The Effects of Synthetic Oligopeptide Derived from Enamel Matrix Derivative on Cell Proliferation and Osteoblastic Differentiation of Human Mesenchymal Stem Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
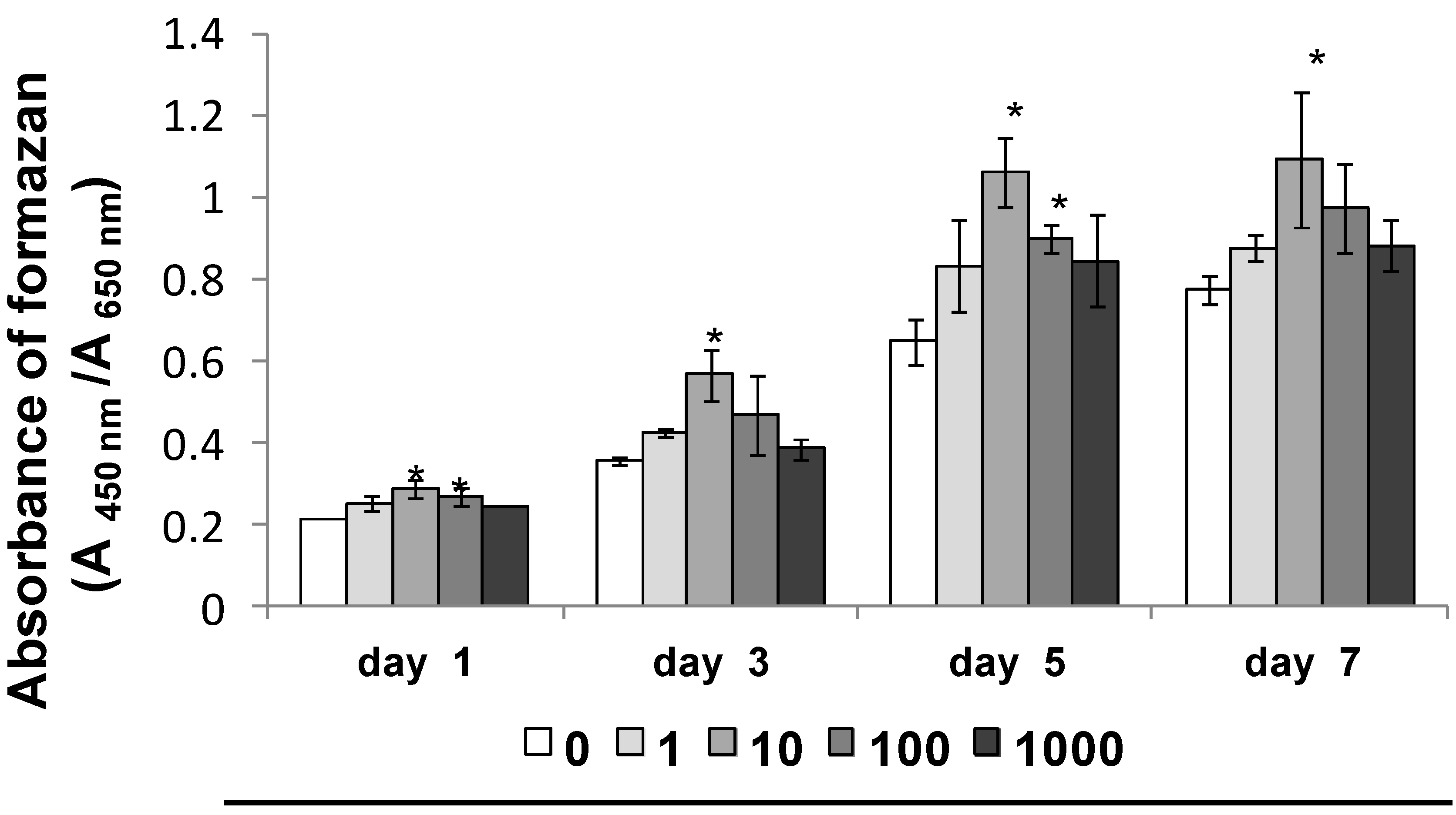
2.1.1. Cell Proliferation

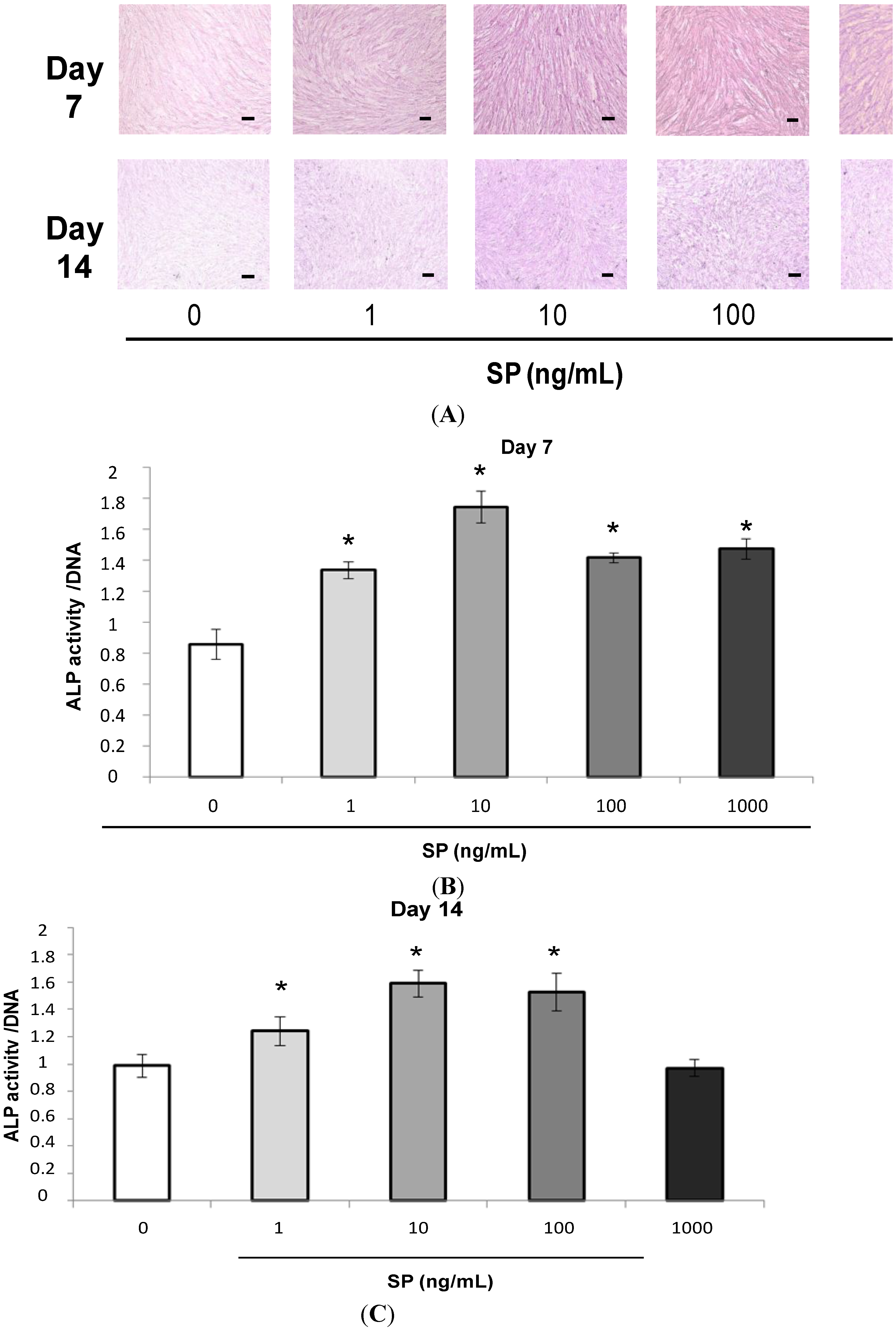
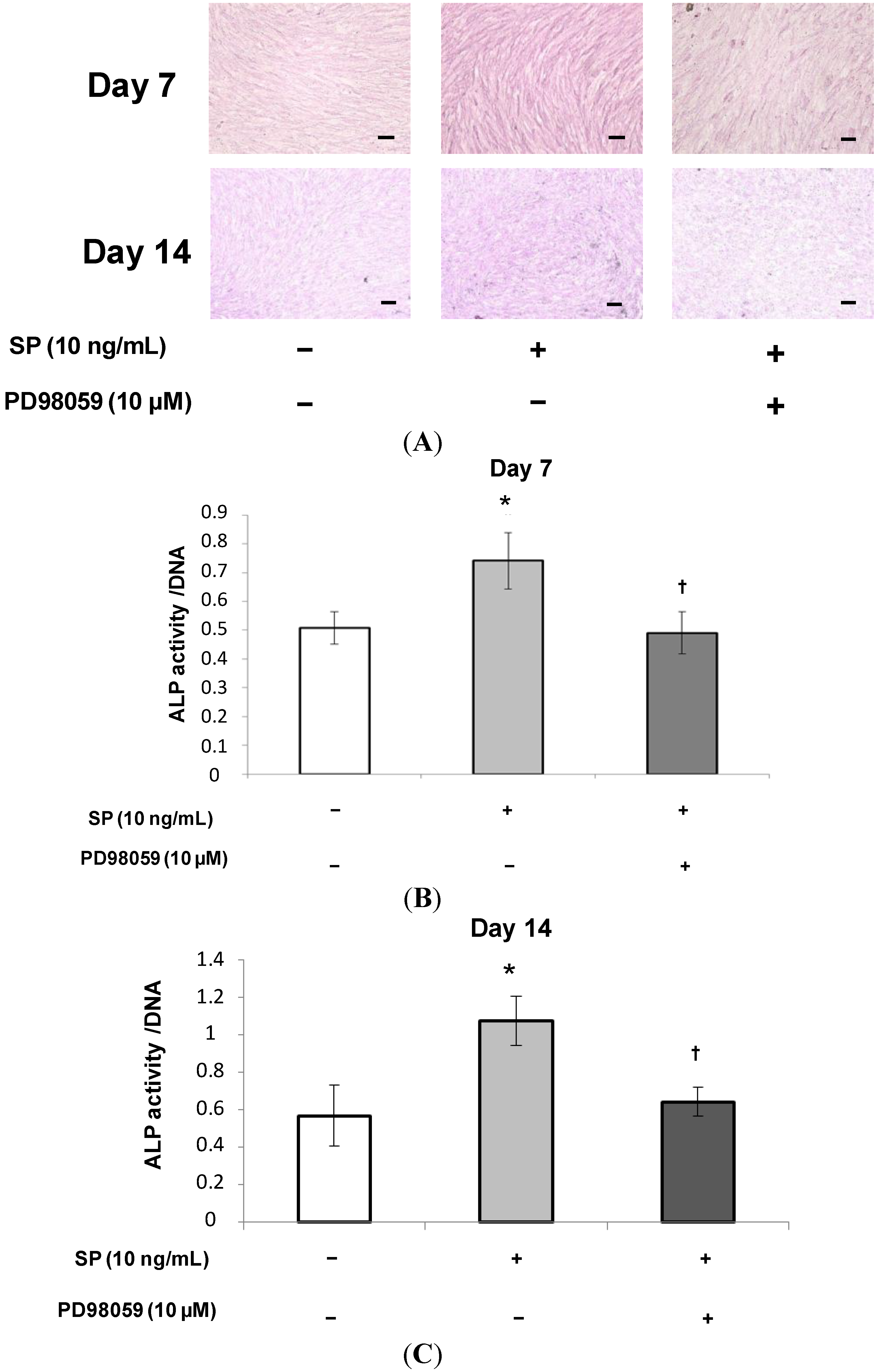
2.1.2. Alkaline Phosphatase (ALP) Staining and Measurement of ALP Activity

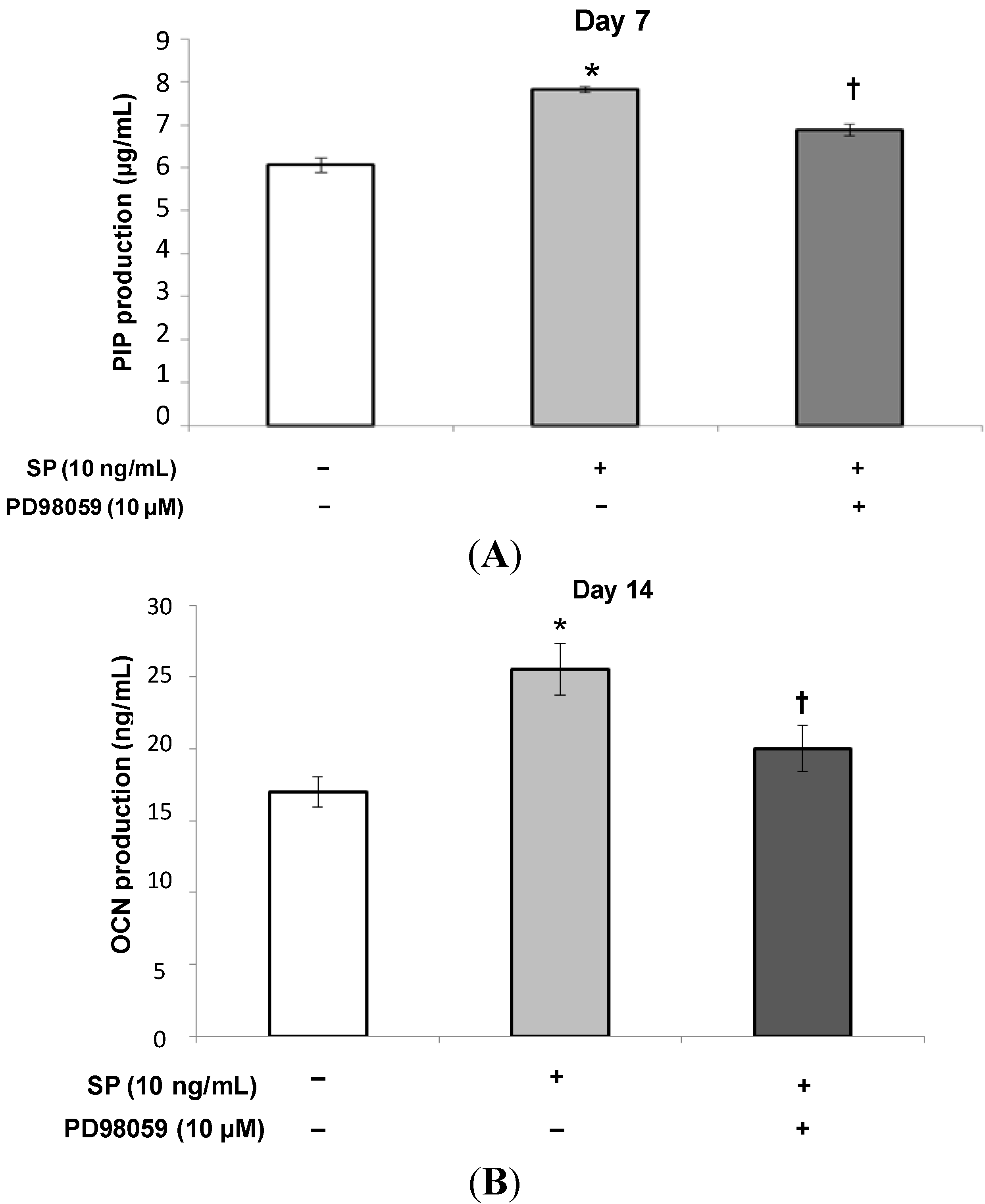
2.1.3. Procollagen Type 1 C-Peptide (PIP) and Osteocalcin (OCN) Production

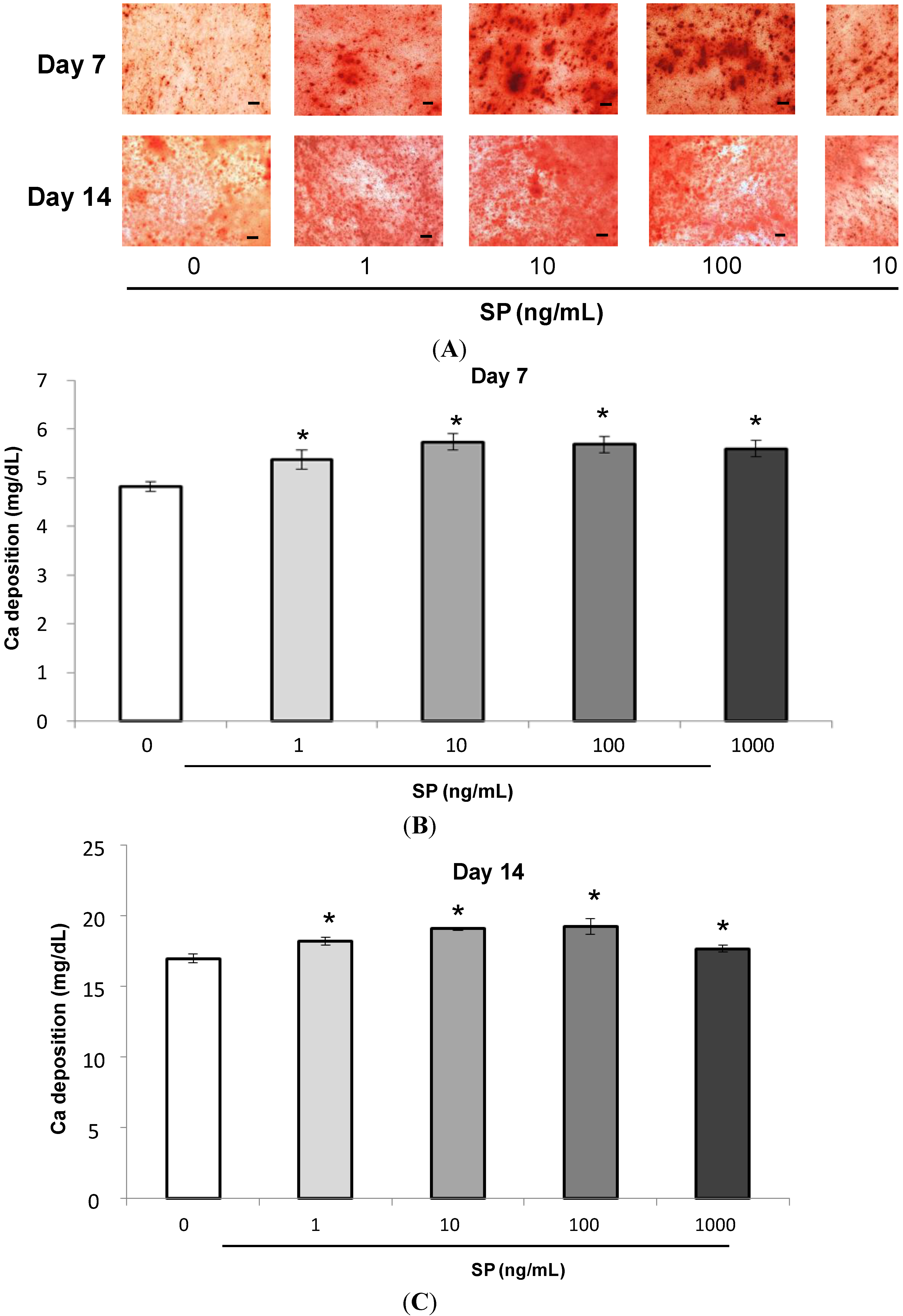
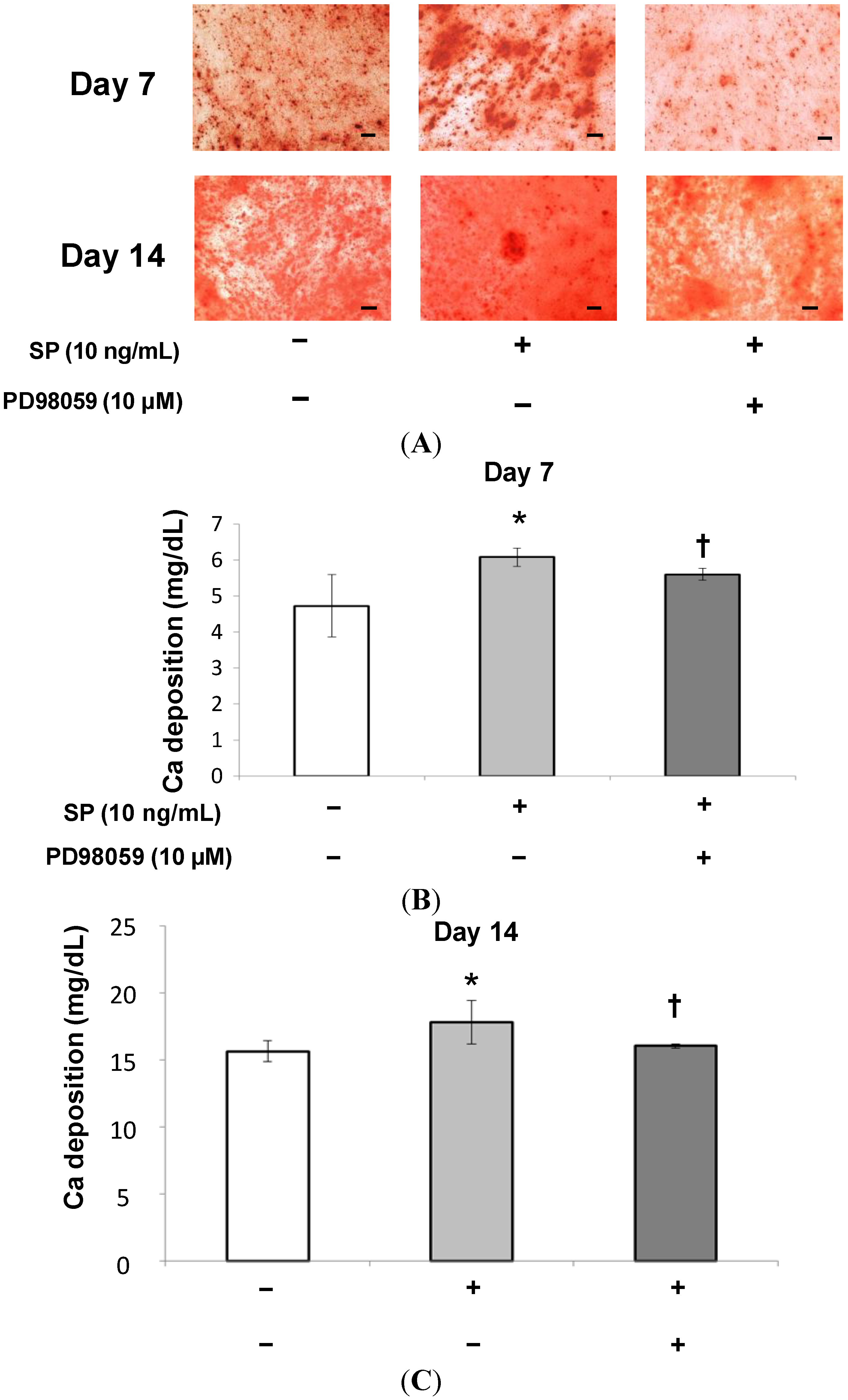
2.1.4. Mineralization Assay (Alizarin Red Staining and Extracellular Calcium Deposition)

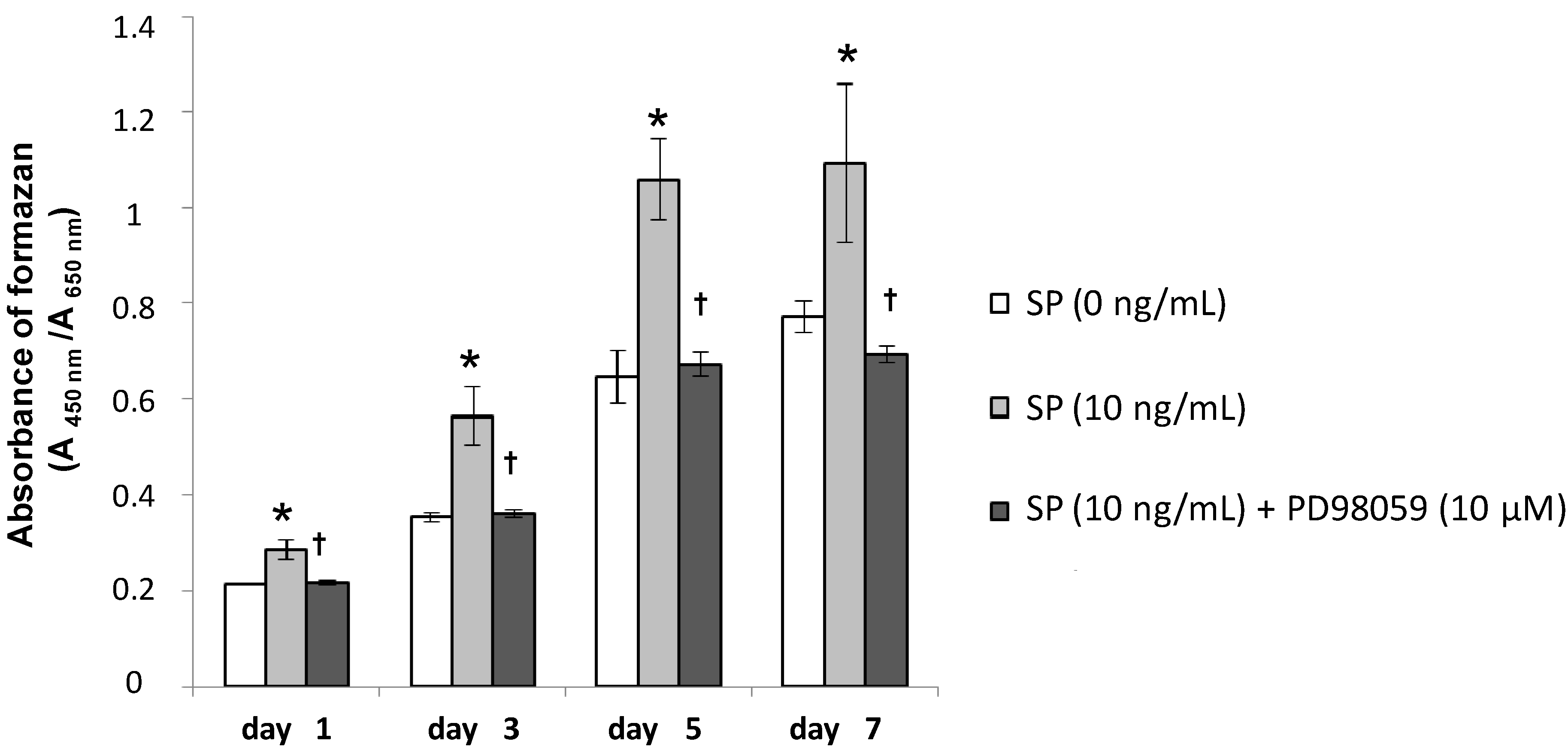
2.1.5. Effects of an Extracellular Signal-Related Kinases (ERK) 1/2 Inhibitor on SP-Induced Cell Proliferation and Osteoblastic Differentiation




2.2. Discussion
3. Experimental Section
3.1. Preparation of Synthetic Oligopeptide and Cell Culture
3.2. Cell Proliferation Assay
3.3. Alkaline Phosphatase (ALP) Staining and Measurement of ALP Activity
3.4. Measurement of Procollagen Type 1 C-Peptide (PIP) and Osteocalcin (OCN) Production
3.5. Mineralization Assay (Alizarin Red Staining and Measurement of Extracellular Calcium Deposition)
3.6. Effect of Extracellular Signal-Related Kinases (ERK) 1/2 Inhibition on Cell Proliferation and Osteoblast Differentiation
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nanci, A.; Bosshardt, D.D. Structure of periodontal tissues in health and disease. Periodontology 2000 2006, 40, 11–28. [Google Scholar] [CrossRef]
- Jung, R.E.; Glauser, R.; Schärer, P.; Hämmerle, C.H.; Sailer, H.F.; Weber, F.E. Effect of rhBMP-2 on guided bone regeneration in humans. Clin. Oral Implants Res. 2003, 14, 556–568. [Google Scholar] [CrossRef]
- Nevins, M.; Camelo, M.; Nevins, M.L.; Schenk, R.K.; Lynch, S.E. Periodontal regeneration in humans using recombinant human platelet-derived growth factor-BB (rhPDGF-BB) and allogenic bone. J. Periodontol. 2003, 74, 1282–1292. [Google Scholar] [CrossRef]
- Heijl, L. Periodontal regeneration with enamel matrix derivative in one human experimental defect—A case report. J. Clin. Periodontol. 1997, 24, 693–696. [Google Scholar] [CrossRef]
- Maycock, J.; Wood, S.R.; Brookes, S.J.; Shore, R.C.; Robinson, C.; Kirkham, J. Characterization of a porcine amelogenin preparation, EMDOGAIN, a biological treatment for periodontal disease. Connect. Tissue Res. 2002, 43, 472–476. [Google Scholar] [CrossRef]
- Hammarström, L. Enamel matrix, cementum development and regeneration. J. Clin. Periodontol. 1997, 24, 658–668. [Google Scholar] [CrossRef]
- Hammarström, L.; Heijl, L.; Gestrelius, S. Periodontal regeneration in a buccal dehiscence model in monkeys after application of enamel matrix proteins. J. Clin. Periodontol. 1997, 24, 669–677. [Google Scholar] [CrossRef]
- Bosshardt, D.D. Biological mediators and periodontal regeneration: A review of enamel matrix proteins at the cellular and molecular levels. J. Clin. Periodontol. 2008, 35, 87–105. [Google Scholar] [CrossRef]
- Grandin, H.M.; Gemperli, A.C.; Dard, M. Enamel matrix derivative: A review of cellular effects in vitro and a model of molecular arrangement and functioning. Tissue Eng. 2012, 18, 181–202. [Google Scholar]
- Gestrelius, S.; Andersson, C.; Lidström, D.; Hammarström, L.; Somerman, M. In vitro studies on periodontal ligament cells and enamel matrix derivative. J. Clin. Periodontol. 1997, 24, 685–692. [Google Scholar] [CrossRef]
- Kim, N.H.; Tominaga, K.; Tanaka, A. Analysis of eosinophilic round bodies formed after injection of enamel matrix derivative into the backs of rats. J. Periodontol. 2005, 76, 1934–1941. [Google Scholar] [CrossRef]
- Yuan, K.; Hsu, C.W.; Tsai, W.H. The induction and possible subsequent effect of human antibodies against porcine enamel matrix derivative. J. Periodontol. 2006, 77, 1355–1361. [Google Scholar] [CrossRef]
- Shinnick, T.M.; Sutcliffe, J.G.; Green, N.; Lerner, R.A. Synthetic peptide immunogens as vaccines. Annu. Rev. Microbiol. 1983, 37, 425–446. [Google Scholar] [CrossRef]
- Lerner, R.A. Tapping the immunological repertoire to produce antibodies of predetermined specificity. Nature 1982, 299, 593–596. [Google Scholar] [CrossRef]
- Hida, T.; Tominaga, K.; Tanaka, A. Tissue Reaction to synthetic oligopeptide derived from enamel matrix derivative in rats. Oral Sci. Int. 2010, 7, 26–33. [Google Scholar] [CrossRef]
- Noguchi, M.; Tominaga, K.; Tanaka, A.; Ueda, M. Hard tissue formation induced by synthetic oligopeptide derived from an enamel matrix derivative. Oral Med. Pathol. 2012, 16, 75–80. [Google Scholar] [CrossRef]
- Kawanaka, A.; Tominaga, K.; Tanaka, A. Effect of peptide derived from Emdogain® on human periodontal ligament fibroblasts. J. Osaka Dent. Univ. 2009, 43, 111–117. [Google Scholar]
- Yasui, N.; Taguchi, Y.; Tanaka, A.; Ueda, M.; Umeda, M. Biological effects of emdogain (R)-derived oligopeptides on rat bone marrow cells in vitro. J. Oral Tissue Eng. 2012, 9, 126–135. [Google Scholar]
- Taguchi, Y.; Yasui, N.; Takahashi, S.; Tominaga, K.; Kato, H.; Komasa, S.; Shida, M.; Hayashi, H.; Tanaka, A.; Umeda, M. Hard tissue formation by human periodontal ligament fibroblast cells treated with an emdogain®-derived oligopeptide in vitro. J. Hard Tissue Biol. 2012, 21, 375–384. [Google Scholar] [CrossRef]
- Kato, H.; Katayama, N.; Taguchi, Y.; Tominaga, K.; Umeda, M.; Tanaka, A. A synthetic oligopeptide derived from enamel matrix derivative promotes the differentiation of human periodontal ligament stem cells into osteoblast-like cells with increased mineralization. J. Periodontol. 2013, 84, 1476–1483. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef]
- Hasegawa, N.; Kawaguchi, H.; Hirachi, A.; Takeda, K.; Mizuno, N.; Nishimura, M.; Koike, C.; Tsuji, K.; Iba, H.; Kato, Y.; et al. Behavior of transplanted bone marrow-derived mesenchymal stem cells in periodontal defects. J. Periodontol. 2006, 77, 1003–1007. [Google Scholar] [CrossRef]
- Yang, Y.; Rossi, F.M.; Putnins, E.E. Periodontal regeneration using engineered bone marrow mesenchymal stromal cells. Biomaterials 2010, 31, 8574–8582. [Google Scholar] [CrossRef]
- Tsumanuma, Y.; Iwata, T.; Washio, K.; Yoshida, T.; Yamada, A.; Takagi, R.; Ohno, T.; Lin, K.; Yamato, M.; Ishikawa, I.; et al. Comparison of different tissue-derived stem cell sheets for periodontal regeneration in a canine 1-wall defect model. Biomaterials 2011, 32, 5819–5825. [Google Scholar] [CrossRef]
- Junttila, M.R.; Li, S.P.; Westermarck, J. Phosphatase-mediated crosstalk between MAPK signaling pathways in the regulation of cell survival. FASEB J. 2008, 22, 954–965. [Google Scholar] [CrossRef]
- Lai, C.F.; Chaudhary, L.; Fausto, A.; Halstead, L.R.; Ory, D.S.; Avioli, L.V.; Cheng, S.L. Erk is essential for growth, differentiation, integrin expression, and cell function in human osteoblastic cells. J. Biol. Chem. 2001, 276, 14443–14450. [Google Scholar]
- Matsuda, N.; Horikawa, M.; Watanabe, M.; Kitagawa, S.; Kudo, Y.; Takata, T. Possible involvement of extracellular signal-regulated kinases 1/2 in mitogenic response of periodontal ligament cells to enamel matrix derivative. Eur. J. Oral Sci. 2002, 110, 439–444. [Google Scholar] [CrossRef]
- Cheng, L.; Lin, Z.K.; Shu, R.; Liu, D.L.; Zhang, X.L.; Liu, B.; Wang, J.; Tian, L. Analogous effects of recombinant human full-length amelogenin expressed by Pichia pastoris yeast and enamel matrix derivative in vitro. Cell Prolif. 2012, 45, 456–465. [Google Scholar] [CrossRef]
- Ohyama, M.; Suzuki, N.; Yamaguchi, Y.; Maeno, M.; Otsuka, K.; Ito, K. Effect of enamel matrix derivative on the differentiation of C2C12 cells. J. Periodontol. 2002, 73, 543–550. [Google Scholar] [CrossRef]
- Guida, L.; Annunziata, M.; Carinci, F.; di Feo, A.; Passaro, I.; Oliva, A. In vitro biologic response of human bone marrow stromal cells to enamel matrix derivative. J. Periodontol. 2007, 78, 2190–2196. [Google Scholar] [CrossRef]
- Weinreb, M.; Shinar, D.; Rodan, G.A. Different pattern of alkaline phosphatase, osteopontin, and osteocalcin expression in developing rat bone visualized by in situ hybridization. J. Bone Miner. Res. 1990, 5, 831–842. [Google Scholar] [CrossRef]
- Aubin, J.E.; Liu, F.; Malaval, L.; Gupta, A.K. Osteoblast and chondroblast differentiation. Bone 1995, 17, 77S–83S. [Google Scholar] [CrossRef]
- Hägewald, S.; Pischon, N.; Jawor, P.; Bernimoulin, J.P.; Zimmermann, B. Effects of enamel matrix derivative on proliferation and differentiation of primary osteoblasts. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2004, 98, 243–249. [Google Scholar] [CrossRef]
- Blair, H.C.; Zaidi, M.; Schlesinger, P.H. Mechanisms balancing skeletal matrix synthesis and degradation. Biochem. J. 2002, 364, 329–341. [Google Scholar] [CrossRef]
- Nokhbehsaim, M.; Deschner, B.; Bourauel, C.; Reimann, S.; Winter, J.; Rath, B.; Jäger, A.; Jepsen, S.; Deschner, J. Interactions of enamel matrix derivative and biomechanical loading in periodontal regenerative healing. J. Periodontol. 2011, 82, 1725–1734. [Google Scholar] [CrossRef]
- Jue, S.S.; Lee, W.Y.; Kwon, Y.D.; Kim, Y.R.; Pae, A.; Lee, B. The effects of enamel matrix derivative on the proliferation and differentiation of human mesenchymal stem cells. Clin. Oral Implants Res. 2010, 21, 741–746. [Google Scholar] [CrossRef]
- Ikeda, T.; Nomura, S.; Yamaguchi, A.; Suda, T.; Yoshiki, S. In situ hybridization of bone matrix proteins in undecalcified adult rat bone sections. J. Histochem. Cytochem. 1992, 40, 1079–1088. [Google Scholar] [CrossRef]
- Iwata, T.; Morotome, Y.; Tanabe, T.; Fukae, M.; Ishikawa, I.; Oida, S. Noggin blocks osteoinductive activity of porcine enamel extracts. J. Dent. Res. 2002, 81, 387–391. [Google Scholar] [CrossRef]
- Keila, S.; Nemcovsky, C.E.; Moses, O.; Artzi, Z.; Weinreb, M. In vitro effects of enamel matrix proteins on rat bone marrow cells and gingival fibroblasts. J. Dent. Res. 2004, 83, 134–138. [Google Scholar] [CrossRef]
- Van den Dolder, J.; Vloon, A.P.; Jansen, J.A. The effect of Emdogain on the growth and differentiation of rat bone marrow cells. J. Periodontal. Res. 2006, 41, 471–476. [Google Scholar] [CrossRef]
- Huang, Y.C.; Tanimoto, K.; Tanne, Y.; Kamiya, T.; Kunimatsu, R.; Michida, M.; Yoshioka, M.; Yoshimi, Y.; Kato, Y.; Tanne, K. Effects of human full-length amelogenin on the proliferation of human mesenchymal stem cells derived from bone marrow. Cell Tissue Res. 2010, 342, 205–212. [Google Scholar] [CrossRef]
- Zeldich, E.; Koren, R.; Nemcovsky, C.; Weinreb, M. Enamel matrix derivative stimulates human gingival fibroblast proliferation via ERK. J. Dent. Res. 2007, 86, 41–46. [Google Scholar] [CrossRef]
- Miron, R.J.; Caluseru, O.M.; Guillemette, V.; Zhang, Y.; Gemperli, A.C.; Chandad, F.; Sculean, A. Influence of enamel matrix derivative on cells at different maturation stages of differentiation. PLoS One 2013, 8, e71008. [Google Scholar]
- Dean, D.D.; Lohmann, C.H.; Sylvia, V.L.; Cochran, D.L.; Liu, Y.; Boyan, B.D.; Schwartz, Z. Effect of porcine fetal enamel matrix derivative on chondrocyte proliferation, differentiation, and local factor production is dependent on cell maturation state. Cells Tissues Organs. 2002, 171, 117–127. [Google Scholar] [CrossRef]
- Miron, R.J.; Hedbom, E.; Ruggiero, S.; Bosshardt, D.D.; Zhang, Y.; Mauth, C.; Gemperli, A.C.; Iizuka, T.; Buser, D.; Sculean, A. Premature osteoblast clustering by enamel matrix proteins induces osteoblast differentiation through up-regulation of connexin 43 and N-cadherin. PLoS One 2011, 6, e23375. [Google Scholar] [CrossRef] [Green Version]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Katayama, N.; Kato, H.; Taguchi, Y.; Tanaka, A.; Umeda, M. The Effects of Synthetic Oligopeptide Derived from Enamel Matrix Derivative on Cell Proliferation and Osteoblastic Differentiation of Human Mesenchymal Stem Cells. Int. J. Mol. Sci. 2014, 15, 14026-14043. https://doi.org/10.3390/ijms150814026
Katayama N, Kato H, Taguchi Y, Tanaka A, Umeda M. The Effects of Synthetic Oligopeptide Derived from Enamel Matrix Derivative on Cell Proliferation and Osteoblastic Differentiation of Human Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2014; 15(8):14026-14043. https://doi.org/10.3390/ijms150814026
Chicago/Turabian StyleKatayama, Nobuhito, Hirohito Kato, Yoichiro Taguchi, Akio Tanaka, and Makoto Umeda. 2014. "The Effects of Synthetic Oligopeptide Derived from Enamel Matrix Derivative on Cell Proliferation and Osteoblastic Differentiation of Human Mesenchymal Stem Cells" International Journal of Molecular Sciences 15, no. 8: 14026-14043. https://doi.org/10.3390/ijms150814026