Population Structure of the Greenhouse Whitefly, Trialeurodes vaporariorum (Westwood), an Invasive Species from the Americas, 60 Years after Invading China

Abstract
:1. Introduction
2. Results and Discussion
2.1. Genetic Diversity
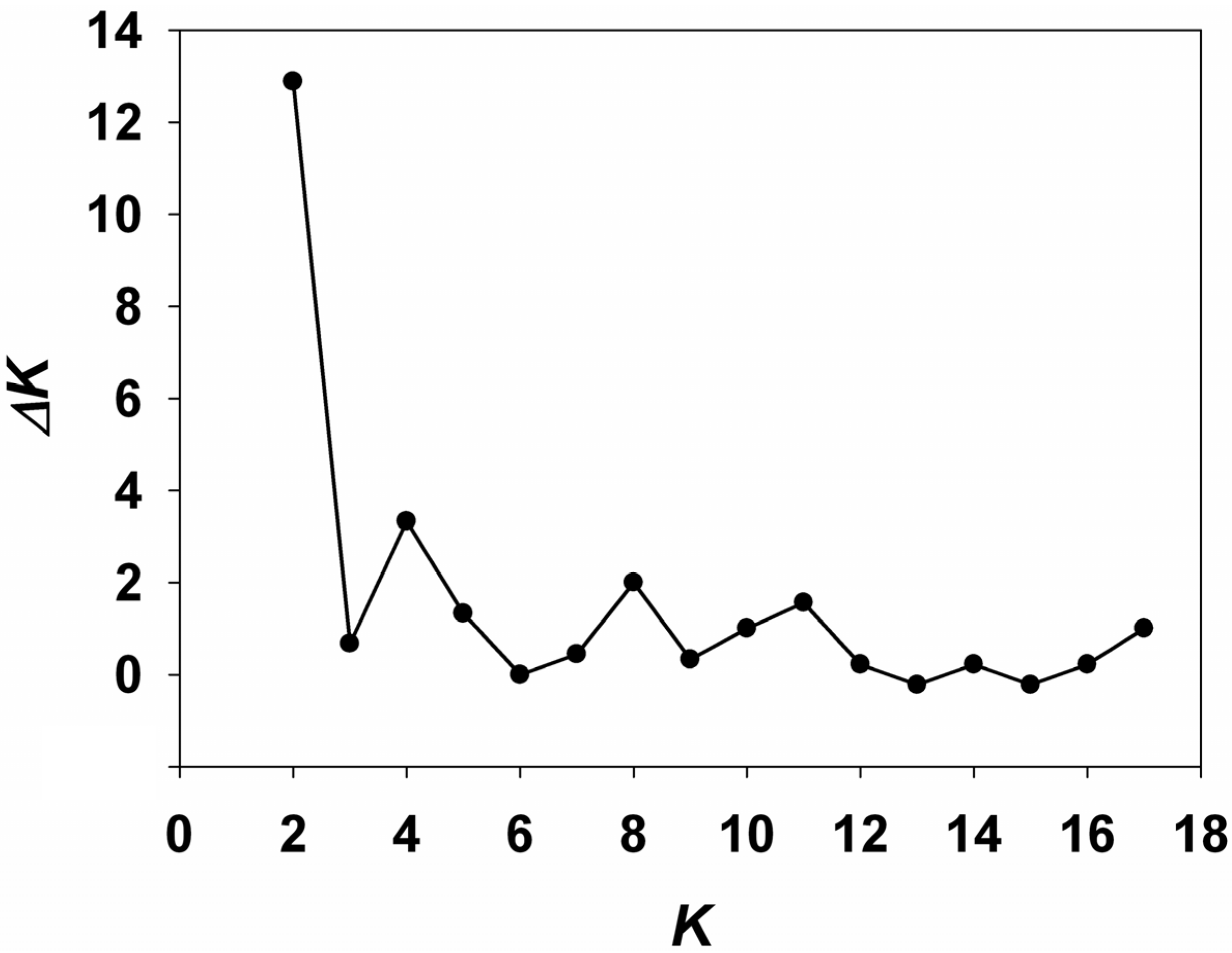
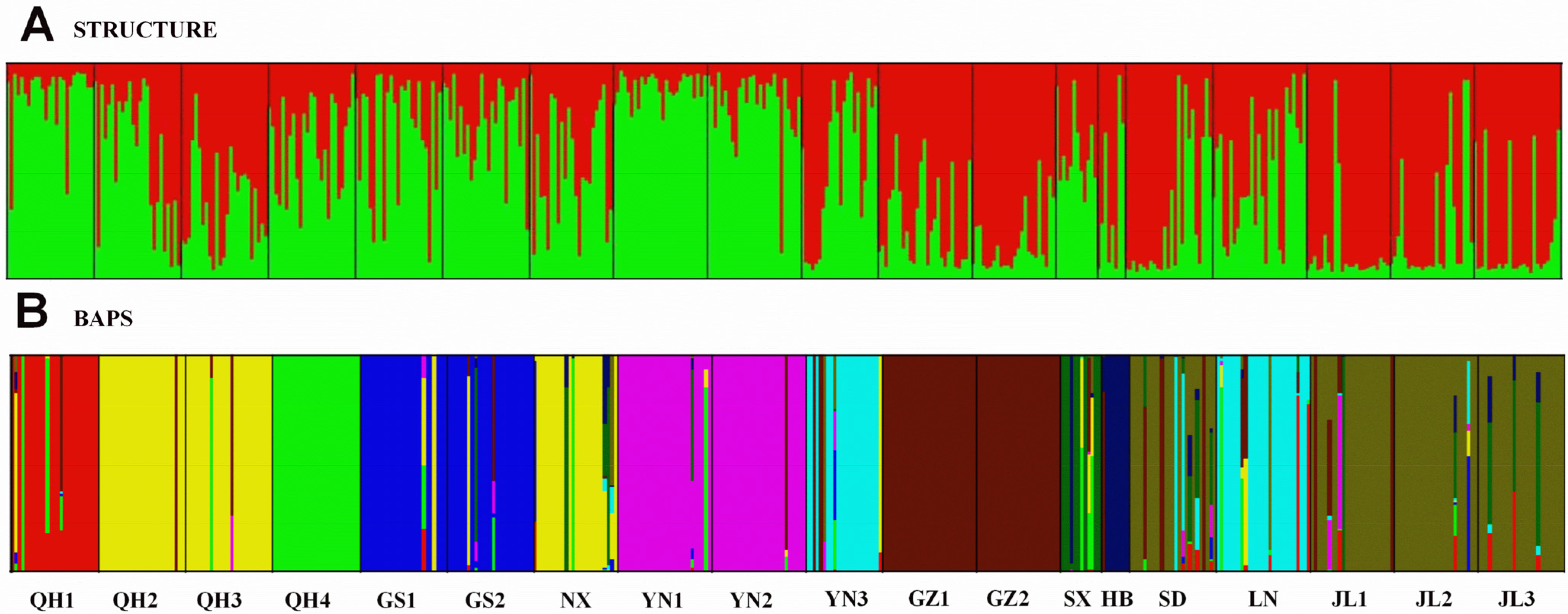
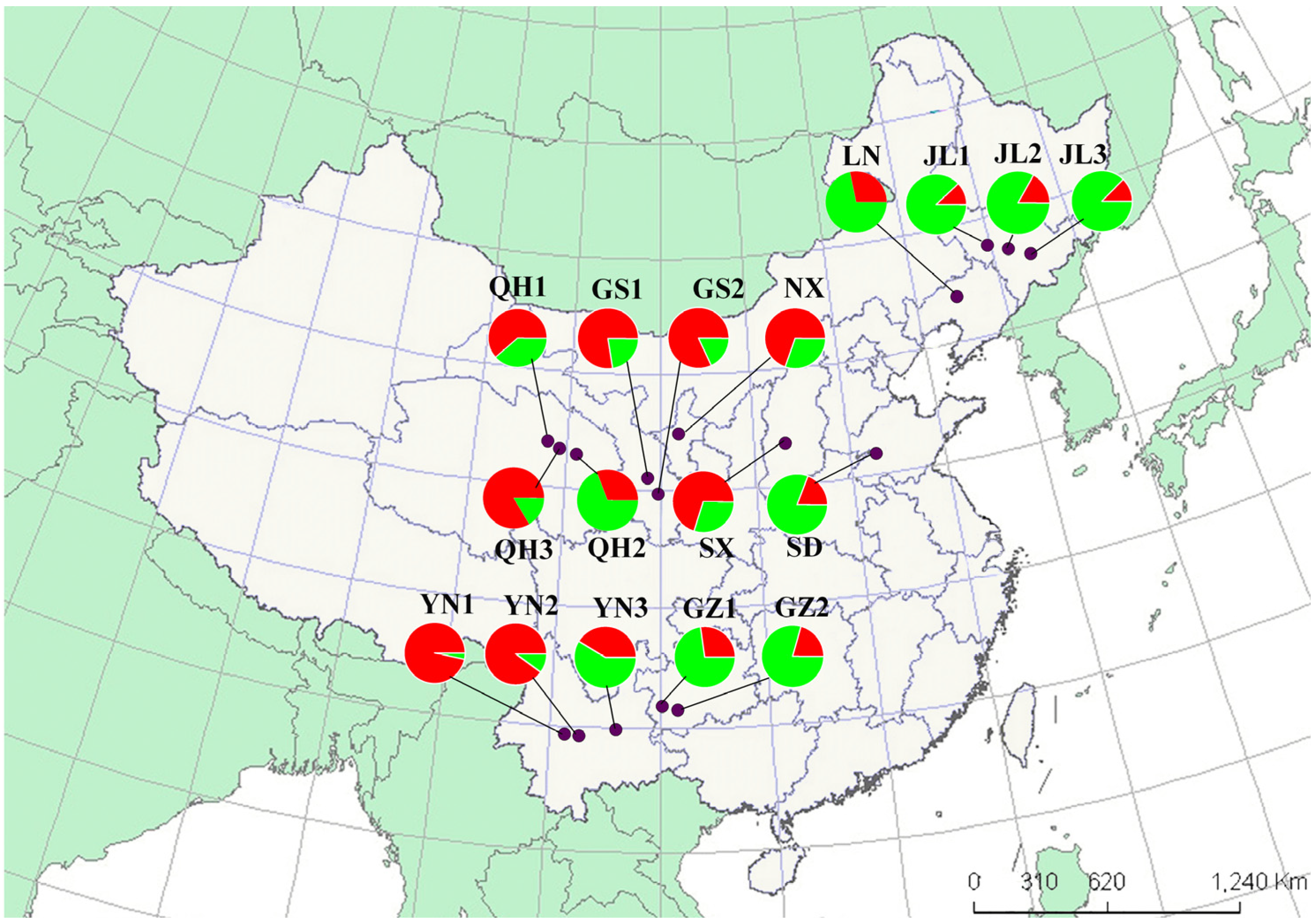
2.2. Analyses of Genetic Structure within Populations
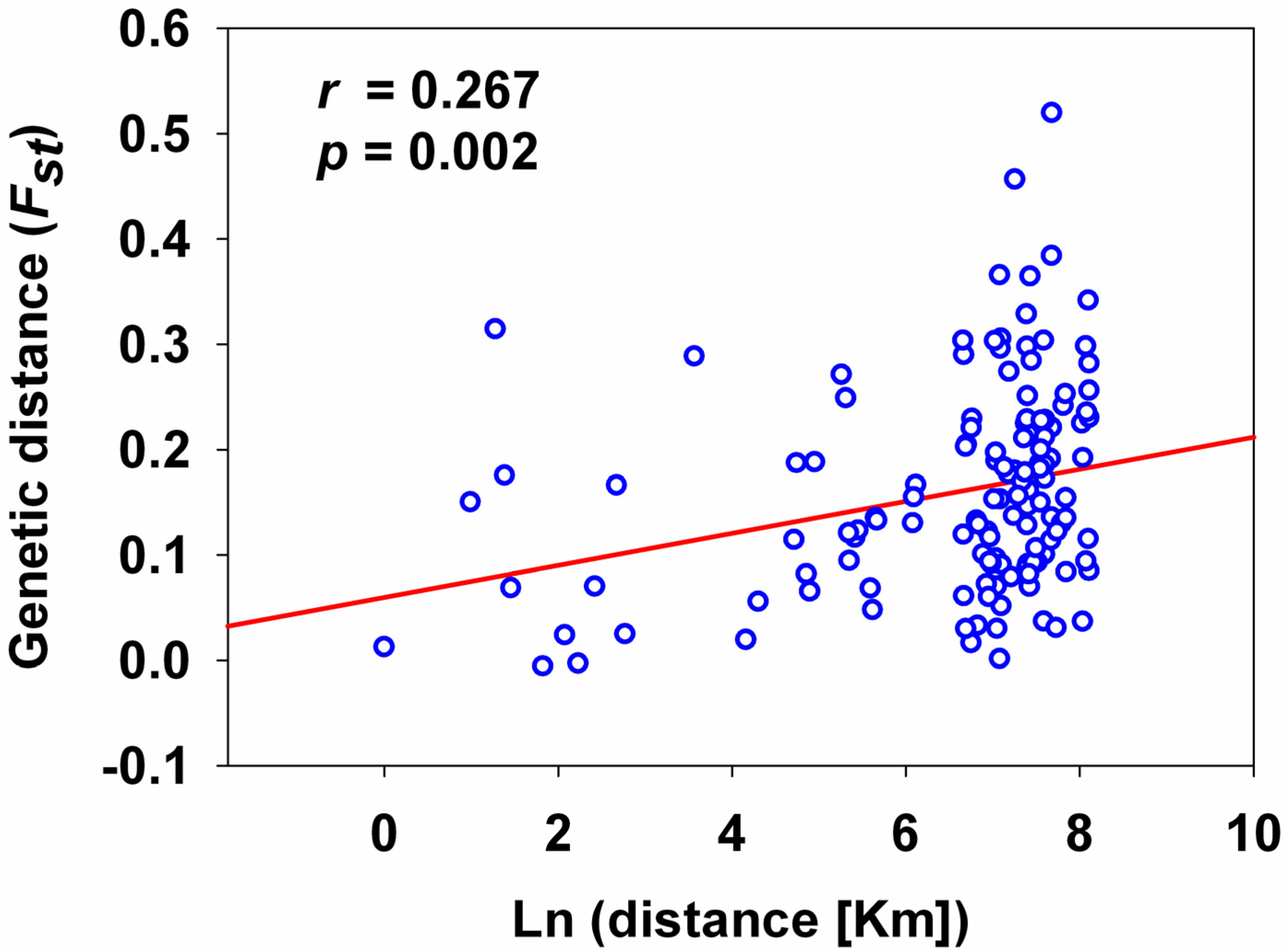
2.3. Analyses of Genetic Structure among Populations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Code | Date | Host | N | Na | Ne | Ho | He | Nei | Ar | Fis | p Values | Pwil |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Xining, Qinghai | QH1 | 2011.8 | Eggplant | 25 | 2.2857 | 1.6987 | 0.3200 | 0.3413 | 0.3345 | 2.0350 | −0.0094 | 0.4250 | 0.9844 |
| Xining, Qinghai | QH2 | 2012.9 | Eggplant | 25 | 2.1429 | 1.5074 | 0.3029 | 0.3144 | 0.3081 | 1.9549 | 0.0460 | 0.5966 | 0.9453 |
| Xining, Qinghai | QH3 | 2012.9 | Kidney bean | 25 | 2.1429 | 1.4428 | 0.2229 | 0.2499 | 0.2449 | 1.7926 | 0.1648 | 0.3768 | 0.6875 |
| Baiyin, Gansu | GS1 | 2011.8 | Tomato | 25 | 2.5714 | 1.8688 | 0.3143 | 0.3774 | 0.3698 | 2.2439 | 0.1919 | 0.1261 | 0.8906 |
| Baiyin, Gansu | GS2 | 2011.8 | Eggplant | 25 | 2.8571 | 1.7495 | 0.3386 | 0.3849 | 0.3771 | 2.2721 | 0.0916 | 0.2804 | 0.5000 |
| Yinchuan, Ningxia | NX | 2012.9 | Eggplant | 24 | 3.1429 | 1.5985 | 0.2229 | 0.3277 | 0.3208 | 2.2344 | 0.3338 | 0.2016 | 0.0547 |
| Xishuangbanna, Yunnan | YN1 | 2011.9 | Tomato | 27 | 3.0000 | 1.9985 | 0.4308 | 0.4563 | 0.4478 | 2.5229 | 0.0496 | 0.2631 | 0.9219 |
| Xishuangbanna, Yunnan | YN2 | 2011.9 | Eggplant | 27 | 3.0000 | 2.1305 | 0.4550 | 0.4836 | 0.4746 | 2.6001 | 0.0329 | 0.4914 | 0.9453 |
| Yuxi, Yunnan | YN3 | 2012.1 | Kidney bean | 22 | 5.0000 | 2.4447 | 0.2993 | 0.5061 | 0.4945 | 3.6000 | 0.4247 | 0.1700 | 0.1484 |
| Guiyang, Guizhou | GZ1 | 2011.7 | Tomato | 27 | 2.5714 | 1.6685 | 0.4011 | 0.3651 | 0.3583 | 2.2656 | −0.0462 | 0.4125 | 0.9219 |
| Guiyang, Guizhou | GZ2 | 2011.8 | Eggplant | 24 | 3.1429 | 1.5665 | 0.3003 | 0.3121 | 0.3055 | 2.2867 | −0.0203 | 0.5758 | 0.0156 |
| Yuncheng, Shanxi | SX | 2011.1 | Tomato | 12 | 3.4286 | 2.0678 | 0.3693 | 0.4408 | 0.4189 | 3.0106 | 0.1476 | 0.1863 | 0.3438 |
| Jinan, Shandong | SD | 2012.9 | Cucumber | 25 | 4.4286 | 1.9886 | 0.3026 | 0.4307 | 0.4219 | 3.0181 | 0.2721 | 0.1771 | 0.0195 |
| Fushun, Liaoning | LN | 2011.1 | Tomato | 27 | 3.4286 | 1.8539 | 0.2116 | 0.3700 | 0.3631 | 2.5496 | 0.2853 | 0.2208 | 0.2891 |
| Changchun, Jilin | JL1 | 2012.8 | Tomato | 24 | 2.4286 | 1.6735 | 0.3323 | 0.3495 | 0.3421 | 2.1044 | 0.0699 | 0.3141 | 0.9453 |
| Changchun, Jilin | JL2 | 2012.8 | Cucumber | 24 | 3.0000 | 1.7421 | 0.2307 | 0.3181 | 0.3113 | 2.3799 | 0.2063 | 0.1560 | 0.2813 |
| Changchun, Jilin | JL3 | 2012.9 | Pepper | 25 | 2.4286 | 1.4949 | 0.1068 | 0.2324 | 0.2276 | 2.0003 | 0.5812 | 0.0505 | 0.0781 |
| Mean | - | - | - | - | 3.0000 | 1.7938 | 0.3036 | 0.3683 | 0.3600 | 2.4042 | - | - | - |
| Locality | Population Code | Heterozygosity Excess p Values | ||
|---|---|---|---|---|
| IAM | TPM | SMM | ||
| Xining, Qinghai | QH1 | 0.0313 | 0.0313 | 0.1094 |
| Xining, Qinghai | QH2 | 0.0234 | 0.0781 | 0.2813 |
| Xining, Qinghai | QH3 | 0.3125 | 0.4063 | 0.8906 |
| Baiyin, Gansu | GS1 | 0.0469 | 0.3125 | 0.4063 |
| Baiyin, Gansu | GS2 | 0.3438 | 0.5781 | 0.5781 |
| Yinchuan, Ningxia | NX | 0.7109 | 0.9609 | 0.9883 |
| Xishuangbanna, Yunnan | YN1 | 0.0391 | 0.2188 | 0.2813 |
| Xishuangbanna, Yunnan | YN2 | 0.0195 | 0.1484 | 0.3438 |
| Yuxi, Yunnan | YN3 | 0.7656 | 0.9453 | 0.9883 |
| Guiyang, Guizhou | GZ1 | 0.0547 | 0.2188 | 0.4219 |
| Guiyang, Guizhou | GZ2 | 0.9453 | 0.9922 | 1.0000 |
| Yuncheng, Shanxi | SX | 0.4219 | 0.7188 | 0.9453 |
| Jinan, Shandong | SD | 0.9453 | 0.9883 | 0.9922 |
| Fushun, Liaoning | LN | 0.7109 | 0.7656 | 0.9727 |
| Changchun, Jilin | JL1 | 0.0234 | 0.0781 | 0.5000 |
| Changchun, Jilin | JL2 | 0.5781 | 0.7813 | 0.9609 |
| Changchun, Jilin | JL3 | 0.7188 | 0.9453 | 0.9922 |

| Population Code | QH1 | QH2 | QH3 | GS1 | GS2 | NX | YN1 | YN2 | YN3 | GZ1 | GZ2 | SX | SD | LN | JL1 | JL2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| QH2 | 0.1661 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| QH3 | 0.2883 | 0.3145 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| GS1 | 0.1164 | 0.1208 | 0.2489 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| GS2 | 0.1232 | 0.0943 | 0.2711 | −0.0057 | - | - | - | - | - | - | - | - | - | - | - | - |
| NX | 0.1664 | 0.1549 | 0.1301 | 0.1352 | 0.1327 | - | - | - | - | - | - | - | - | - | - | - |
| YN1 | 0.1615 | 0.2246 | 0.2508 | 0.0896 | 0.0877 | 0.2003 | - | - | - | - | - | - | - | - | - | - |
| YN2 | 0.1282 | 0.2148 | 0.2978 | 0.0913 | 0.0928 | 0.2275 | 0.0126 | - | - | - | - | - | - | - | - | - |
| YN3 | 0.1463 | 0.0699 | 0.2289 | 0.0826 | 0.0816 | 0.1499 | 0.1755 | 0.1503 | - | - | - | - | - | - | - | - |
| GZ1 | 0.1529 | 0.0015 | 0.2958 | 0.1144 | 0.0969 | 0.1768 | 0.2045 | 0.2025 | 0.0609 | - | - | - | - | - | - | - |
| GZ2 | 0.3056 | 0.0517 | 0.3656 | 0.1973 | 0.1872 | 0.2737 | 0.2900 | 0.3035 | 0.1195 | 0.0248 | - | - | - | - | - | - |
| SX | 0.0703 | 0.0856 | 0.1530 | 0.0723 | 0.0603 | 0.0907 | 0.1326 | 0.1289 | 0.0331 | 0.0817 | 0.1879 | - | - | - | - | - |
| SD | 0.1891 | 0.0301 | 0.3029 | 0.1223 | 0.1169 | 0.1830 | 0.2293 | 0.2204 | 0.0165 | 0.0194 | 0.0558 | 0.1008 | - | - | - | - |
| LN | 0.1002 | 0.0370 | 0.3036 | 0.0929 | 0.1064 | 0.1698 | 0.2250 | 0.1922 | 0.0368 | 0.0309 | 0.1221 | 0.0790 | 0.0238 | - | - | - |
| JL1 | 0.1914 | 0.1139 | 0.3838 | 0.1726 | 0.1855 | 0.2842 | 0.2562 | 0.2301 | 0.0848 | 0.0837 | 0.1541 | 0.1784 | 0.0300 | 0.0478 | - | - |
| JL2 | 0.1803 | 0.1370 | 0.4562 | 0.1864 | 0.1819 | 0.3284 | 0.2981 | 0.2354 | 0.0939 | 0.1298 | 0.2418 | 0.1562 | 0.0916 | 0.0654 | 0.0686 | - |
| JL3 | 0.2210 | 0.1356 | 0.5194 | 0.2280 | 0.2118 | 0.3644 | 0.3415 | 0.2817 | 0.1150 | 0.1351 | 0.2527 | 0.2109 | 0.0936 | 0.0684 | 0.0703 | −0.0031 |


2.4. Discussion
3. Experimental Section
3.1. Field Sampling

3.2. DNA Extraction and Microsatellite Genotyping
| Code | GenBank Number | Primer (5'–3') | Annealing Temperature (°C) | Range (bp) |
|---|---|---|---|---|
| Tvap-1-2 | GF112025 | PrimerA: CTGTGAATCCCTCAGAAATC PrimerB: TGACCTCTCTCAGGCTTTTA | 57 | 180–238 |
| Tvap-2-2C | GF112021 | PrimerA: CTGAAAGTCTTATTAGAGCC PrimerB: CTAACTGATTCCATAGTCG | 55 | 150–220 |
| Tvap-3-3 | GF112019 | PrimerA: CGCAAATCATACTTCCTTTC PrimerB: AAATACAGGCGACTCATGTC | 55 | 222–238 |
| Tvap-3-2 | GF112017 | PrimerA: GGAGGTCATTACTCATTTCG PrimerB: CATAAATTTTCGGCTCACTC | 55 | 176–184 |
| Tvap-1-1C | GF112015 | PrimerA: GAGACTCCACGATGTCTGTC PrimerB: TTCCCCTATCGTATGTTCAC | 57 | 193–233 |
| Tvap-1-3 | GF112026 | PrimerA: TATAGGGGTGTTGTGGTGTT PrimerB: CGCTACCAAATCGTAATTAC | 55 | 147–197 |
| Tvap-1-4 | GF112020 | PrimerA: GATTTAGCCCAGTTCATTTG PrimerB: CTTCAGTTGAGCTGCTGATG | 55 | 218–268 |
3.3. Analyses of Genetic Diversity
3.4. Analyses of Genetic Structure within Populations
3.5. Analyses of Genetic Structure among Populations
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- López, Y.I.A.; Martínez-Gallardo, N.A.; Ramírez-Romero, R.; López, M.G.; Sánchez-Hernández, C. Cross-kingdom effects of plant-plant signaling via volatile organic compounds emitted by tomato (Solanum lycopersicum) plants infested by the greenhouse whitefly (Trialeurodes vaporariorum). J. Chem. Phys. 2012, 38, 1376–1386. [Google Scholar]
- Simmonds, M.S.J.; Manlove, J.D.; Blaney, W.M.; Khambay, B.P.S. Effects of selected botanical insecticides on the behaviour and mortality of the glasshouse whitefly Trialeurodes. vaporariorum and the parasitoid Encarsia formosa. Entomol. Exp. Appl. 2002, 102, 39–47. [Google Scholar]
- De Vis, R.M.J.; Fuentes, L.E.; van Lenteren, J.C. Life history of Amitus fuscipennis (Hym., Platygastridae) as parasitoid of the greenhouse whitefly Trialeurodes vaporariorum (Hom., Aleyrodidae) on tomato as function of temperature. J. Appl. Entomol. 2002, 126, 24–33. [Google Scholar] [CrossRef]
- Bouhous, M.; Larous, L. Efficiency of the entomopathogenic fungus Verticillium. lecanii in the biological control of Trialeurodes vaporariorum, (Homoptera: Aleyrodidae), a greenhouse culture pest. Afr. J. Microbiol. Res. 2012, 6, 2435–2442. [Google Scholar]
- Puthoff, D.P.; Holzer, F.M.; Perring, T.M.; Walling, L.L. Tomato pathogenesis-related protein genes are expressed in response to Trialeurodes vaporariorum and Bemisia. tabaci biotype B feeding. J. Chem. Ecol. 2010, 36, 1271–1285. [Google Scholar]
- Lourenção, A.L.; Alves, A.C.; Fugi, C.G.; Matos, E.S. Outbreaks of Trialeurodes vaporariorum (West.) (Hemiptera: Aleyrodidae) under field conditions in the State of São Paulo, Brazil. Neotrop. Entomol. 2008, 37, 89–91. [Google Scholar] [CrossRef]
- Russell, L.M. Hosts and distribution of five species of Trialeurodes. (Homoptera: Aleyrodidae). Ann. Entomol. Soc. Am. 1963, 56, 149–153. [Google Scholar]
- Landa, Z.; Osborne, L.; Lopez, F.; Eyal, J. A bioassay for determining pathogenicity of entomogenous fungi on whiteflies. Biol. Control. 1994, 4, 341–350. [Google Scholar] [CrossRef]
- Duffus, J.E.; Johnstone, G.R. Beet pseudo-yellows virus in Tasmania: The first report of a whitefly transmitted virus in Australiasia. Aust. Plant. Path. 1981, 10, 68–69. [Google Scholar] [CrossRef]
- Yamashita, S.; Doi, Y.; Yora, K.; Yoshino, M. Cucumber yellows virus: Its transmission by the greenhouse whitefly, Trialeurodes vaporariorum (Westwood), and the yellowing disease of cucumber and muskmelon caused by the virus. Ann. Phytopath. Soc. Jpn. 1979, 45, 484–496. [Google Scholar] [CrossRef]
- Coffin, R.S.; Coutts, R.H.A. The occurrence of beet pseudo yellows virus in England. Plant Pathol. 1990, 39, 632–635. [Google Scholar] [CrossRef]
- Jones, D.R. Plant viruses transmitted by whiteflies. Eur. J. Plant Pathol. 2003, 109, 195–219. [Google Scholar] [CrossRef]
- Helgesen, R.G.; Tauber, M.J. Biological control of greenhouse whitefly, Trialeurodes vaporariorum (Aleyrodidae: Homoptera), on short-term crops by manipulating biotic and abiotic factors. Can. Entomol. 1974, 106, 1175–1188. [Google Scholar] [CrossRef]
- Xiang, Y.Y.; Li, Z.Z.; Zhang, F.; Luo, C. Progress of study on Bemisia tabaci (Gennadius) and Trialeurodes vaprorariorum (Westwood). J. Mt. Agric. Biol. 2004, 23, 352–359. [Google Scholar]
- Zhang, Z.L.; Chen, W.L.; Wang, J. Preliminary observation and control of the greenhouse whitefly, Trialeurodes. vaporariorum in the suburban regions of Beijing. Entomol. Knowl. 1980, 17, 158–160. [Google Scholar]
- Chu, D.; Gao, C.S.; de Barro, P.; Zhang, Y.J.; Wan, F.H. Investigation of the genetic diversity of an invasive whitefly (Bemisia tabaci) in China using both mitochondrial and nuclear DNA markers. Bull. Entomol. Res. 2011, 101, 467–475. [Google Scholar] [CrossRef]
- Men, Q.L.; Chen, M.H.; Zhang, Y.L.; Feng, J.N. Genetic structure and diversity of a newly invasive species, the codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae) in China. Biol. Invasions 2012, 15, 447–458. [Google Scholar]
- Chu, D.; Pan, H.P.; Li, X.C.; Guo, D.; Tao, Y.L.; Liu, B.M.; Zhang, Y.J. Spatial genetic heterogeneity in populations of a newly invasive whitefly in China revealed by a nation-wide field survey. PLoS One 2013, 8, e79997. [Google Scholar]
- Yang, X.M.; Sun, J.T.; Xue, X.F.; Li, J.B.; Hong, X.Y. Invasion genetics of the western flower thrips in China: Evidence for genetic bottleneck, hybridization and bridgehead effect. PLoS One 2012, 7, e34567. [Google Scholar]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J. The population biology of invasive species. Annu. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef]
- Kirk, H.; Dorn, S.; Mazzi, D. Molecular genetics and genomics generate new insights into invertebrate pest invasions. Evol. Appl. 2013, 6, 842–856. [Google Scholar] [CrossRef]
- Selkoe, K.A.; Toonen, R.J. Microsatellites for ecologists: A practical guide to using and evaluating microsatellite markers. Ecol. Lett. 2006, 9, 615–629. [Google Scholar] [CrossRef]
- Teng, Z.Q.; Kang, L. Microsatellites reveal the genetic structure of thelytokous strains of the migratory locust. Insect Sci. 2007, 14, 193–199. [Google Scholar] [CrossRef]
- Sun, J.T.; Lian, C.L.; Navajas, M.; Hong, X.Y. Microsatellites reveal a strong subdivision of genetic structure in Chinese populations of the mite Tetranychus urticae Koch (Acari: Tetranychidae). BMC Med. Genet. 2012, 13. [Google Scholar] [CrossRef]
- Cornuet, J.M.; Luikart, G. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics 1996, 144, 2001–2014. [Google Scholar]
- Van, O.C.; Hutchinson, W.F.; Wills, D.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–553. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Corander, J.; Waldmann, P.; Sillanpaa, M.J. Bayesian analysis of genetic differentiation between populations. Genetics 2003, 163, 367–374. [Google Scholar]
- Corander, J.; Sirén, J.; Arjas, E. Bayesian spatial modeling of genetic population structure. Comput. Stat. 2008, 23, 111–129. [Google Scholar] [CrossRef]
- Sanz, N.; Araguas, R.M.; Vidal, O.; Diez-del-Molino, D.; Fernández-Cebrián, R.; García-Marín, J.L. Genetic characterization of the invasive mosquitofish (Gambusia spp.) introduced to Europe: Population structure and colonization routes. Biol. Invasions 2013, 15, 2333–2346. [Google Scholar] [CrossRef]
- Ghabooli, S.; Shiganova, T.A.; Zhan, A.; Cristescu, M.E.; Eghtesadi-Araghi, P.; MacIsaac, H.J. Multiple introductions and invasion pathways for the invasive ctenophore Mnemiopsis leidyi in Eurasia. Biol. Invasions 2011, 13, 679–690. [Google Scholar] [CrossRef]
- Cabezas, M.P.; Xavier, R.; Branco, M.; Santos, A.M.; Guerra-García, J.M. Invasion history of Caprella scaura Templeton, 1836 (Amphipoda: Caprellidae) in the Iberian Peninsula: Multiple introductions revealed by mitochondrial sequence data. Biol. Invasions 2014. [Google Scholar] [CrossRef]
- Reem, E.; Douek, J.; Katzir, G.; Rinkevich, B. Long-term population genetic structure of an invasive urochordate: The ascidian Botryllus schlosseri. Biol. Invasions 2013, 15, 225–241. [Google Scholar] [CrossRef]
- Roman, J.; Darling, J.A. Paradox lost: Genetic diversity and the success of aquatic invasions. Trends Ecol. Evol. 2007, 22, 454–464. [Google Scholar] [CrossRef]
- Suarez, A.V.; Tsutsui, N.D. The evolutionary consequences of biological invasions. Mol. Ecol. 2008, 17, 351–360. [Google Scholar] [CrossRef]
- Dakin, E.E.; Avise, J.C. Microsatellite null alleles in parentage analysis. Heredity 2004, 93, 504–509. [Google Scholar] [CrossRef]
- Ovcarenko, I.; Clouet, C.; Knott, K.; Tsagkarakou, A.; Gauthier, N. Thirteen polymorphic microsatellite loci and PCR multiplexing in the greenhouse whitefly, Trialeurodes vaporariorum Westwood (Homoptera: Aleyrodidae). Mol. Ecol. Res. 2013, 13, 341–343. [Google Scholar] [CrossRef] [Green Version]
- POPGENE, version 1.31. The User-Friendly Shareware for Population Genetic Analysis; Molecular Biology and Biotechnology Centre, University of Alberta: Edmonton, AB, Canada, 1997.
- Franks, S.J.; Pratt, P.D.; Tsutsui, N.D. The genetic consequences of a demographic bottleneck in an introduced biological control insect. Conserv. Genet. 2010, 12, 201–211. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1955, 86, 248–249. [Google Scholar]
- Di Rienzo, A.; Peterson, A.C.; Garcza, J.C.; Valdes, A.M.; Slatkin, M. Mutational processes of simple sequence repeat loci in human populations. Proc. Natl. Acad. Sci. USA 1994, 91, 3166–3170. [Google Scholar] [CrossRef]
- Jensen, J.L.; Bohonak, A.J.; Kelley, S.T. Isolation by distance web service. BMC Genet. 2005, 6. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Res. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: Asimulation study. Mol. Ecol. Notes 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gao, R.-R.; Zhang, W.-P.; Wu, H.-T.; Zhang, R.-M.; Zhou, H.-X.; Pan, H.-P.; Zhang, Y.-J.; Brown, J.K.; Chu, D. Population Structure of the Greenhouse Whitefly, Trialeurodes vaporariorum (Westwood), an Invasive Species from the Americas, 60 Years after Invading China. Int. J. Mol. Sci. 2014, 15, 13514-13528. https://doi.org/10.3390/ijms150813514
Gao R-R, Zhang W-P, Wu H-T, Zhang R-M, Zhou H-X, Pan H-P, Zhang Y-J, Brown JK, Chu D. Population Structure of the Greenhouse Whitefly, Trialeurodes vaporariorum (Westwood), an Invasive Species from the Americas, 60 Years after Invading China. International Journal of Molecular Sciences. 2014; 15(8):13514-13528. https://doi.org/10.3390/ijms150813514
Chicago/Turabian StyleGao, Rui-Rui, Wen-Ping Zhang, Huai-Tong Wu, Rui-Ming Zhang, Hong-Xu Zhou, Hui-Peng Pan, You-Jun Zhang, Judith K. Brown, and Dong Chu. 2014. "Population Structure of the Greenhouse Whitefly, Trialeurodes vaporariorum (Westwood), an Invasive Species from the Americas, 60 Years after Invading China" International Journal of Molecular Sciences 15, no. 8: 13514-13528. https://doi.org/10.3390/ijms150813514