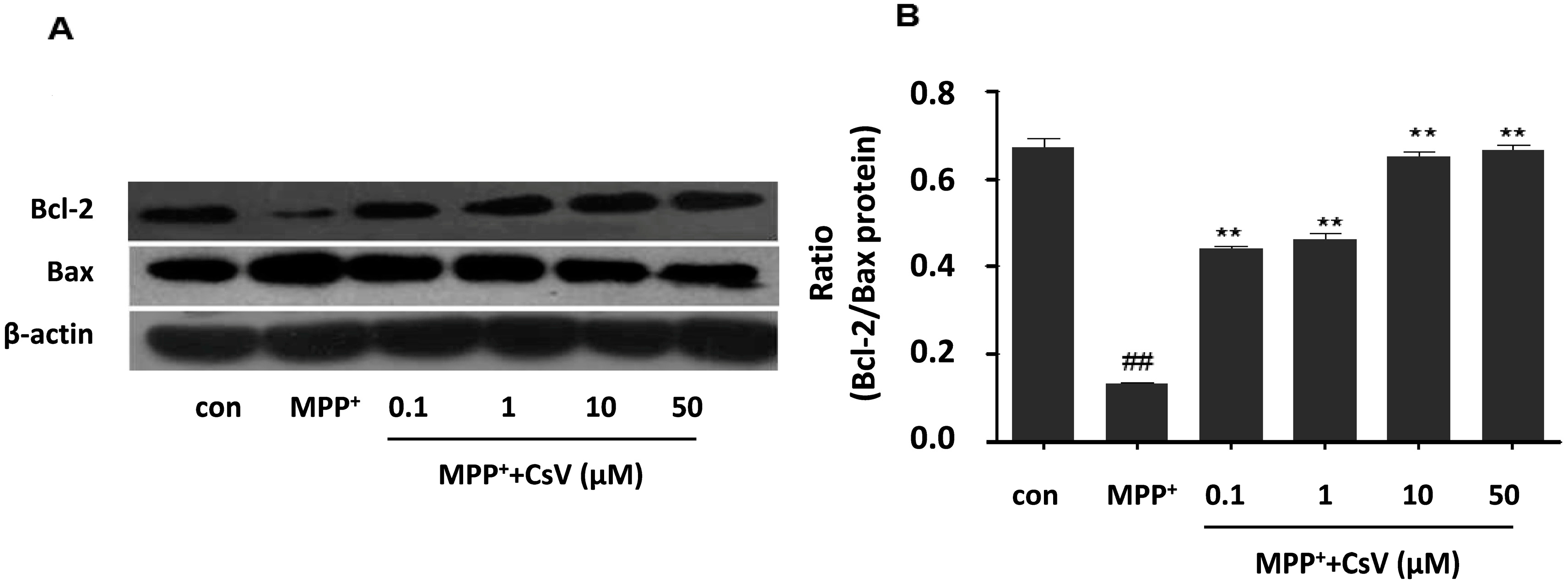
2.6. CsV Reversed MPP+-Induced Apoptosis in SH-SY5Y Cells

3. Discussion
4. Experimental Section
4.1. Materials
4.2. Cell Culture
4.3. MTT Viability Assay
4.4. Lactate Dehydrogenase (LDH) Release Assay
4.5. Detection of Intracellular ROS Level
4.6. Detection of Mitochondrial Membrane Potential
4.7. Western Blot
4.8. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Braak, H.; del Tredici, K.; Rub, U.; de Vos, R.A.; Jansen Steur, E.N.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Bloem, B.R.; Irwin, I.; Buruma, O.J.; Haan, J.; Roos, R.A.; Tetrud, J.W.; Langston, J.W. The MPTP model: Versatile contributions to the treatment of idiopathic Parkinson’s disease. J. Neurol. Sci. 1990, 97, 273–293. [Google Scholar] [CrossRef]
- Cleeter, M.W.; Cooper, J.M.; Schapira, A.H. Irreversible inhibition of mitochondrial complex I by 1-methyl-4-phenylpyridinium: Evidence for free radical involvement. J. Neurochem. 1992, 58, 786–789. [Google Scholar] [CrossRef]
- Hauser, D.N.; Hastings, T.G. Mitochondrial dysfunction and oxidative stress in Parkinson’s disease and monogenic parkinsonism. Neurobiol. Dis. 2013, 51, 35–42. [Google Scholar] [CrossRef]
- Moore, D.J.; West, A.B.; Dawson, V.L.; Dawson, T.M. Molecular pathophysiology of Parkinson’s disease. Annu. Rev. Neurosci. 2005, 28, 57–87. [Google Scholar]
- Corbi, G.; Conti, V.; Scapagnini, G.; Filippelli, A.; Ferrara, N. Role of sirtuins, calorie restriction and physical activity in aging. Front. Biosci. 2012, 4, 768–778. [Google Scholar]
- Wang, T.; Gu, J.; Wu, P.F.; Wang, F.; Xiong, Z.; Yang, Y.J.; Wu, W.N.; Dong, L.D.; Chen, J.G. Protection by tetrahydroxystilbene glucoside against cerebral ischemia: Involvement of JNK, Sirt1, and NF-êB pathways and inhibition of intracellular ROS/RNS generation. Free Radic. Biol. Med. 2009, 47, 229–240. [Google Scholar] [CrossRef]
- Brunet, A.; Sweeney, L.B.; Sturgill, J.F.; Chua, K.F.; Greer, P.L.; Lin, Y.; Tran, H.; Ross, S.E.; Mostoslavsky, R.; Cohen, H.Y. Stress-dependent regulation of FOXO transcription factors by the Sirt1 deacetylase. Science 2004, 303, 2011–2015. [Google Scholar] [CrossRef]
- Alcendor, R.R.; Gao, S.; Zhai, P.; Zablocki, D.; Holle, E.; Yu, X.; Tian, B.; Wagner, T.; Vatner, S.F.; Sadoshima, J. Sirt1 regulates aging and resistance to oxidative stress in the heart. Circ. Res. 2007, 100, 1512–1521. [Google Scholar] [CrossRef]
- Yun, J.M.; Chien, A.; Jialal, I.; Devaraj, S. Resveratrol up-regulates Sirt1 and inhibits cellular oxidative stress in the diabetic milieu: Mechanistic insights. J. Nutr. Biochem. 2012, 23, 699–705. [Google Scholar]
- Tang, B.L. Sirt1, neuronal cell survival and the insulin/IGF-1 aging paradox. Neurobiol. Aging 2006, 27, 501–505. [Google Scholar] [CrossRef]
- Yoon, H.; Kim, D.S.; Lee, G.H.; Kim, K.W.; Kim, H.R.; Chae, H.J. Apoptosis induced by manganese on neuronal sk-n-mc cell line: endoplasmic reticulum (er) stress and mitochondria dysfunction. Environ. Health Toxicol. 2011, 26, e2011017. [Google Scholar]
- Egawa, N.; Yamamoto, K.; Inoue, H.; Hikawa, R.; Nishi, K.; Mori, K.; Takahashi, R. The endoplasmic reticulum stress sensor, ATF6á, protects against neurotoxin-induced dopaminergic neuronal death. J. Biol. Chem. 2011, 286, 7947–7957. [Google Scholar]
- Hara, H.; Kamiya, T.; Adachi, T. Endoplasmic reticulum stress inducers provide protection against 6-hydroxydopamine-induced cytotoxicity. Neurochem. Int. 2011, 58, 35–43. [Google Scholar] [CrossRef]
- Hoozemans, J.J.; Scheper, W. Endoplasmic reticulum: The unfolded protein response is tangled in neurodegeneration. Int. J. Biochem. Cell Biol. 2012, 44, 1295–1298. [Google Scholar] [CrossRef]
- Chen, Z.; Peng, I.C.; Cui, X.; Li, Y.S.; Chien, S.; Shyy, J.Y. Shear stress, Sirt1, and vascular homeostasis. Proc. Natl. Acad. Sci. USA 2010, 107, 10268–10273. [Google Scholar]
- Fujita, E.; Kouroku, Y.; Jimbo, A.; Isoai, A.; Maruyama, K.; Momoi, T. Caspase-12 processing and fragment translocation into nuclei of tunicamycin-treated cells. Cell Death Differ. 2002, 9, 1108–1114. [Google Scholar] [CrossRef]
- Choi, A.Y.; Choi, J.H.; Yoon, H.; Hwang, K.Y.; Noh, M.H.; Choe, W.; Yoon, K.S.; Ha, J.; Yeo, E.J.; Kang, I. Luteolin induces apoptosis through endoplasmic reticulum stress and mitochondrial dysfunction in Neuro-2a mouse neuroblastoma cells. Eur. J. Pharmacol. 2011, 668, 115–126. [Google Scholar] [CrossRef]
- Han, L.K.; Zheng, Y.N.; Yoshikawa, M.; Okuda, H.; Kimura, Y. Anti-obesity effects of chikusetsusaponins isolated from Panax japonicus rhizomes. BMC Complement. Altern. Med. 2005, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Xu, J.; Xu, Y.; Zhang, C.; Wang, H.; He, Y.; Wang, T.; Yuan, D. Cardioprotective effects of saponins from Panax japonicus on acute myocardial ischemia against oxidative stress-triggered damage and cardiac cell death in rats. J. Ethnopharmacol. 2012, 140, 73–82. [Google Scholar] [CrossRef]
- Li, Y.G.; Ji, D.F.; Zhong, S.; Shi, L.G.; Hu, G.Y.; Chen, S. Saponins from Panax japonicus protect against alcohol-induced hepatic injury in mice by up-regulating the expression of GPX3, SOD1 and SOD3. Alcohol Alcohol. 2010, 45, 320–331. [Google Scholar] [CrossRef]
- Hosono-Nishiyama, K.; Matsumoto, T.; Kiyohara, H.; Nishizawa, A.; Atsumi, T.; Yamada, H. Suppression of Fas-mediated apoptosis of keratinocyte cells by chikusetsusaponins isolated from the roots of Panax japonicus. Planta Med. 2006, 72, 193–198. [Google Scholar] [CrossRef]
- Li, C.; Zhang, J.; Li, J.; Li, H.; Lin, Q.; Li, B. Research on chemical constituents and biological activities of rhizoma of Panax japonicus. Guid. J. Tradit. Chin. Med. Pharm. 2012, 4, 68–71. [Google Scholar]
- Zhang, Z.G.; Wu, L.; Wang, J.L.; Yang, J.D.; Zhang, J.; Zhang, J.; Li, L.H.; Xia, Y.; Yao, L.B.; Qin, H.Z.; et al. Astragaloside IV prevents MPP+-induced SH-SY5Y cell death via the inhibition of Bax-mediated pathways and ROS production. Mol. Cell. Biochem. 2012, 364, 209–216. [Google Scholar] [CrossRef]
- Kim, I.S.; Choi, D.K.; Jung, H.J. Neuroprotective effects of vanillyl alcohol in Gastrodia elata Blume through suppression of oxidative stress and anti-apoptotic activity in toxin-induced dopaminergic MN9D cells. Molecules 2011, 16, 5349–5361. [Google Scholar] [CrossRef]
- Wang, Y.H.; Yu, H.T.; Pu, X.P.; Du, G.H. Baicalein prevents 6-hydroxydopamine-induced mitochondrial dysfunction in SH-SY5Y cells via inhibition of mitochondrial oxidation and up-regulation of DJ-1 protein expression. Molecules 2013, 18, 14726–14738. [Google Scholar] [CrossRef]
- Yan, W.; Fang, Z.; Yang, Q.; Dong, H.; Lu, Y.; Lei, C.; Xiong, L. Sirt1 mediates hyperbaric oxygen preconditioning-induced ischemic tolerance in rat brain. J. Cereb. Blood Flow Metab. 2013, 33, 396–406. [Google Scholar] [CrossRef]
- He, W.; Wang, Y.; Zhang, M.Z.; You, L.; Davis, L.S.; Fan, H.; Yang, H.C.; Fogo, A.B.; Zent, R.; Harris, R.C.; et al. Sirt1 activation protects the mouse renal medulla from oxidative injury. J. Clin. Investig. 2010, 120, 1056–1068. [Google Scholar] [CrossRef]
- Xin, Y.F.; Wan, L.L.; Peng, J.L.; Guo, C. Alleviation of the acute doxorubicin-induced cardiotoxicity by Lycium barbarum polysaccharides through the suppression of oxidative stress. Food Chem. Toxicol. 2011, 49, 259–264. [Google Scholar] [CrossRef]
- Li, S.; Yan, T.; Yang, J.Q.; Oberley, T.D.; Oberley, L.W. The role of cellular glutathione peroxidase redox regulation in the suppression of tumor cell growth by manganese superoxide dismutase. Cancer Res. 2000, 60, 3927–3939. [Google Scholar]
- Chen, J.C.; Wu, M.L.; Huang, K.C.; Lin, W.W. HMG-CoA reductase inhibitors activate the unfolded protein response and induce cytoprotective GRP78 expression. Cardiovasc. Res. 2008, 80, 138–150. [Google Scholar] [CrossRef]
- Li, X.; Chen, W.; Zhang, L.; Liu, W.B.; Fei, Z. Inhibition of store-operated calcium entry attenuates MPP+-induced oxidative stress via preservation of mitochondrial function in PC12 cells: Involvement of Homer1a. PLoS One 2013, 8, e83638. [Google Scholar]
- Wang, Z.C.; Wang, J.F.; Li, Y.B.; Guo, C.X.; Liu, Y.; Fang, F.; Gong, S.L. Involvement of endoplasmic reticulum stress in apoptosis of testicular cells induced by low-dose radiation. J. Huazhong Univ. Sci. Technol. Med. Sci. 2013, 33, 551–558. (In Chinese) [Google Scholar] [CrossRef]
- Lakshmanan, A.P.; Thandavarayan, R.A.; Palaniyandi, S.S.; Sari, F.R.; Meilei, H.; Giridharan, V.V.; Soetikno, V.; Suzuki, K.; Kodama, M.; Watanabe, K. Modulation of AT-1R/CHOP-JNK-Caspase12 pathway by olmesartan treatment attenuates ER stress-induced renal apoptosis in streptozotocin-induced diabetic mice. Eur. J. Pharm. Sci. 2011, 44, 627–634. [Google Scholar]
- Jin, M.L.; Park, S.Y.; Kim, Y.H.; Oh, J.I.; Lee, S.J.; Park, G. The neuroprotective effects of cordycepin inhibit glutamate-induced oxidative and ER stress-associated apoptosis in hippocampal HT22 cells. Neurotoxicology 2014, 41, 102–111. [Google Scholar] [CrossRef]
- Henshall, D.C.; Engel, T. Contribution of apoptosis-associated signaling pathways to epileptogenesis: Lessons from Bcl-2 family knockouts. Front. Cell. Neurosci. 2013, 7, 110. [Google Scholar]
- Levy, O.A.; Malagelada, C.; Greene, L.A. Cell death pathways in Parkinson’s disease: Proximal triggers, distal effectors, and final steps. Apoptosis 2009, 14, 478–500. [Google Scholar] [CrossRef]
- Offen, D.; Beart, P.M.; Cheung, N.S.; Pascoe, C.J.; Hochman, A.; Gorodin, S.; Melamed, E.; Bernard, R.; Bernard, O. Transgenic mice expressing human Bcl-2 in their neurons are resistant to 6-hydroxydopamine and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine neurotoxicity. Proc. Natl. Acad. Sci. USA 1998, 95, 5789–5794. [Google Scholar] [CrossRef]
- Blum, D.; Torch, S.; Lambeng, N.; Nissou, M.; Benabid, A.L.; Sadoul, R.; Verna, J.M. Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamine and MPTP: Contribution to the apoptotic theory in Parkinson’s disease. Prog. Neurobiol. 2001, 65, 135–172. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yuan, D.; Wan, J.-Z.; Deng, L.-L.; Zhang, C.-C.; Dun, Y.-Y.; Dai, Y.-W.; Zhou, Z.-Y.; Liu, C.-Q.; Wang, T. Chikusetsu Saponin V Attenuates MPP+-Induced Neurotoxicity in SH-SY5Y Cells via Regulation of Sirt1/Mn-SOD and GRP78/Caspase-12 Pathways. Int. J. Mol. Sci. 2014, 15, 13209-13222. https://doi.org/10.3390/ijms150813209
Yuan D, Wan J-Z, Deng L-L, Zhang C-C, Dun Y-Y, Dai Y-W, Zhou Z-Y, Liu C-Q, Wang T. Chikusetsu Saponin V Attenuates MPP+-Induced Neurotoxicity in SH-SY5Y Cells via Regulation of Sirt1/Mn-SOD and GRP78/Caspase-12 Pathways. International Journal of Molecular Sciences. 2014; 15(8):13209-13222. https://doi.org/10.3390/ijms150813209
Chicago/Turabian StyleYuan, Ding, Jing-Zhi Wan, Li-Li Deng, Chang-Cheng Zhang, Yao-Yan Dun, Yan-Wen Dai, Zhi-Yong Zhou, Chao-Qi Liu, and Ting Wang. 2014. "Chikusetsu Saponin V Attenuates MPP+-Induced Neurotoxicity in SH-SY5Y Cells via Regulation of Sirt1/Mn-SOD and GRP78/Caspase-12 Pathways" International Journal of Molecular Sciences 15, no. 8: 13209-13222. https://doi.org/10.3390/ijms150813209