Effects of Pulsed Electromagnetic Field on Differentiation of HUES-17 Human Embryonic Stem Cell Line

Abstract
:1. Introduction
2. Results and Discussion
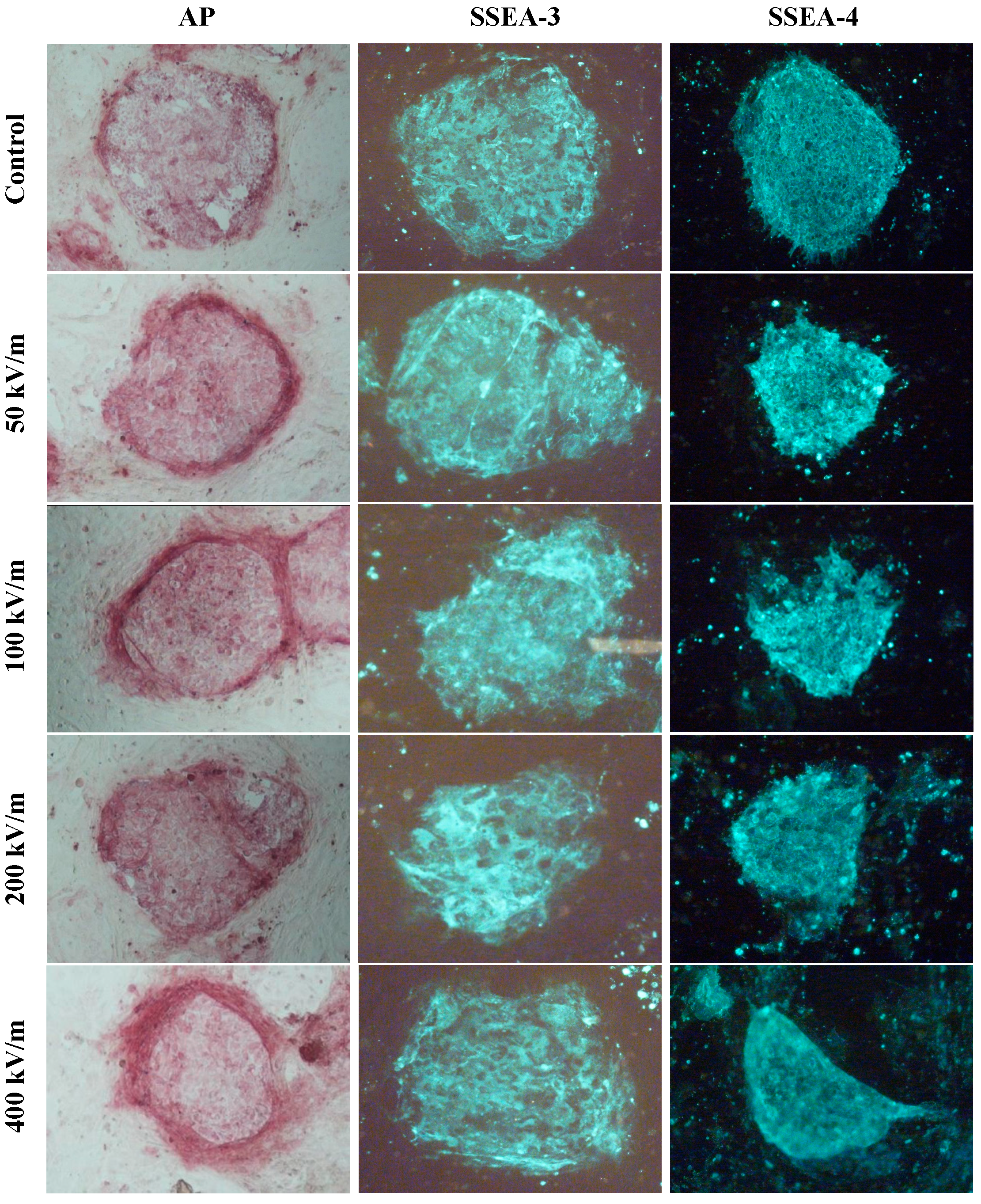
2.1. Effects of Pulsed Electromagnetic Field (PEMF) on Undifferentiated Markers in HUES-17 Cells

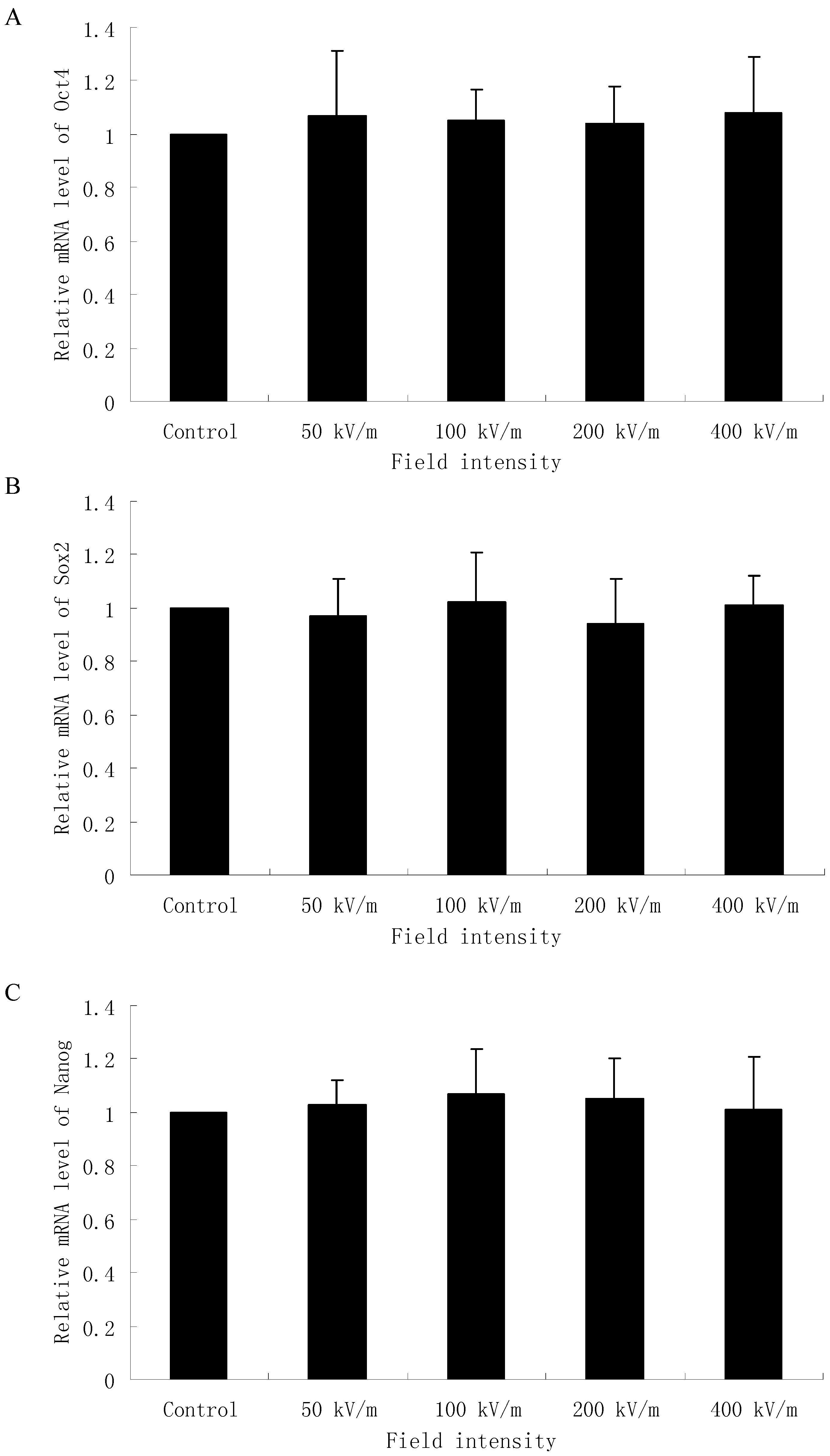
2.2. Effects of PEMF on mRNA Level of Oct4, Sox2 and Nanog in HUES-17 Cells

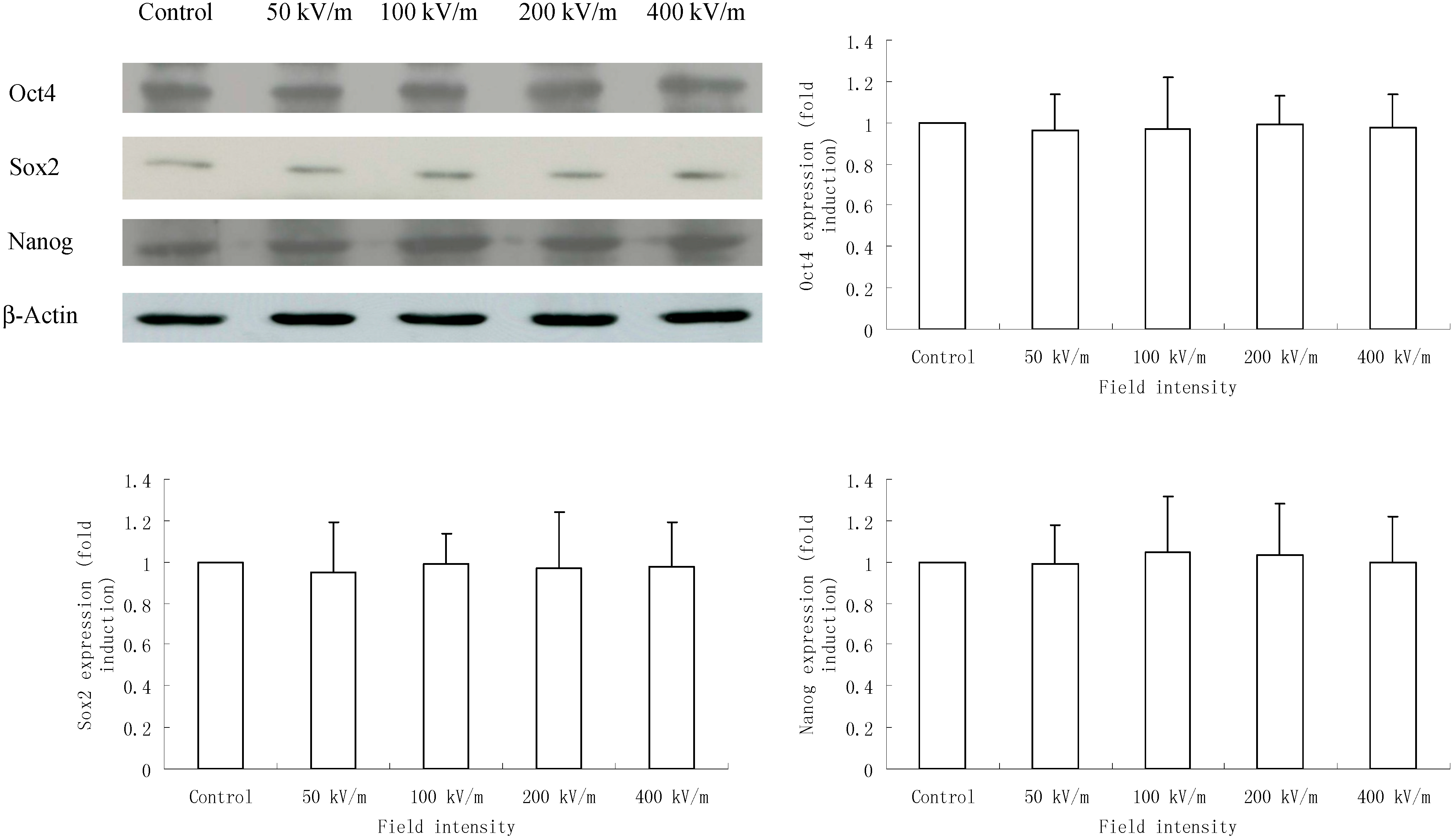
2.3. Effects of PEMF on Protein Level of Oct4, Sox2 and Nanog in HUES-17 Cells

2.4. Discussion
3. Experimental Section
3.1. Human Embryonic Stem Cell (hESC) Culture
3.2. PEMF Exposure
3.3. Alkaline Phosphatase (AP) Staining
3.4. Immunofluorescence Staining for Specific Markers
3.5. RNA Extraction and Real-Time Reverse Transcription-Polymerase Chain Reaction

{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer |
|---|---|
| Oct4 | Forward: AGAAGGATGTGGTCCGAGTGTG |
| Reverse: CCACCCTTTGTGTTCCCAATTCC | |
| Sox2 | Forward: CCCCCGGCGGCAATAGCA |
| Reverse: TCGGCGCCGGGGAGATACAT | |
| Nanog | Forward: TGAACCTCAGCTACAAACAGGTG |
| Reverse: AACTGCATGCAGGACTGCAGAG | |
| GAPDH | Forward: AGGTCGGAGTCAACGGATTTGG |
| Reverse: AGGCTGTTGTCATACTTCTCATGG |
3.6. Western Blotting
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bojjawar, T.; Jalari, M.; Aamodt, E.; Ware, M.F.; Haynie, D.T. Effect of electromagnetic nanopulses on C. elegans fertility. Bioelectromagnetics 2006, 27, 515–520. [Google Scholar] [CrossRef]
- Saito, K.; Suzuki, H.; Suzuki, K. Teratogenic effects of static magnetic field on mouse fetuses. Reprod. Toxicol. 2006, 22, 118–124. [Google Scholar] [CrossRef]
- Lahijani, M.S.; Sajadi, K. Development of preincubated chicken eggs following exposure to 50 Hz electromagnetic fields with 1.33–7.32 mT flux densities. Indian J. Exp. Biol. 2004, 42, 858–865. [Google Scholar]
- Beraldi, R.; Sciamanna, I.; Mangiacasale, R.; Lorenzini, R.; Spadafora, C. Mouse early embryos obtained by natural breeding or in vitro fertilization display a differential sensitivity to extremely low-frequency electromagnetic fields. Mutat. Res. 2003, 538, 163–170. [Google Scholar] [CrossRef]
- Lahijani, M.S.; Ghafoori, M. Teratogenic effects of sinusoidal extremely low frequency electromagnetic fields on morphology of 24 hour chick embryos. Indian J. Exp. Biol. 2000, 38, 692–699. [Google Scholar]
- Yang, M.J.; Liu, J.Y.; Wang, Y.F.; Lang, H.Y.; Miao, X.; Zhang, L.Y.; Zeng, L.H.; Guo, G.Z. Effects of electromagnetic pulse on polydactyly of mouse fetuses. Theriogenology 2013, 80, 18–23. [Google Scholar] [CrossRef]
- Ahuja, Y.R.; Vijayalakshmi, V.; Polasa, K. Stem cell test: A practical tool in toxicogenomics. Toxicology 2007, 231, 1–10. [Google Scholar] [CrossRef]
- Maher, E.R.; Afnan, M.; Barratt, C.L. Epigenetic risks related to assisted reproductive technologies: Epigenetics, imprinting, ART and icebergs? Hum. Reprod. 2003, 18, 2508–2511. [Google Scholar] [CrossRef]
- Jacob, S.; Moley, K.H. Gametes and embryo epigenetic reprogramming affect developmental outcome: Implication for assisted reproductive technologies. Pediatr. Res. 2005, 58, 437–446. [Google Scholar] [CrossRef]
- Malagoli, D.; Gobba, F.; Ottaviani, E. Effects of 50-Hz magnetic fields on the signalling pathways of fMLP-induced shape changes in invertebrate immunocytes: The activation of an alternative “stress pathway”. Biochim. Biophys. Acta 2003, 1620, 185–190. [Google Scholar] [CrossRef]
- Czyz, J.; Nikolova, T.; Schuderer, J.; Kuster, N.; Wobus, A.M. Non-thermal effects of power-line magnetic fields (50 Hz) on gene expression levels of pluripotent embryonic stem cells-the role of tumour suppressor p53. Mutat. Res. 2004, 557, 63–74. [Google Scholar] [CrossRef]
- Czyz, J.; Guan, K.; Zeng, Q.; Nikolova, T.; Meister, A.; Schonborn, F.; Schuderer, J.; Kuster, N.; Wobus, A.M. High frequency electromagnetic fields (GSM signals) affect gene expression levels in tumor suppressor p53-deficient embryonic stem cells. Bioelectromagnetics 2004, 25, 296–307. [Google Scholar] [CrossRef]
- Chang, S.H.; Hsiao, Y.W.; Lin, H.Y. Low-frequency electromagnetic field exposure accelerates chondrocytic phenotype expression on chitosan substrate. Orthopedics 2011, 34, 20. [Google Scholar]
- Levin, M. Bioelectromagnetics in morphogenesis. Bioelectromagnetics 2003, 24, 295–315. [Google Scholar] [CrossRef]
- Ahuja, Y.R.; Bhargava, S.C.; Ratnakar, K.S. Electric and magnetic fields in stem cell research. Electromagn. Biol. Med. 2005, 24, 121–134. [Google Scholar] [CrossRef]
- Pagliara, P.; Lanubile, R.; Dwikat, M.; Abbro, L.; Dini, L. Differentiation of monocytic U937 cells under static magnetic field exposure. Eur. J. Histochem. 2005, 49, 75–86. [Google Scholar]
- Collard, J.F.; Mertens, B.; Hinsenkamp, M. In vitro study of the effects of ELF electric fields on gene expression in human epidermal cells. Bioelectromagnetics 2011, 32, 28–36. [Google Scholar] [CrossRef]
- Collard, J.F.; Lazar, C.; Nowe, A.; Hinsenkamp, M. Statistical validation of the acceleration of the differentiation at the expense of the proliferation in human epidermal cells exposed to extremely low frequency electric fields. Prog. Biophys. Mol. Biol. 2013, 111, 37–45. [Google Scholar] [CrossRef]
- Pesce, M.; Patruno, A.; Speranza, L.; Reale, M. Extremely low frequency electromagnetic field and wound healing: Implication of cytokines as biological mediators. Eur. Cytokine Netw. 2013, 24, 1–10. [Google Scholar]
- Serena, E.; Figallo, E.; Tandon, N.; Cannizzaro, C.; Gerecht, S.; Elvassore, N.; Vunjak-Novakovic, G. Electrical stimulation of human embryonic stem cells: Cardiac differentiation and the generation of reactive oxygen species. Exp. Cell. Res. 2009, 315, 3611–3619. [Google Scholar] [CrossRef]
- Ventura, C.; Maioli, M.; Asara, Y.; Santoni, D.; Mesirca, P.; Remondini, D.; Bersani, F. Turning on stem cell cardiogenesis with extremely low frequency magnetic fields. FASEB J. 2005, 19, 155–157. [Google Scholar]
- Sauer, H.; Rahimi, G.; Hescheler, J.; Wartenberg, M. Effects of electrical fields on cardiomyocyte differentiation of embryonic stem cells. J. Cell. Biochem. 1999, 75, 710–723. [Google Scholar] [CrossRef]
- Boyer, L.A.; Lee, T.I.; Cole, M.F.; Johnstone, S.E.; Levine, S.S.; Zucker, J.P.; Guenther, M.G.; Kumar, R.M.; Murray, H.L.; Jenner, R.G. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell 2005, 122, 947–956. [Google Scholar] [CrossRef]
- Niwa, H.; Miyazaki, J.; Smith, A.G. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cells. Nat. Genet. 2000, 24, 372–376. [Google Scholar] [CrossRef]
- Rizzino, A. Sox2 and Oct-3/4: A versatile pair of master regulators that orchestrate the self-renewal and pluripotency of embryonic stem cells. Wiley Interdiscip. Rev. Syst. Biol. Med. 2009, 1, 228–236. [Google Scholar] [CrossRef]
- Cowan, C.A.; Klimanskaya, I.; McMahon, J.; Atienza, J.; Witmyer, J.; Zucker, J.P.; Wang, S.; Morton, C.C.; McMahon, A.P.; Powers, D.; et al. Derivation of embryonic stem-cell lines from human blastocysts. N. Engl. J. Med. 2004, 350, 1353–1356. [Google Scholar] [CrossRef]
- Cheng, L.; Qiu, R.Z.; Deng, H.; Zhang, Y.A.; Jin, Y.; Li, L. Ethics: China already has clear stem-cell guidelines. Nature 2006, 440, 992. [Google Scholar]
- Li, K.C.; Hui, Y.P.; Ma, S.R.; Ding, G.R.; Guo, Y.; Liu, J.Y.; Li, Y.R.; Guo, G.Z. Inhibition of bone formation by high intensity pulsed electromagnetic field in mc3t3-e1 cells. Prog. Electromagn. Res. 2011, 139–153. [Google Scholar]
- Xiao, L.; Yuan, X.; Sharkis, S.J. Activin a maintains self-renewal and regulates fibroblast growth factor, Wnt, and bone morphogenic protein pathways in human embryonic stem cells. Stem Cells 2006, 24, 1476–1486. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, W.; Chen, G.; Cheng, L.; Liao, J.; Jia, N.; Gao, Y.; Dai, H.; Yuan, J.; Cheng, L.; et al. Combinatorial signals of activin/nodal and bone morphogenic protein regulate the early lineage segregation of human embryonic stem cells. J. Biol. Chem. 2008, 283, 24991–25002. [Google Scholar] [CrossRef]
- Guo, Y.; Liu, S.; Wang, P.; Zhao, S.; Wang, F.; Bing, L.; Zhang, Y.; Ling, E.A.; Gao, J.; Hao, A. Expression profile of embryonic stem cell-associated genes Oct4, Sox2 and Nanog in human gliomas. Histopathology 2011, 59, 763–775. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, Y.-L.; Ma, S.-R.; Peng, T.; Teng, Z.-H.; Liang, X.-Y.; Guo, G.-Z.; Zhang, H.-F.; Li, K.-C. Effects of Pulsed Electromagnetic Field on Differentiation of HUES-17 Human Embryonic Stem Cell Line. Int. J. Mol. Sci. 2014, 15, 14180-14190. https://doi.org/10.3390/ijms150814180
Wu Y-L, Ma S-R, Peng T, Teng Z-H, Liang X-Y, Guo G-Z, Zhang H-F, Li K-C. Effects of Pulsed Electromagnetic Field on Differentiation of HUES-17 Human Embryonic Stem Cell Line. International Journal of Molecular Sciences. 2014; 15(8):14180-14190. https://doi.org/10.3390/ijms150814180
Chicago/Turabian StyleWu, Yi-Lin, Shi-Rong Ma, Tao Peng, Zeng-Hui Teng, Xiang-Yan Liang, Guo-Zhen Guo, Hai-Feng Zhang, and Kang-Chu Li. 2014. "Effects of Pulsed Electromagnetic Field on Differentiation of HUES-17 Human Embryonic Stem Cell Line" International Journal of Molecular Sciences 15, no. 8: 14180-14190. https://doi.org/10.3390/ijms150814180