Effects of Lead on the Morphology and Structure of the Nucleolus in the Root Tip Meristematic Cells of Allium cepa L.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
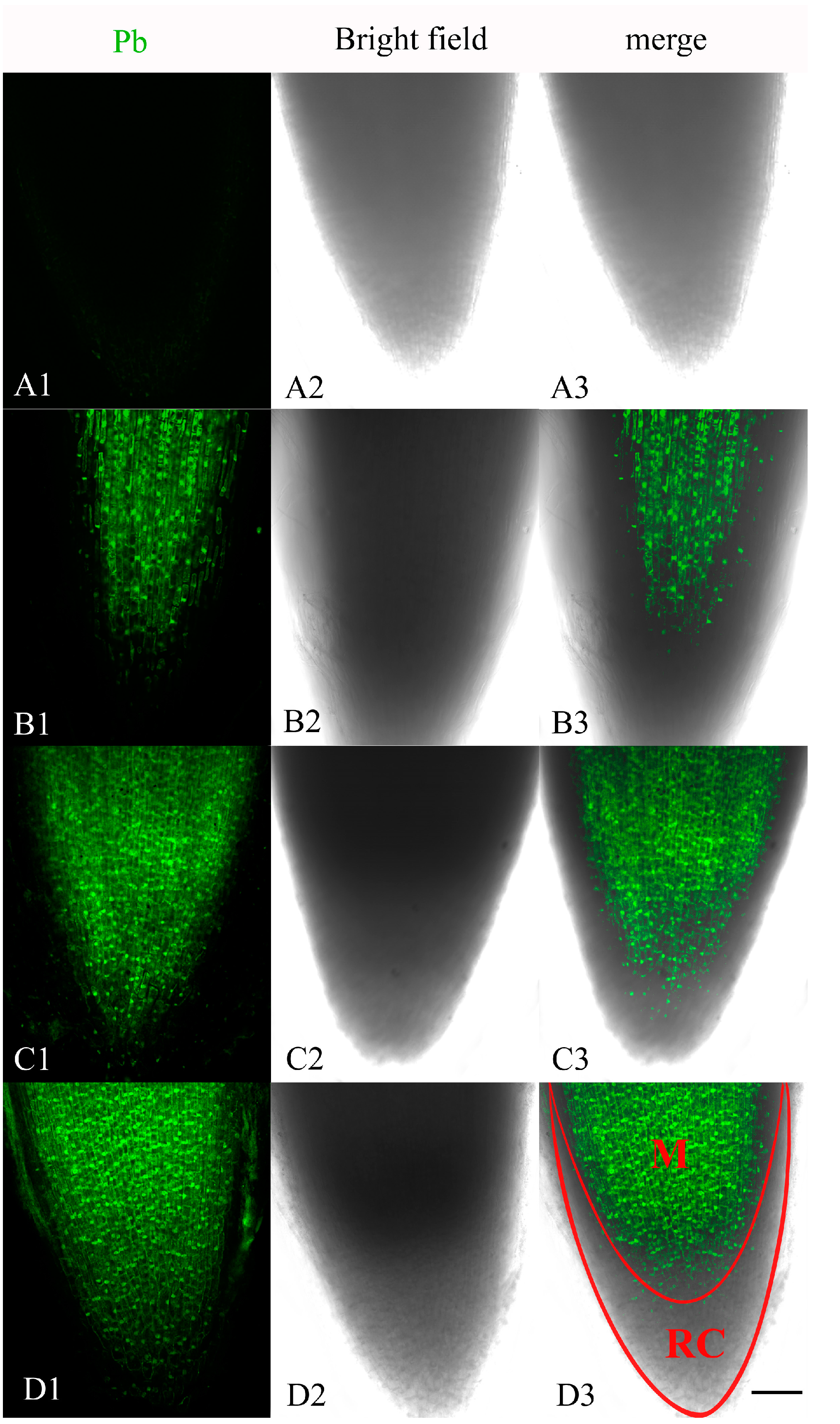
2.1. Localization of Pb in the Root Tip Meristematic Cells

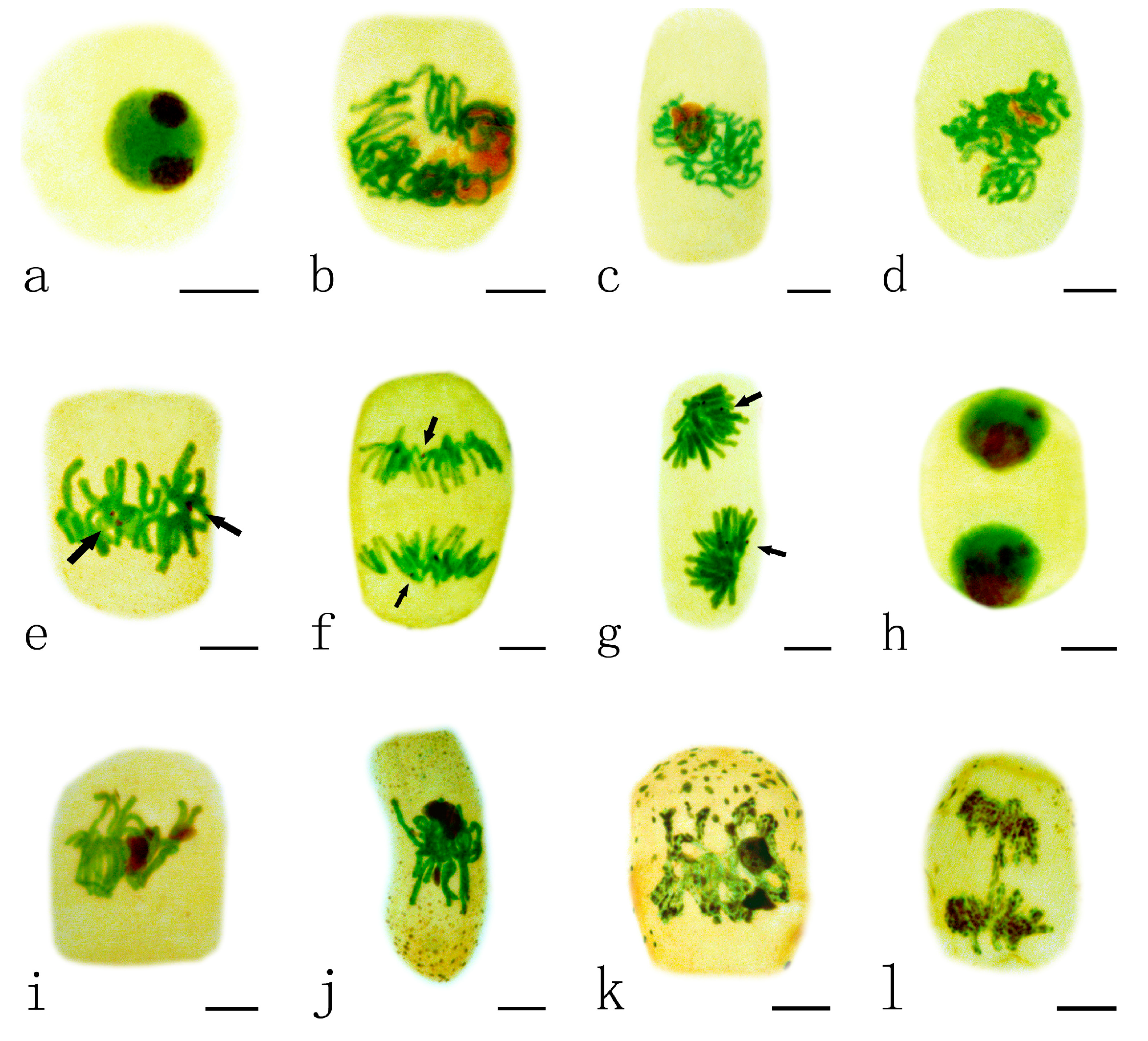
2.2. Effects of Pb on the Nucleolar Organization Regions

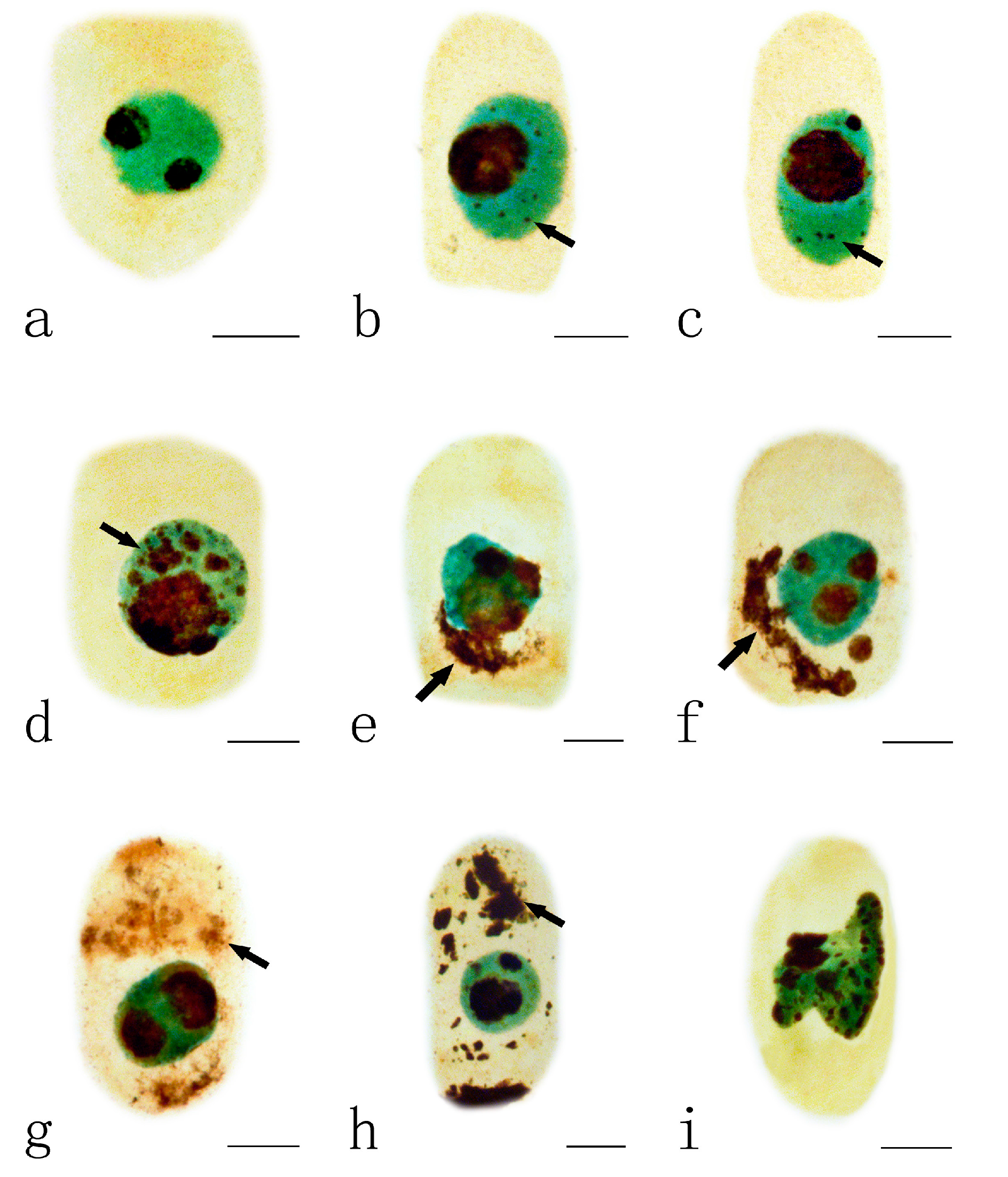
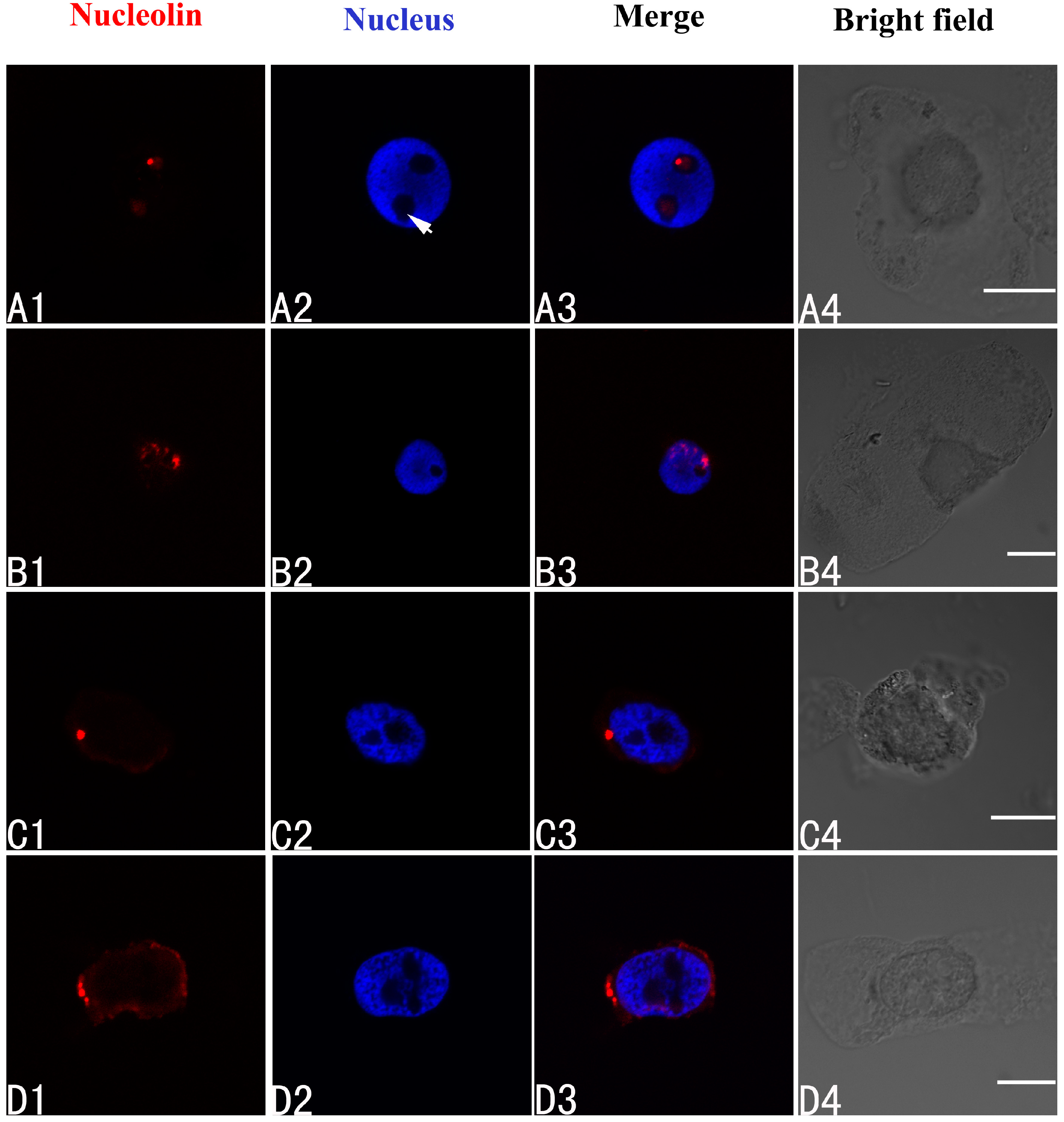
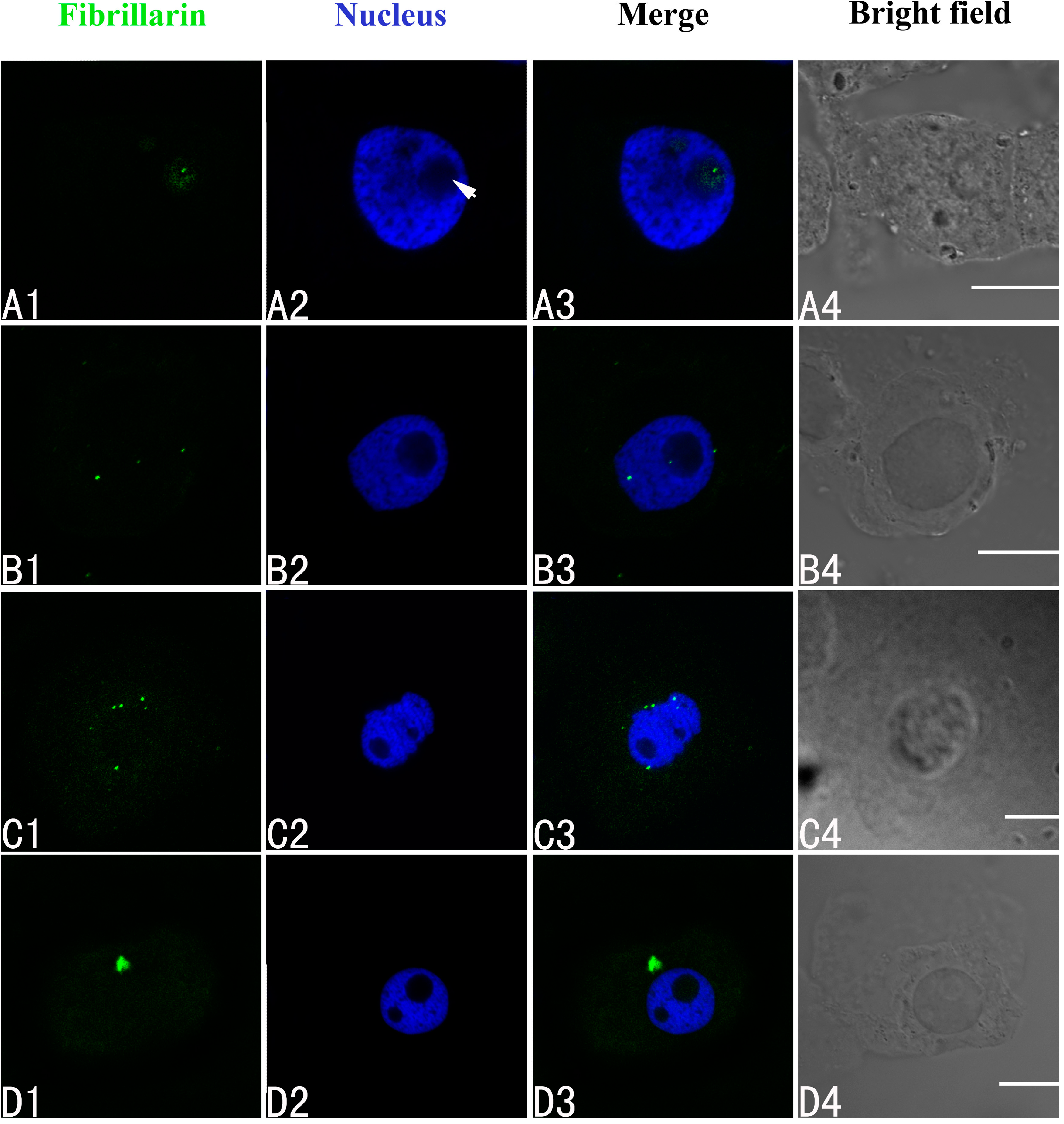
2.3. Effects of Pb on Nucleoli

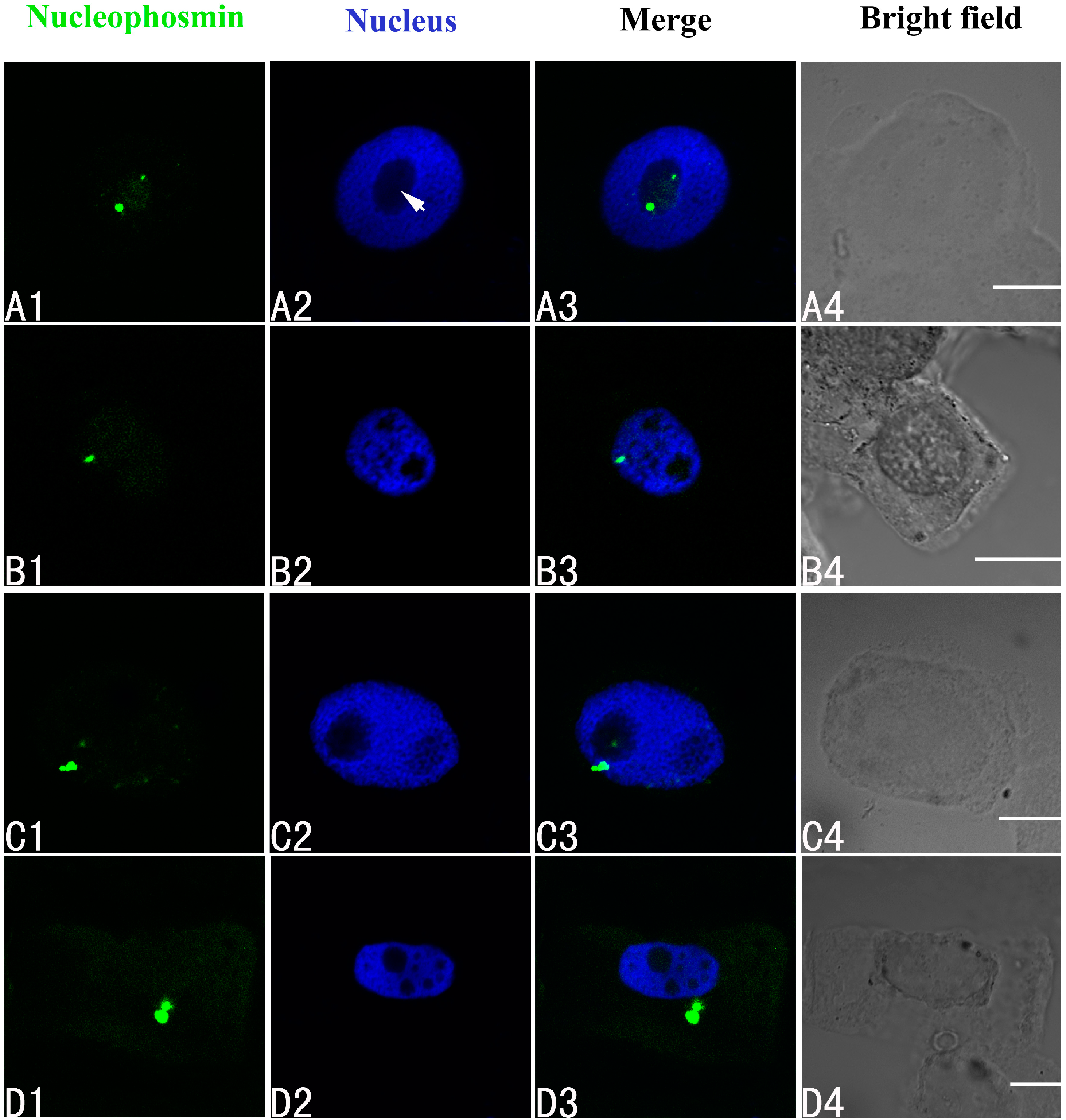
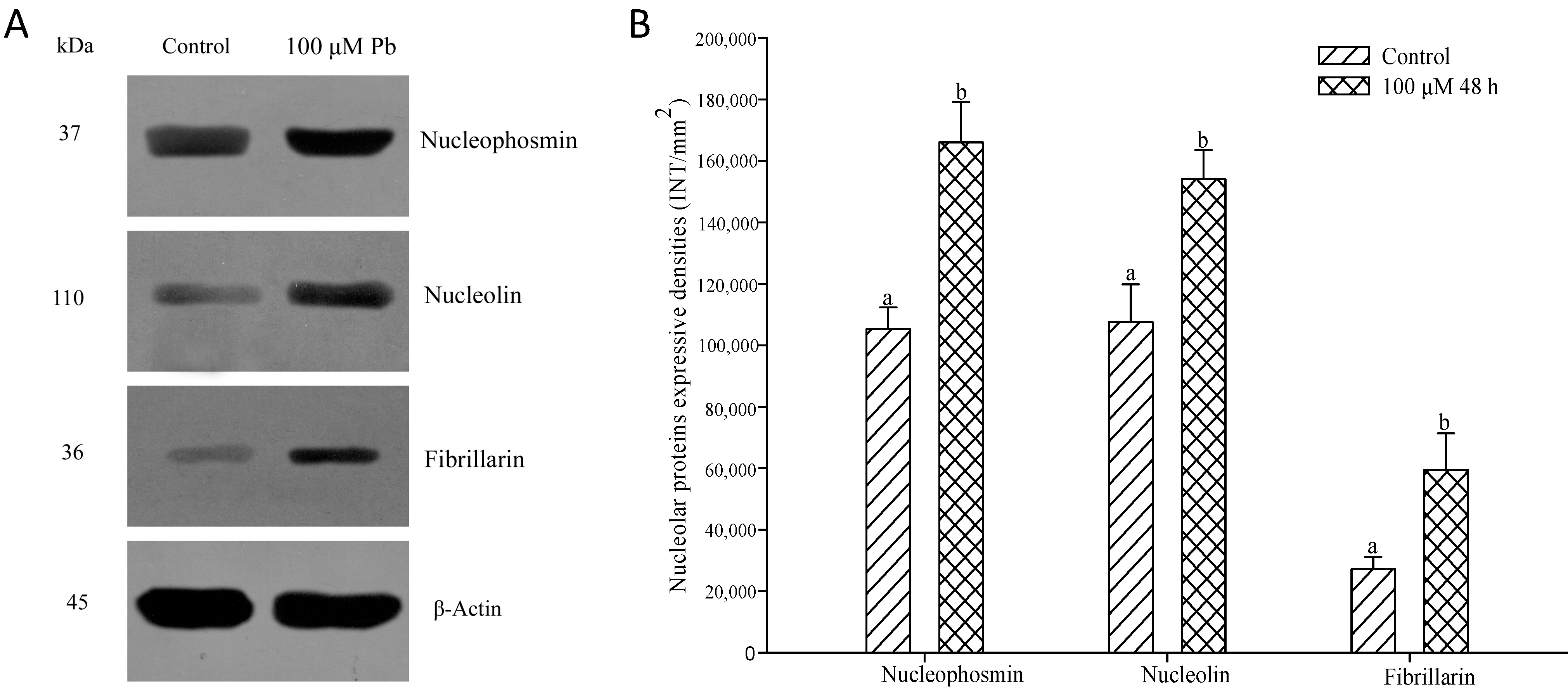
2.4. Effects of Pb on Three Major Nucleoproteins



2.5. Expression of Three Major Nucleoproteins in Relation to Pb Treatment

2.6. Discussion
3. Experimental Section
3.1. Culture Condition and Pb Treatment
3.2. Fluorescence Labeling
3.3. Silver-Staining
3.4. Immunofluorescence Staining
3.5. Western Blotting
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sengar, R.S.; Gautam, M.; Sengar, R.S.; Garg, S.K.; Sengar, K.; Chaudhary, R. Lead stress effects on physiobiochemical activities of higher plants. Rev. Environ. Contam. Toxicol. 2008, 196, 73–93. [Google Scholar]
- Waranusantigul, P.; Lee, H.; Kruatrachue, M.; Pokethitiyook, P.; Auesukaree, C. Isolation and characterization of lead-tolerant Ochrobactrum intermedium and its role in enhancing lead accumulation by Eucalyptus camaldulensis. Chemosphere 2011, 85, 584–590. [Google Scholar] [CrossRef]
- Zheng, L.J.; Liu, X.M.; Lutz-Meindl, U.; Peer, T. Effects of lead and EDTA-assisted lead on biomass, lead uptake and mineral nutrients in Lespedeza chinensis and Lespedeza davidii. Water AirSoil. Poll. 2011, 220, 57–68. [Google Scholar] [CrossRef]
- Levan, A. Cytological reaction induced by inorganic salt solutions. Nature 1945, 156. [Google Scholar] [CrossRef]
- Jiang, W.; Liu, D. Effects of Pb2+ on root growth, cell division, and nucleolus of Brassica juncea L. Isr. J. Plant Sci. 1999, 47, 153–156. [Google Scholar] [CrossRef]
- Glińska, S.; Bartczak, M.; Oleksiak, S.; Wolska, A.; Gabara, B.; Posmyk, M.; Janas, K. Effects of anthocyanin-rich extract from red cabbage leaves on meristematic cells of Allium cepa L. roots treated with heavy metals. Ecotoxicol. Environ. Saf. 2007, 68, 343–350. [Google Scholar] [CrossRef]
- Kozhevnikova, A.D.; Seregin, I.V.; Bystrova, E.I.; Belyaeva, A.I.; Kataeva, M.N.; Ivanov, V.B. The effects of lead, nickel, and strontium nitrates on cell division and elongation in maize root. Rus. J. Plant Physiol. 2009, 56, 242–250. [Google Scholar] [CrossRef]
- Liu, D.H.; Jiang, W.S.; Wang, W.; Zhao, F.M.; Lu, C. Effects of lead on root growth cell division and nucleolus of Allium cepa. Environ. Pollut. 1994, 86, 1–4. [Google Scholar] [CrossRef]
- Balcerzak, Ł.; Glińska, S.; Godlewski, M. The reaction of Lupinus angustifolius L. root meristematic cell nucleoli to lead. Protoplasma 2011, 248, 353–361. [Google Scholar] [CrossRef]
- Liu, D.; Islam, E.; Li, T.Q.; Yang, X.; Jin, X.F.; Mahmood, Q. Comparison of synthetic chelators and low molecular weight organic acids in enhancing phytoextraction of heavy metals by two ecotypes of Sedum alfredii Hance. J. Hazard. Mater. 2008, 153, 114–122. [Google Scholar] [CrossRef]
- Gupta, D.K.; Nicoloso, F.T.; Schetinger, M.R.C.; Rossato, L.V.; Huang, H.G.; Srivastava, S.; Yang, X.E. Lead induced responses of Pfaffia glomerata, an economically important Brazilian medicinal plant, under in vitro culture conditions. Bull. Environ. Contam. Toxicol. 2011, 86, 272–277. [Google Scholar] [CrossRef]
- Mroczek-Zdyrska, M.; Wójcik, M. The influence of selenium on root growth and oxidative stress induced by lead in Vicia faba L. minor plants. Biol. Trace Elem. Res. 2012, 147, 320–328. [Google Scholar] [CrossRef]
- Shu, X.; Yin, L.Y.; Zhang, Q.F.; Wang, W.B. Effect of Pb toxicity on leaf growth, antioxidant enzyme activities, and photosynthesis in cuttings and seedlings of Jatropha curcas L. Environ. Sci. Polut. Res. 2012, 19, 893–902. [Google Scholar] [CrossRef]
- Gichner, T.; Žnidar, I.; Száková, J. Evaluation of DNA damage and mutagenicity induced by lead in tobacco plants. Mutat. Res. 2008, 652, 186–190. [Google Scholar] [CrossRef]
- Liu, D.H.; Zou, J.; Meng, Q.M.; Zou, J.H.; Jiang, W.S. Uptake and accumulation and oxidative stress in garlic (Allium sativum L.) under lead phytotoxicity. Ecotoxicology 2009, 18, 134–143. [Google Scholar] [CrossRef]
- Olson, M.O.J.; Dundr, M. The moving parts of the nucleolus. Histochem. Cell Biol. 2005, 123, 203–216. [Google Scholar] [CrossRef]
- Gjerset, R.A. DNA damage, p14ARF, Nucleophosmin (NPM/B23), and cancer. J. Mol. Histol. 2006, 37, 239–251. [Google Scholar] [CrossRef]
- Sobol, M.; Gonzalez-Camacho, F.; Rodríguez-Vilariño, V.; Kordyum, E.; Medina, F.J. Subnucleolar location of fibrillarin and NopA64 in Lepidium sativum root meristematic cells is changed in altered gravity. Protoplasma 2006, 228, 209–219. [Google Scholar] [CrossRef]
- Liu, D.H.; Jiang, W.S. Effects of Al3+ on the nucleolus in root tip cells of Allium cepa. Hereditas 1991, 115, 213–219. [Google Scholar]
- Zhang, H.M.; Zhang, S.S.; Meng, Q.M.; Zou, J.; Jiang, W.S.; Liu, D.H. Effects of aluminum on nucleoli in root tip cells, root growth and the antioxidant defense system in Vicia faba L. Acta Biol. Crac. Bot. 2009, 51, 99–106. [Google Scholar]
- Liu, D.H.; Jiang, W.S.; Wang, W.; Zhai, L. Evaluation of metal ion toxicity on root tip cells by the Allium test. Isr. J. Plant Sci. 1995, 43, 125–133. [Google Scholar] [CrossRef]
- Lu, L.L.; Tian, S.K.; Yang, X.E.; Wang, X.C.; Brown, P.; Li, T.Q.; He, Z.L. Enhanced root-to-shoot translocation of cadmium in the hyperaccumulating ecotype of Sedum alfredii. J. Exp. Bot. 2008, 59, 3203–3213. [Google Scholar] [CrossRef]
- Li, L.Z.; Liu, X.L.; Peijnenburg, W.J.G.M.; Zhao, J.M.; Chen, X.B.; Yu, J.B.; Wu, H.F. Pathways of cadmium fluxes in the root of the halophyte Suaeda salsa. Ecotoxicol. Environ. Saf. 2012, 75, 1–7. [Google Scholar] [CrossRef]
- Zhao, F.Y.; Hu, F.; Zhang, S.Y.; Wang, K.; Zhang, C.R.; Liu, T. MAPKs regulate root growth by influencing auxin signaling and cell cycle-related gene expression in cadmium-stressed rice. Environ. Sci. Pollut. Res. 2013, 20, 5449–5460. [Google Scholar] [CrossRef]
- Jiang, W.S.; Liu, D.H. Pb-induced cellular defense system in the root meristematic cells of Allium sativum L. BMC Plant Biol. 2010, 10. [Google Scholar] [CrossRef]
- Leonova, O.G.; Karajan, B.P.; Ivlev, Y.F.; Ivanova, J.L.; Skarlato, S.O.; Popenko, V.I. Quantitative analysis of nucleolar chromatin distribution in the complex convoluted nucleoli of Didinium nasutum (Ciliophora). Biol. Res. 2013, 46, 69–74. [Google Scholar] [CrossRef]
- Boisvert, F.M.; van Koningsbruggen, S.; Navascués, J.; Lamond, A.I. The multifunctional nucleolus. Nat. Rev. Mol. Cell Biol. 2007, 8, 574–585. [Google Scholar] [CrossRef]
- Lempiainen, H.; Shore, D. Growth control and ribosome biogenesis. Curr. Opin. Cell Biol. 2009, 21, 855–863. [Google Scholar] [CrossRef]
- Trerè, D. AgNOR staining and quantification. Micron 2000, 2, 127–131. [Google Scholar] [CrossRef]
- Sato, S. Incorporation of argyrophilic proteins into the nucleolus-associated chromatin caused by exposure to actinomycin D. Biol. Cell 1988, 63, 19–26. [Google Scholar] [CrossRef]
- Raska, I.; Shaw, P.J.; Cmarko, D. New insights into nucleolar architecture and activity. Int. Rev. Cytol. 2006, 255, 177–135. [Google Scholar] [CrossRef]
- Qin, R.; Jiao, Y.Q.; Zhang, S.S.; Jiang, W.S.; Liu, D.H. Effects of aluminum on nucleoli in root tip cells and selected physiological and biochemical characters in Allium cepa var. agrogarum L. BMC Plant Biol. 2010, 10. [Google Scholar] [CrossRef]
- Zou, J.H.; Yue, J.Y.; Jiang, W.S.; Liu, D.H. Effects of cadmium stress on root tip cells and some physiological indexes in Allium cepa var. agrogarum L. Acta Biol. Crac. Bot. 2012, 54, 129–141. [Google Scholar]
- Sheldon, S.; Speers, W.C.; Lenhman, J.M. Nucleolar persistence in embryonal carcinoma cells. Exp. Cell Res. 1981, 132, 185–192. [Google Scholar] [CrossRef]
- Vostrikova, T.V.; Butorina, A.K. Cytogenetic responses of birch to stress factors. Biol. Bull. 2006, 33, 185–190. [Google Scholar] [CrossRef]
- Imazawa, T.; Nishikawa, A.; Tada, M.; Takahashi, M.; Hayashi, Y. Nucleolar segregation as an early marker for DNA damage; an experimental study in rats treated with 4-hydroxyaminoquinoline l-oxide. Virchows Arch. 1995, 426, 295–300. [Google Scholar]
- Chai, Z.F.; Zhu, H.M. Microelement; Peoples’ Health Publishing House: Beijing, China, 1983; pp. 283–298. (In Chinese) [Google Scholar]
- Goyer, R.A. Toxic and essential metal interactions. Annu. Rev. Nutr. 1997, 17, 37–50. [Google Scholar] [CrossRef]
- Goering, P.L. Lead-protein interactions as a basis for lead toxicity. Neurotoxicology 1993, 14, 45–60. [Google Scholar]
- Ouyang, H.; Vogel, H.J. Metal ion binding to calmodulin: NMR and fluorescence studies. Biometals 1998, 11, 213–222. [Google Scholar] [CrossRef]
- Xu, Y.H. Structure and function of calmodulin (I). Biochem. Biophys. 1985, 1, 9–13. [Google Scholar]
- Li, J.X.; Sun, D.Y. A study on CaM distribution in cells of living things. Chin. J. Cell Biol. 1991, 12, 1–6. [Google Scholar]
- Chen, Z.H.; Bai, J.; Shi, D.W.; Sun, D.Y. Production of antibody against wheat native calmodulin and its application to enzyme labeled immunohisto-chemistry. Acta Agric. BorealSin. 1991, 6, 76–80. (In Chinese) [Google Scholar]
- Andersen, J.S.; Lam, Y.W.; Leung, A.K.L.; Ong, S.E.; Lyon, C.E.; Lamond, A.I.; Mann, M. Nucleolar proteome dynamics. Nature 2005, 433, 77–83. [Google Scholar] [CrossRef]
- Hsieh, S.Y.; Hsu, C.Y.; He, J.R.; Liu, C.L.; Lo, S.J.; Chen, Y.C. Identifying apoptosis-evasion proteins/pathways in human hepatoma cells via induction of cellular hormesis by UV irradiation. J. Proteome Res. 2009, 8, 3977–3986. [Google Scholar] [CrossRef]
- Amin, M.A.; Matsunaga, S.; Ma, N.; Takata, H.; Yokoyama, M.; Uchiyama, S.; Fukui, K. Fibrillarin, a nucleolar protein, is required for normal nuclear morphology and cellular growth in HeLa cells. Biochem. Biophys. Res. Commun. 2007, 360, 320–326. [Google Scholar] [CrossRef]
- Hingorani, K.; Szebeni, A.; Olson, M.O. Mapping the functional domains of nucleolar protein B23. J. Biol. Chem. 2000, 275, 24451–24457. [Google Scholar] [CrossRef]
- Chiarella, S.; de, Cola. A.; Scaglione, G.L.; Carletti, E.; Graziano, V.; Barcaroli, D.; Lo Sterzo, C.; di Matteo, A.; di llio, C.; Falini, B.; et al. Nucleophosmin mutations alter its nucleolar localization by impairing G-quadruplex binding at ribosomal DNA. In Nucleic Acids Res.; 2013; Volume 41, pp. 3228–3239. [Google Scholar]
- Chathoth, K.T.; Ganesan, G.; Rao, M.R.S. Identification of a novel nucleolin related protein (NRP) gene expressed during rat spermatogenesis. BMC Mol. Biol. 2009, 10. [Google Scholar] [CrossRef]
- Sripinyowanich, S.; Chamnanmanoontham, N.; Udomchalothorn, T.; Maneeprasopsuk, S.; Santawee, P.; Buaboocha, T.; Qu, L.J.; Gu, H.Y.; Chadchawan, S. Overexpression of a partial fragment of the salt-responsive gene OsNUC1 enhances salt adaptation in transgenic Arabidopsis thaliana and rice (Oryza sativa L.) during salt stress. Plant Sci 2013, 213, 67–78. [Google Scholar] [CrossRef]
- Derenzini, M. The AgNORs. Micron 2000, 31, 117–120. [Google Scholar] [CrossRef]
- Sushil, L.M.D. AgNOR expression in central nervous system tumours. J. Med. Biol. Sci. 2011, 4, 1–9. [Google Scholar]
- Strauss, P.R.; Wilson, S.H. The Eukaryotic Nucleus: Molecular Biochemistry and Macromolecular Assemblies; The Telford Press: Caldwell, NJ, USA, 1990; pp. 783–811. [Google Scholar]
- Qin, R.; Jiang, W.S.; Liu, D.H. Cadmium can induce alterations in the cellular localization and expression of three major nucleolar proteins in root tip cells of Vicia faba L. Plant Soil 2013, 368, 365–373. [Google Scholar] [CrossRef]
- Qin, R.; Jiang, W.S.; Liu, D.H. Aluminum can induce alterations in the cellular localization and expression of three major nucleolar proteins in root tip cells of Allium cepa var. agrogarum L. Chemosphere 2013, 90, 827–834. [Google Scholar] [CrossRef]
- Dundr, M. Nuclear bodies: Multifunctional companions of the genome. Curr. Opin. Cell Biol. 2012, 24, 415–422. [Google Scholar] [CrossRef]
- Makimoto, Y.; Yano, H.; Kaneta, T.; Sato, Y.; Sato, S. Molecular cloning and gene expression of a fibrillarin homolog of tobacco BY-2 cells. Protoplasma 2006, 229, 53–62. [Google Scholar] [CrossRef]
- Chen, M.; von Mikecz, A. Specific inhibition of rRNA transcription and dynamic relocation of fibrillarin induced by mercury. Exp. Cell Res. 2000, 259, 225–238. [Google Scholar] [CrossRef]
- Rubbi, C.P.; Milner, J. Non-activated p53 co-localizes with sites of transcription within both the nucleoplasm and the nucleolus. Oncogene 2000, 19, 85–96. [Google Scholar] [CrossRef]
- Bunney, T.D.; Watkins, P.A.; Beven, A.F.; Shaw, P.J.; Hernandez, L.E.; Lomonossoff, G.P. Association of phosphatidylinositol 3-kinase with nuclear transcription sites in higher plants. Plant Cell 2000, 12, 1679–1688. [Google Scholar] [CrossRef]
- Ni, X.; Zhang, F. PI3K is involved in nucleolar structure and function on root-tip meristematic cells of Triticum aestivum L. Acta Histochem. 2014, 116, 838–843. [Google Scholar] [CrossRef]
- Dundr, M.; Leno, G.H.; Hammarskjöld, M.L.; Rekosh, D.; Helga-Maria, C.; Olson, M.O. The roles of nucleolar structure and function in the subcellular location of the HIV-1 Rev protein. J. Cell Sci. 1995, 108, 2811–2823. [Google Scholar]
- Fiskesjö, G. Nucleolar dissolution induced by aluminium in root cells of Allium. Physiol. Plant 1983, 59, 508–511. [Google Scholar] [CrossRef]
- Liu, D.; Jiang, W.; Guo, L.; Yuqing, H.; Cheng, L.; Fengmei, Z. Effects of nickel sulfate on root growth and nucleoli in root tip cells of Allium cepa. Isr. J. Plant Sci. 1994, 42, 143–148. [Google Scholar] [CrossRef]
- Liu, D.; Jiang, W.; Gao, X. Effects of cadmium on root growth, cell division and nucleoli in root tip cells of garlic. Biol. Plant 2003, 47, 79–83. [Google Scholar]
- Jiang, W.; Liu, D. Effects of Pb2+ on root growth, cell division, and nucleolus of Zea mays L. Bull. Environ. Contam. Toxicol. 2000, 65, 786–793. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jiang, Z.; Zhang, H.; Qin, R.; Zou, J.; Wang, J.; Shi, Q.; Jiang, W.; Liu, D. Effects of Lead on the Morphology and Structure of the Nucleolus in the Root Tip Meristematic Cells of Allium cepa L. Int. J. Mol. Sci. 2014, 15, 13406-13423. https://doi.org/10.3390/ijms150813406
Jiang Z, Zhang H, Qin R, Zou J, Wang J, Shi Q, Jiang W, Liu D. Effects of Lead on the Morphology and Structure of the Nucleolus in the Root Tip Meristematic Cells of Allium cepa L. International Journal of Molecular Sciences. 2014; 15(8):13406-13423. https://doi.org/10.3390/ijms150813406
Chicago/Turabian StyleJiang, Ze, Huaning Zhang, Rong Qin, Jinhua Zou, Junran Wang, Qiuyue Shi, Wusheng Jiang, and Donghua Liu. 2014. "Effects of Lead on the Morphology and Structure of the Nucleolus in the Root Tip Meristematic Cells of Allium cepa L." International Journal of Molecular Sciences 15, no. 8: 13406-13423. https://doi.org/10.3390/ijms150813406