Immunoproteome of Aspergillus fumigatus Using Sera of Patients with Invasive Aspergillosis

Abstract
:1. Introduction
2. Results and Discussion
2.1. Western Immunoblots and Antigenic Proteins Identified


{kind=link}
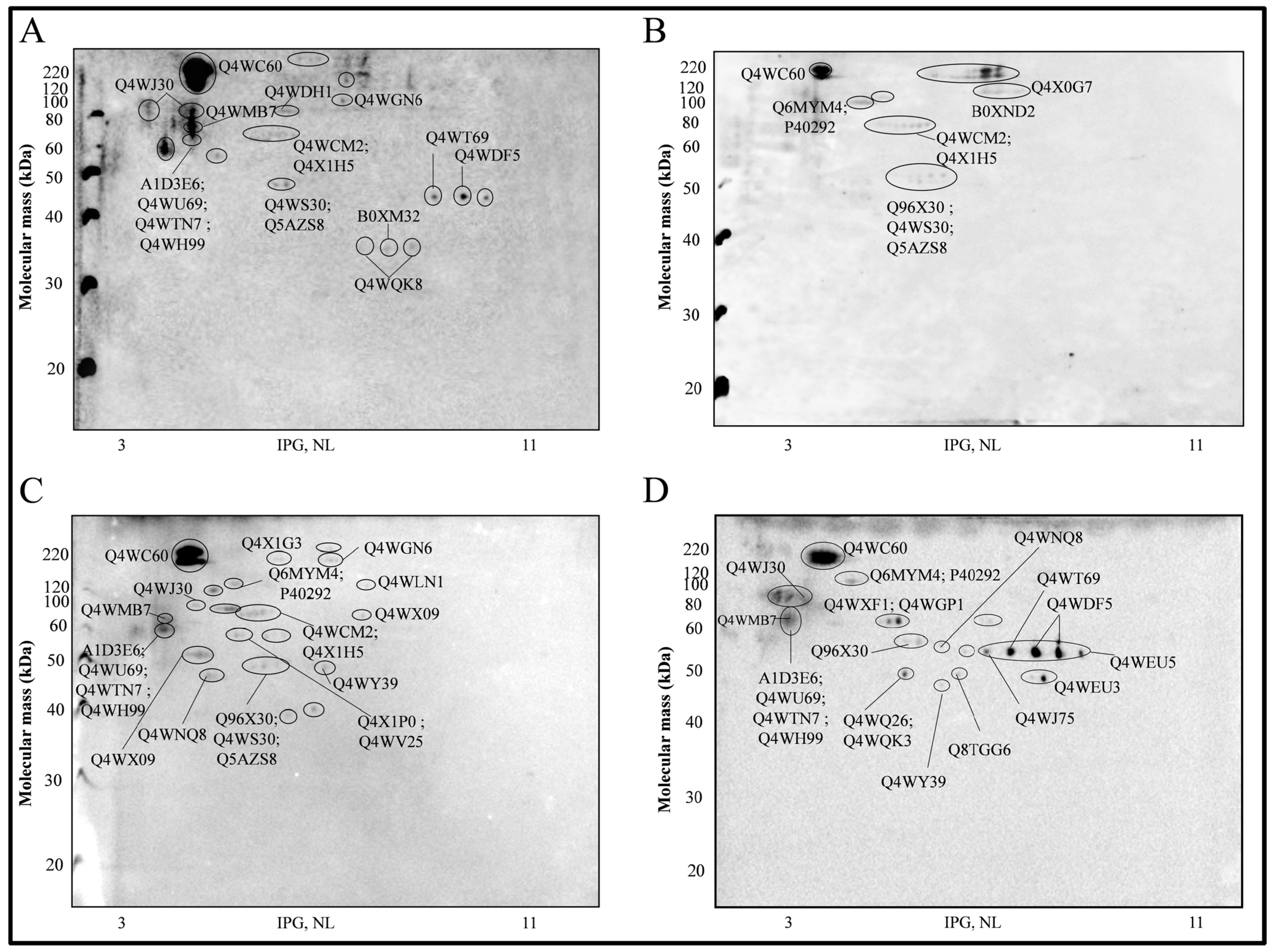{kind=link}
| Spot | Cov. (95%) | Accession Number | EC Number | Theorical Mr/PI | Orf | Organism | Name | Function | Peptides | Conf. | Sequence |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 4.60 | Q4WC60 | 3.2.1.21 | 58.148/4.93 | AFUA_8G05610 | A. fumigatus | Probable β-glucosidase btgE | Degradation of cellulosic biomass | 2 | 99 | EPGQFGVER |
| 99 | VYSTDCNSLEYIGEAAR | ||||||||||
| 8 | 5.02 | Q4X1G3 | 6.3.5.5 | 129.214/5.87 | AFUA_2G10070 | A. fumigatus | Carbamoyl-phosphate synthase, large subunit | Nitrogen compound metabolic process | 4 | 99 | FAESVGYPVLVR |
| 99 | QIALLVGSTEDDVR | ||||||||||
| 99 | AAESVGYPIIVR | ||||||||||
| 99 | LADEVYYLPVTPEYVTHVIER | ||||||||||
| 11, 12, 13, 14, 15 | 8.94 | Q4X0G7 | - | 93.198/6.51 | AFUA_2G13530 | A. fumigatus | Translation elongation factor EF-2 subunit, putative | Translation elongation factor activity | 6 | 99 | GHVYSEEQRPGTPLFNVK |
| 99 | ALGDVQVYPDR | ||||||||||
| 99 | AYLPVNESFGFNGDLR | ||||||||||
| 99 | DLEEDHAGVPLR | ||||||||||
| 99 | VNFTIEEIR | ||||||||||
| 99 | FSVSPVVQR | ||||||||||
| 13, 15 | 5.45 | B0XND2 | - | 81.445/5.74 | AFUB_004530 | A. fumigatus | Polyadenylate-binding protein | RNA-binding | 3 | 99 | NIDQEVTDEEFR |
| 98 | NLTDDVDDEKLR | ||||||||||
| 99 | SLGYAYVNYNNTADGER | ||||||||||
| 24 | 9.66 | Q6MYM4 | - | 80.04/5.08 | AfA5C5.047 | A. fumigatus | Heat shock protein Hsp88, putative | Response to stress | 5 | 99 | FIAGPIVQR |
| 99 | KNELESTIYELR | ||||||||||
| 99 | LDLPGPEEKPR | ||||||||||
| 99 | STPTLVGFGTR | ||||||||||
| 99 | TLSFTLNQDEAIAR | ||||||||||
| 14.16 | P40292 | - | 80.64/4.94 | AFUA_5G04170 | A. fumigatus | Heat shock protein 90-Heat shock protein hsp1 (Asp f 12) | Promotes maturation, structural maintenance and proper regulation of specific target proteins involved for instance in cell cycle control and signal transduction | 8 | 99 | ADLINNLGTIAR | |
| 99 | GVVDSEDLPLNLSR | ||||||||||
| 99 | HFSVEGQLEFR | ||||||||||
| 99 | IILHLKDEQTDYLNESR | ||||||||||
| 99 | RAPFDLFETK | ||||||||||
| 99 | TGQFGWSANMER | ||||||||||
| 98 | LGIHEDAQNR | ||||||||||
| 96 | DFELEETEEEKAER | ||||||||||
| 26 | 7.83 | Q4WDH1 | 6.3.4.13 | 86.42/5.31 | AFUA_6G04730 | A. fumigatus | Bifunctional purine biosynthetic protein Ade1, putative | de novo IMP biosynthetic process, purine nucleobase biosynthetic process | 4 | 99 | EGEVVYQVGELKPR |
| 99 | GLAHITGGGLVENVPR | ||||||||||
| 99 | HNIPTAAYQNFYEYEPAR | ||||||||||
| 99 | VIASTATASTLEEALR | ||||||||||
| 35, 36, 37 | 8.77 | Q4WLN1 | 4.2.1.3 | 85.53/6.26 | AFUA_6G12930 | A. fumigatus | Mitochondrial aconitate hydratase, putative | Mitochondrial genome maintenance | 6 | 99 | FTGEYDAVPATAR |
| 99 | HLGGLAIITR | ||||||||||
| 99 | LQRPLTYAEK | ||||||||||
| 99 | QHIGDFAR | ||||||||||
| 99 | SLFTVTPGSEQIR | ||||||||||
| 99 | WVVIGDWNYGEGSSR | ||||||||||
| 46, 47 | 12.66 | Q4WJ30 | - | 69.66/5.08 | AFUA_1G07440 | A. fumigatus | Molecular chaperone Hsp70 | ATP and nucleotide binding; protein refolding | 6 | 99 | ATAGDTHLGGEDFDNR |
| 99 | DAGLIAGLNVLR | ||||||||||
| 99 | FELTGIPPAPR | ||||||||||
| 99 | SSVHEIVLVGGSTR | ||||||||||
| 99 | TTPSFVAFTDTER | ||||||||||
| 96 | LVNHFVNEFKR | ||||||||||
| 59 | 11.37 | Q4WMB7 | - | 53.56/4.58 | AFUA_6G10470 | A. fumigatus | Zinc finger protein ZPR1 | Cellular response to starvation; regulation of mitotic cell cycle | 4 | 99 | DIILESFECEHCGHR |
| 99 | FTTVEGLLTEIR | ||||||||||
| 99 | GESQLTTVEGVIQR | ||||||||||
| 99 | YTLDVENEEDFQR | ||||||||||
| 61, 62 | 7.98 | Q4WCM2 | - | 66.97/5.30 | AFUA_8G03930 | A. fumigatus | Hsp70 chaperone (HscA), putative | Protein refolding; ATP-binding | 4 | 99 | AVITVPAYFNDNQR |
| 99 | DAGAIAGLNVLR | ||||||||||
| 99 | QQLESYISR | ||||||||||
| 99 | SQVDEIVLVGGSTR | ||||||||||
| 63, 64, 65 | 14.01 | Q4X1H5 | - | 74.46/6.02 | AFUA_2G09960 | A. fumigatus | Mitochondrial Hsp70 chaperone (Ssc70), putative | Protein refolding; protein targeting to mitochondrion | 7 | 99 | DAGQIAGLNVLR |
| 99 | IVQHTNGDAWVEAR | ||||||||||
| 99 | LLGNFQLVGIPPAHR | ||||||||||
| 99 | NAVVTVPAYFNDSQR | ||||||||||
| 99 | SQLESLVEPLINR | ||||||||||
| 99 | TTPSVVAFAQDGER | ||||||||||
| 98 | YSPSQIGGFILQK | ||||||||||
| 71 | 10.47 | A1D3E6 | - | 46.64/4.69 | NFIA_016350 | A. fischerianus | Protein phosphatase 2C, putative | Protein dephosphorylation | 3 | 99 | ISAAGGFVDFGR |
| 99 | NQFEETPDNYDLENDR | ||||||||||
| 99 | VANGDGPCAPPEYAEFR | ||||||||||
| 8.35 | Q4WU69 | - | 54.25/4.50 | AFUA_5G07390 | A. fumigatus | 60S ribosome biogenesis protein Sqt1, putative | Structural constituent of ribosome | 3 | 99 | GEYVVTAGLDGR | |
| 99 | VEFLQTNLAALASR | ||||||||||
| 96 | DERPVLPQSYESNPQPK | ||||||||||
| 8.98 | Q4WTN7 | - | 48.34/4.43 | AFUA_5G05540 | A. fumigatus | Nucleosome assembly protein Nap1, putative | Nucleosome assembly | 3 | 99 | EESLDHATAASLFAR | |
| 99 | SSGYIESLPAPVR | ||||||||||
| 99 | MEYLDRPGFR | ||||||||||
| 2.71 | Q4WH99 | 5.3.4.1 | 56.19/4.58 | AFUA_2G06150 | A. fumigatus | Protein disulfide isomerase Pdi1, putative | Cell redox homeostasis; glycerol ether metabolic process | 1 | 99 | AANDVFTSFAESQR | |
| 73, 74 | 7.11 | Q4WXF1 | 5.4.2.1 | 57.45/5.44 | AFUA_3G09290 | A. fumigatus | Phosphoglycerate mutase, 2,3-bisphosphoglycerate-independent | Glucose catabolic process | 3 | 99 | VQDNDTLFFFNYR |
| 99 | EIGIGEIATVVGR | ||||||||||
| 99 | EITQLLGDYDR | ||||||||||
| 15.26 | Q4WGP1 | 2.3.1.12 | 52.03/6.26 | AFUA_7G05720 | A. fumigatus | Pyruvate dehydrogenase complex, dihydrolipoamide acetyltransferase component, putative | Acetyl-CoA biosynthetic process from pyruvate | 5 | 99 | FTAVINPPQAAILAVGTTR | |
| 99 | LQPSLDREPNISPAAK | ||||||||||
| 99 | NVHSLGLSSISNQIK | ||||||||||
| 99 | VPAVNSSWR | ||||||||||
| 99 | ENPHFFVSTTLSVTK | ||||||||||
| 114, 117 | 22.60 | Q96X30 | 4.2.1.11 | 47.31/5.39 | AFUA_6G06770 | A. fumigatus | Enolase (Asp f 22) | Glycolysis;regulation of vacuole fusion, non-autophagic | 6 | 99 | AIVPSGASTGQHEAHELR |
| 99 | DSYADNWGVMVSHR | ||||||||||
| 99 | GNPTVEVDVVTETGLHR | ||||||||||
| 99 | GVPLYAHISDLAGTK | ||||||||||
| 99 | SGETEDVTIADIAVGLR | ||||||||||
| 99 | TSDFQIVGDDLTVTNPGR | ||||||||||
| 119, 120 | 12.75 | Q4WS30 | 3.4.24.64 | 53.27/5.90 | AFUA_1G14200 | A. fumigatus | Mitochondrial processing peptidase beta subunit, putative | Metalloendopeptidase activity | 5 | 99 | ASILLSLDGTTAVAEDIGR |
| 99 | ITEKDVMDFANR | ||||||||||
| 99 | LCYNVSAAEVER | ||||||||||
| 99 | LNDLVHFALR | ||||||||||
| 99 | TPEFIGSEIR | ||||||||||
| 120 | 4.70 | Q5AZS8 | - | 49.75/9.88 | AN6202.2 | A. nidulans | RL3_NEUCR 60S ribosomal protein L3 | Structural constituent of ribosome | 1 | 99 | DEMIDVIAVTKGHGFQGVTSR |
| 131 | 25.66 | Q4WT69 | 2.7.2.3 | 44.76/6.31 | AFUA_1G10350 | A. fumigatus | Phosphoglycerate kinase | Phosphoglycerate kinase activity | 8 | 99 | ALESPSRPFLAILGGSK |
| 99 | ASGGQVILLENLR | ||||||||||
| 99 | FHPEEEGSYKDEEGK | ||||||||||
| 99 | FHPEEEGSYKDEEGKK | ||||||||||
| 99 | GLTALGDIYINDAFGTAHR | ||||||||||
| 99 | IGNSLFDEAGSK | ||||||||||
| 99 | IVLPVDYITADKFSADAK | ||||||||||
| 99 | YSLKPVVPELEK | ||||||||||
| 132 | 14.52 | Q4WDF5 | - | 54.18/7.18 | AFUA_6G04570 | A. fumigatus | Translation elongation factor eEF-1 subunit gamma, putative | Translation elongation factor activity | 7 | 99 | AVVPSPVFAEEAIK |
| 99 | EYPHVDGHVFK | ||||||||||
| 99 | HLTANTYLVGER | ||||||||||
| 99 | ITLADYFGASLLTR | ||||||||||
| 99 | TKQDYAAILR | ||||||||||
| 98 | QDYAAILR | ||||||||||
| 97 | LYGLPENGR | ||||||||||
| 141 | 5.65 | Q4WEU3 | 1.10.2.2 | 48.09/8.89 | AFUA_5G04210 | A. fumigatus | Ubiquinol-cytochrome C reductase complex core protein 2, putative | Ubiquinolcytochrome-C reductase activity | 2 | 99 | ATQGFSQVR SNIAIVGSGSSTAEVSR |
| 178 | 2.99 | B0XM32 | - | 56.40/6.84 | AFUB_000800 | A. fumigatus | Cytochrome P450 | Oxidoreductase activity, acting on paired donors, with incorporation or reduction of molecular oxygen | 1 | 99 | LLSDQFAGFPSVNSR |
| 176, 180 | 22.15 | Q4WQK8 | - | 34.99/6.06 | AFUA_4G13170 | A. fumigatus | G-protein comlpex beta subunit CpcB | Cell signaling | 5 | 99 | VDELKPEFIEK |
| 99 | HLYSLHAGDEIHALVFSPNR | ||||||||||
| 99 | LWELATGETTR | ||||||||||
| 99 | TFVGHTSDVLSVSFSADNR | ||||||||||
| 99 | TLIIWNLTR | ||||||||||
| A | 3.29 | Q4WGN6 | 3.6.3.- | 117.77/5.84 | AFUA_7G05660 | A. fumigatus | Translation elongation factor eEF-3 | Translation elongation factor activity | 3 | 99 | FLDNVIQHVVHYER |
| 99 | TFEGGVVIITHSR | ||||||||||
| 97 | LEEFGFLR | ||||||||||
| B | 5.87 | Q4WX09 | - | 71.15/6.50 | AFUA_3G07810 | A. fumigatus | Succinate dehydrogenase subunit Sdh1, putative | Eectron transport chain; tricarboxylic acid cycle | 3 | 99 | AHHTVLATGGYGR |
| 99 | KPHGEINLGYR | ||||||||||
| 99 | GIIAYNQEDGTLHR | ||||||||||
| C | 5.45 | Q4X1P0 | - | 61.95/5.53 | AFUA_2G09290 | A. fumigatus | Antigenic mitochondrial protein HSP60, putative | Cellular response to temperature stimulus; protein refolding | 2 | 99 | AITLQDKFENLGAR |
| 99 | ISAVQDIIPALEASTTLR | ||||||||||
| D | 20.81 | Q4WV25 | 3.6.3.14 | 55.62/5.30 | AFUA_5G10550 | A. fumigatus | ATP synthase subunit beta | ATP catabolic process | 9 | 99 | DTGAPIKIPVGPGTLGR |
| 99 | FTQAGSEVSALLGR | ||||||||||
| 99 | IPVGPGTLGR | ||||||||||
| 99 | IVGEEHYAVATR | ||||||||||
| 99 | IVNVTGDPIDER | ||||||||||
| 99 | LVLEVSQHLGENVVR | ||||||||||
| 99 | VALTGLTIAEYFR | ||||||||||
| 99 | VALVFGQMNEPPGAR | ||||||||||
| 99 | VVDLLAPYAR | ||||||||||
| E | 23.15 | Q4WX43 | 3.6.4.13 | 45.78/5.05 | AFUA_3G08160 | A. fumigatus | ATP-dependent RNA helicase eIF4A | Complex eIF4F subunit-involved in the “cap” recognition; necessary to mRNA binding to ribossome | 7 | 99 | ALQEGPQVVVGTPGR |
| 99 | DFTVSAMHGDMEQAQR | ||||||||||
| 99 | GCQALILAPTR | ||||||||||
| 99 | GVAINFVTADDVR | ||||||||||
| 99 | GVYAYGFERPSAIQQR | ||||||||||
| 99 | MFILDEADEMLSR | ||||||||||
| 99 | VLIATDLLAR | ||||||||||
| F | 14.41 | Q4WNQ8 | - | 49.37/5.79 | AFUA_4G06620 | A. fumigatus | Glutamate dehydrogenase -Glu/Leu/Phe/Val dehydrogenase | Oxidoreductase activity | 5 | 99 | AANAGGVAVSGLEMAQNSAR |
| 99 | FLGFEQIFK | ||||||||||
| 99 | VVWEDDNHQVQINR | ||||||||||
| 99 | YIEGARPWVHVGK | ||||||||||
| 99 | EIGFLFGQYR | ||||||||||
| G | 18.06 | Q4WY39 | 4.1.2.13 | 39.79/5.55 | AFUA_3G11690 | A. fumigatus | Fructose-bisphosphate aldolase, class II | Fructose-bisphosphate aldolase activity; zinc ion binding | 6 | 99 | ASIAGSIAAAHYIR |
| 99 | KSGVIVGDDVLR | ||||||||||
| 99 | LFEYAQEK | ||||||||||
| 99 | RVQVALEDFNTAGQL | ||||||||||
| 99 | SGVIVGDDVLR | ||||||||||
| 99 | VNLDTDMQYAYMSGVR | ||||||||||
| I | 6.37 | Q4WQ26 | - | 42.35/5.68 | AFUA_4G11330 | A. fumigatus | Aha1 domain family | ATPase activator activity-Response to stress | 3 | 99 | QNWDVYYVR |
| VAVNTTTVTASDEFR | |||||||||||
| QNWDVYYVR | |||||||||||
| 18.71 | Q4WQK3 | 6.3.1.2 | 39.90/5.48 | AFUA_4G13120 | A. fumigatus | Glutamine synthetase | Glutamate-ammonia ligase activity-Glutamine biosynthetic process | 5 | 99 | DIVEAHYR | |
| FSYGVADR | |||||||||||
| GDWNGAGLHTNVSTAATR | |||||||||||
| GGFPGAQGPYYCGVGTGK | |||||||||||
| HNEHIAVYGEGNEER | |||||||||||
| K | 23.86 | Q8TGG6 | - | 48.29/6.69 | AfA14E5.05 | A. fumigatus | Elongation factor Tu | Translation elongation factor activity-Protein biosynthesis | 7 | 99 | AGDNSGLLLR |
| GITISTAHIEFSTDSR | |||||||||||
| GLANFLEYGAIDKAPEER | |||||||||||
| HYAHVDCPGHADYIK | |||||||||||
| TADEAADLSFPDGDQSR | |||||||||||
| THHPVAAEAGQR | |||||||||||
| TKPHVNIGTIGHVDHGK | |||||||||||
| L | 4.44 | Q4WJ75 | 1.2.4.1 | 41.48/6.36 | AFUA_1G06960 | A. fumigatus | Pyruvate dehydrogenase E1 component subunit alpha | Pyruvate dehydrogenase (acetyl-transferring) activity-Glycolytic process | 2 | 99 | ILFEDIYVR |
| SIIGELLGR | |||||||||||
| N | 14.76 | Q4WEU5 | - | 52.11/8.69 | AFUA_5G04230 | A. fumigatus | Citrate synthase | Citrate (Si)-synthase activity - Tricarboxylic acid cycle/Cellular carbohydrate metabolic process | 4 | 99 | CLVWEGSVLDSEEGIR |
| FIEELIDR | |||||||||||
| ALGAPIERPK | |||||||||||
| ALGVLPQLIIDR | |||||||||||
| DLSAEWAAR | |||||||||||
| FIEELIDR | |||||||||||
| VIGEVTLDQAYGGAR | |||||||||||
| O | 5.02 | Q4WWD5 | 3.-.-.- | 53.02/5.47 | AFUA_3G05450 | A. fumigatus | Glutamate carboxypeptidase, putative | Carboxypeptidase, Hydrolase Protease | 2 | 99 | EHLDLPPVVIAR |
| QVDELSNSFIDR | |||||||||||
| P | 8.85 | Q4WYW4 | 1.1.1.86 | 56.35/9.32 | AFUA_3G14490 | A. fumigatus | Ketol-acid reductoisomerase | Ketol-acid reductoisomerase activity-branched-chain amino acid biosynthetic process | 3 | 99 | DQGLNVIVGVR |
| EVYSDLYGER | |||||||||||
| TLYFSHGFSPVFK |

| Spot | Name | Proven-Hospital 1 | Proven-Hospital 2 | Probable | Other Mycoses | Control |
|---|---|---|---|---|---|---|
| 1 | Probable β-glucosidase btgE | X | X | X | X | X |
| 8 | Carbamoyl-phosphate synthase, large subunit | X | ||||
| 11, 12, 13, 14, 15 | Translation elongation factor EF-2 subunit, putative | X | ||||
| 13, 15 | Polyadenylate-binding protein | X | ||||
| 24 | Heat shock protein Hsp88, putative | X | X | X | X | |
| Heat shock protein 90–Heat shock protein hsp1 (Asp f 12) | X | X | X | |||
| 26 | Bifunctional purine biosynthetic protein Ade1, putative | X | ||||
| 35, 36, 37 | Mitochondrial aconitate hydratase, putative | X | ||||
| 46, 47 | Molecular chaperone Hsp70 | X | X | X | X | |
| 59 | Zinc finger protein ZPR1 | X | X | X | ||
| 61, 62 | Hsp70 chaperone (HscA), putative | X | X | X | ||
| 63, 64, 65 | Mitochondrial Hsp70 chaperone (Ssc70), putative | X | X | X | ||
| 71 | Protein phosphatase 2C, putative | X | X | X | X | |
| 60S ribosome biogenesis protein Sqt1, putative | X | X | X | X | ||
| Nucleosome assembly protein Nap1, putative | X | X | X | X | ||
| Protein disulfide isomerase Pdi1, putative | X | X | X | X | ||
| 73, 74 | Phosphoglycerate mutase, 2,3-bisphosphoglycerate-independent | X | ||||
| Pyruvate dehydrogenase complex, dihydrolipoamide acetyltransferase component, putative | X | |||||
| 114, 117 | Enolase (Asp f 22) | X | X | X | ||
| 119, 120 | Mitochondrial processing peptidase β subunit, putative | X | X | X | ||
| 120 | RL3_NEUCR 60S ribosomal protein L3 | X | X | X | ||
| 131 | Phosphoglycerate kinase | X | X | X | ||
| 132,133 | Translation elongation factor eEF-1 subunit γ, putative | X | X | X | ||
| 140, 141 | Ubiquinol-cytochrome C reductase complex core protein 2, putative | X | ||||
| 178 | Cytochrome P450 | X | ||||
| 176, 180 | G-protein comlpex beta subunit CpcB | X | ||||
| A | Translation elongation factor eEF-3 | X | X | |||
| B | Succinate dehydrogenase subunit Sdh1, putative | X | ||||
| C | Antigenic mitochondrial protein HSP60, putative | X | ||||
| D | ATP synthase subunit β | X | ||||
| E | ATP-dependent RNA helicase eIF4A | X | ||||
| F | Glutamate dehydrogenase—Glu/Leu/Phe/Val dehydrogenase | X | X | |||
| G | Fructose-bisphosphate aldolase, class II | X | X | |||
| I | Aha1 domain family | X | ||||
| I | Glutamine synthetase | X | ||||
| K | Elongation factor Tu | X | ||||
| L | Pyruvate dehydrogenase E1 component subunit α | X | ||||
| N | Citrate synthase | X | ||||
| O | Glutamate carboxypeptidase, putative | X | ||||
| P | Ketol-acid reductoisomerase | X |
2.2. BLAST Analysis
| Microorganisms | Parameters | Cytochrome P450 | eEF-3 |
|---|---|---|---|
| Mucorales | Score | 56.2 | 23.9 |
| E-value | 8 × 10−11 | 2.2 | |
| Identity | 25% | 26% | |
| Protein homology (organism) | Cytochrome P450 51 (Cunninghamella elegans) | Glyceraldehyde-3-phosphate dehydrogenase (Rhizomucor miehei) | |
| Penicillium | Score | 45.1 | 27.3 |
| E-value | 6 × 10−7 | 0.57 | |
| Identity | 23% | 56% | |
| Protein homology (organism) | Eburicol 14-α-demethylase (Penicillium chrysogenum) | Peroxisomal biogenesis factor 6 (Penicillium chrysogenum) | |
| P. brasiliensis | Score | 48.1 | 26.6 |
| E-value | 0.64 | 0.51 | |
| Identity | 56% | 33% | |
| Protein homology (organism) | Translation factor GUF1 | Probable Xaa-Pro aminopeptidase PADG | |
| Rhizopus | Score | 19.6 | 43.5 |
| E-value | 8.8 | 2.7 | |
| Identity | 67% | 26% | |
| Protein homology (organism) | Rhizopuspepsin-2 (Rhizopus niveus) | Peptidyl-prolyl cis–trans isomerase cyp11 (Rhizopus delemar) | |
| Fusarium | Score | 211 | 112 |
| E-value | 5 × 10−63 | 6 × 10−6 | |
| Identity | 32% | 35% | |
| Protein homology (organism) | Cytochrome P450 503A1 (Fusarium proliferatum) | Iron-sulfur clusters transporter ATM1 (Fusarium graminearum) | |
| Paecilomyces | Score | - | - |
| E-value | - | - | |
| Identity | - | - | |
| Protein homology (organism) | No match | No match |
3. Experimental Section
3.1. Fungal Strain and Culture Conditions
3.2. Preparation of Germiling Conidia Protein Extract (GTM6 h)
3.3. Patients and Control Subjects
| Patient Hospital 1 | Gender | Age | Underlying Disease | Histopathology | EORTC/MSG Classification |
|---|---|---|---|---|---|
| 1 | M | 10 | ALL/HSCT | - | Probable |
| 2 | F | 5 | MDS/HSCT | - | Probable |
| 3 | F | 39 | MDS/HSCT | - | Probable |
| 4 | F | 22 | HL/HSCT | - | Probable |
| 5 | M | 16 | ALL/HSCT | - | Probable |
| 6 | M | 34 | HL/HSCT | - | Probable |
| 7 | M | 15 | ALL/HSCT | - | Probable |
| 8 | F | 53 | CML/HSCT | - | Probable |
| 9 | M | 20 | ALL/HSCT | - | Probable |
| 10 | M | 53 | AA/HSCT | - | Probable |
| 11 | M | 50 | AML/HSCT | - | Probable |
| 12 | M | 9 | ALL/HSCT | - | Probable |
| 13 | F | 7 | ALL/HSCT | - | Probable |
| 14 | F | 29 | NHL/HSCT | A, fumigates (lung biopsy) | Proven |
| 15 | F | 11 | AML/HSCT | A, fumigates (lung biopsy) | Proven |
| 16 | F | 28 | AML/HSCT | A, flavus (lung biopsy) | Proven |
| Patient Hospital 2 | Gender | Age | Underlying Disease | Histopathology | EORTC/MSG Classification |
|---|---|---|---|---|---|
| 1 | F | 19 | AML | Aspergillus sp. (necropsy) | Proven |
| 2 | F | 28 | AML/HSCT | Aspergillus sp. (necropsy) | Proven |
| 3 | F | 50 | NHL/HSCT | Aspergillus sp. (lung biopsy) | Proven |
| 4 | F | 58 | ALL | Aspergillus sp. (necropsy) | Proven |
| 5 | M | 26 | ALL | Aspergillus sp. (laryngeal biopsy) | Proven |
| 6 | M | 58 | Lymphoma/HSCT | Aspergillus sp. (sinus biopsy) | Proven |
| 7 | M | 39 | AML | Aspergillus sp. (sinus biopsy) | Proven |
| 8 | M | 59 | NHL | Aspergillus sp. (lung biopsy) | Proven |
| 9 | F | 9 | Fulminant hepatitis/SOT | Aspergillus sp. (lung biopsy and necropsy) | Proven |
| 10 | M | 35 | AA/HSCT | Fusarium sp. (blood culture and skin biopsy) | Proven |
| 11 | M | 17 | AA/HSCT | Fusarium sp. (blood culture and skin biopsy) | Proven |
| 12 | M | 51 | NHL/HSCT | Fusarium sp. (blood culture) | Proven |
| 13 | F | 49 | No | Histoplasma sp. (lymph node biopsy and immuno-histochemistry) | Proven |
| 14 | M | 41 | No | Paracoccidioides sp. (tracheal secretion culture and direct mycroscopy of palatum) | Proven |
| 15 | F | 18 | SEL | Cryptococcus neoformans var. gattii (bronchoalveolar lavage culture) | Proven |
3.4. 2-D SDS PAGE
3.5. Western Immunoblot
3.6. Protein Identification
3.7. Homology Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Patterson, T.F. Risk stratification for invasive aspergillosis: Early assessment of host susceptibility. Med. Mycol. 2009, 47, S255–S260. [Google Scholar] [CrossRef]
- Steinbach, W.J.; Stevens, D.A.; Denning, D.W.; Moss, R.B. Advances against aspergillosis. Clin. Infect. Dis. 2003, 37, S155–S156. [Google Scholar] [CrossRef]
- Denning, D.W. Invasive aspergillosis. Clin. Infect Dis. 1998, 26, 781–805. [Google Scholar]
- Duthie, R.; Denning, D.W. Aspergillus fungemia: Report of two cases and review. Clin. Infect. Dis. 1995, 20, 598–605. [Google Scholar] [CrossRef]
- Ascioglu, S.; Rex, J.H.; de Pauw, B.; Bennett, J.E.; Bille, J.; Crokaert, F., Denning; Donnelly, J.P.; Edwards, J.E.; Erjavec, Z.; et al. Invasive fungal infections cooperative group of the European Organization for research and treatment of cancer; mycoses study group of the National Institute of allergy and infectious diseases. Defining opportunistic invasive fungal infections in immunocompromised patients with cancer and hematopoietic stem cell transplants: An international consensus. Clin. Infect. Dis. 2002, 34, 7–14. [Google Scholar] [CrossRef]
- Soubani, A.O.; Khanchandani, G.; Ahmed, H.P. Clinical significance of lower respiratory tract Aspergillus culture in elderly hospitalized patients. Eur. J. Clin. Microbiol. Infect. Dis. 2004, 23, 491–494. [Google Scholar] [CrossRef]
- Latgé, J.P. Aspergillus fumigatus and Aspergillosis. Clin. Microbiol. Rev. 1999, 12, 310–350. [Google Scholar]
- Thornton, C.R. Detection of invasive Aspergillosis. Adv. Appl. Microbiol. 2010, 70, 187–216. [Google Scholar] [CrossRef]
- Mennink-Kersten, M.A.; Donnelly, J.P.; Verweij, P.E. Detection of circulating galactomannan for the diagnosis and management of invasive aspergillosis. Lancet Infect. Dis. 2004, 4, 349–357. [Google Scholar] [CrossRef]
- Chamilos, G.; Kontoyiannis, D.P. Defining the diagnosis of invasive aspergillosis. Med. Mycol. 2006, 44, 163–172. [Google Scholar] [CrossRef]
- Beauvais, A.; Schmidt, C.; Guadagnini, S.; Roux, P.; Perret, E.; Henry, C.; Paris, S.; Mallet, A.; Prévost, M.C.; Latgé, J.P. An extracellular matrix glues together the aerial-grown hyphae of Aspergillus fumigatus. Cell Microbiol. 2007, 9, 1588–1600. [Google Scholar] [CrossRef]
- Fontaine, T.; Simenel, C.; Dubreucq, G.; Adam, O.; Delepierre, M. Molecular organization of the alkali-insoluble fraction of Aspergillus fumigatus cell wall. J. Biol. Chem. 2000, 275, 27594–27607. [Google Scholar]
- Xavier, M.O.; Pasqualotto, A.C.; Cardoso, I.C.; Severo, L.C. Cross-reactivity of Paracoccidioides brasiliensis, Histoplasma capsulatum, and Cryptococcus species in the commercial Platelia Aspergillus enzyme immunoassay. Clin. Vaccine Immunol. 2009, 16, 132–133. [Google Scholar] [CrossRef]
- Adam, O.; Aupérin, A.; Wilquin, F.; Bourhis, J.H.; Gachot, B.; Chachaty, E. Treatment with piperacillin-tazobactam and false-positive Aspergillus galactomannan antigen test results for patients with hematological malignancies. Clin. Infect. Dis. 2004, 38, 917–920. [Google Scholar] [CrossRef]
- Mikulska, M.; Furfaro, E.; del Bono, V.; Raiola, A.M.; Ratto, S.; Bacigalupo, A.; Viscoli, C. Piperacillin/tazobactam (Tazocin™) seems to be no longer responsible for false-positive results of the galactomannan assay. J. Antimicrob. Chemother. 2012, 67, 1746–1748. [Google Scholar] [CrossRef]
- Tortorano, A.M.; Esposto, M.C.; Prigitano, A.; Grancini, A.; Ossi, C.; Cavanna, C.; Cascio, G.L. Cross-reactivity of Fusarium spp. in the Aspergillus Galactomannan enzyme-linked immunosorbent assay. J. Clin. Microbiol. 2012, 50, 1051–1053. [Google Scholar] [CrossRef]
- Wheat, L.J.; Hackett, E.; Durkin, M.; Connolly, P.; Petraitiene, R.; Walsh, T.J.; Knox, K.; Hage, C. Histoplasmosis-associated cross-reactivity in the BioRad Platelia Aspergillus enzyme immunoassay. Clin. Vaccine Immunol. 2007, 14, 638–640. [Google Scholar] [CrossRef]
- De Pauw, B.; Walsh, T.J.; Donnelly, J.P.; Stevens, D.A.; Edwards, J.E.; Calandra, T.; Pappas, P.G.; Maertens, J.; Lortholary, O.; Kauffman, C.A.; et al. Revised definitions of invasive fungal disease from the European Organization for research and treatment of cancer/invasive fungal infections cooperative group and the National Institute of allergy and infectious diseases mycoses study group (EORTC/MSG) consensus group. Clin. Infect. Dis. 2008, 46, 1813–1821. [Google Scholar] [CrossRef]
- Kniemeyer, O.; Lessing, F.; Brakhage, A.A. Proteome analysis for pathogenicity and new diagnostic markers for Aspergillus fumigatus. Med. Mycol. 2009, 47, S248–S254. [Google Scholar] [CrossRef]
- Singh, B.; Oellerich, M.; Kumar, R.; Kumar, M.; Bhadoria, D.P.; Reichard, U.; Gupta, V.K.; Sharma, G.L.; Asif, A.R. Immuno-reactive molecules identified from the secreted proteome of Aspergillus fumigatus. J. Proteome Res. 2010, 9, 5517–5529. [Google Scholar] [CrossRef]
- Singh, B.; Sharma, G.L.; Oellerich, M.; Kumar, R.; Singh, S.; Bhadoria, D.P.; Katyal, A.; Reichard, U.; Asif, A.R. Novel cytosolic allergens of Aspergillus fumigatus identified from germinating conidia. J. Proteome Res. 2010, 9, 5530–5541. [Google Scholar] [CrossRef]
- Kumar, A.; Ahmed, R.; Singh, P.K.; Shukla, P.K. Identification of virulence factors and diagnostic markers using immunosecretome of Aspergillus fumigatus. J. Proteomics 2011, 74, 1104–1112. [Google Scholar] [CrossRef]
- Asif, A.R.; Oellerich, M.; Amstrong, V.W.; Gross, U.; Reichard, U. Analysis of the cellular Aspergillus fumigatus proteome that reacts with sera from rabbits developing an acquired immunity after experimental aspergillosis. Electrophoresis 2010, 31, 1947–1958. [Google Scholar] [CrossRef]
- Shi, L.N.; Li, F.Q.; Huang, M.; Lu, J.F.; Kong, X.X.; Wang, S.Q.; Shao, H.F. Immunoproteomics based identification of thioredoxin reductase GliT and novel Aspergillus fumigatus antigens for serologic diagnosis of invasive aspergillosis. BMC Microbiol. 2012, 12, 11. [Google Scholar] [CrossRef]
- Sipsas, N.V.; Kontoyiannis, D.P. Invasive fungal infections in patients with cancer in the Intensive Care Unit. Int. J. Antimicrob. Agents 2012, 39, 464–471. [Google Scholar] [CrossRef]
- Denning, D.W. Aspergillus species. In Principles and Practice of Infectious Diseases, 5th ed.; Mandell, G.L., Bennett, J.E., Dolin, R., Eds.; Churchill Livingstone: Philadelphia, PA, USA, 2000; Volume 2, pp. 2674–2685. [Google Scholar]
- Pitarch, A.; Jiménez, A.; Nombela, C.; Gil, C. Decoding serological response to Candida cell wall immunome into novel diagnostic, prognostic, and therapeutic candidates for systemic candidiasis by proteomic and bioinformatic analyses. Mol. Cell. Proteomics 2006, 5, 79–96. [Google Scholar]
- Lai, H.Y.; Tam, M.F.; Tang, R.B.; Chou, H.; Chang, C.Y.; Tsai, J.J.; Shen, H.D. cDNA cloning and immunological characterization of a newly identified enolase allergen from Penicillium citrinum and Aspergillus fumigatus. Int. Arch. Allergy Immunol. 2002, 127, 181–190. [Google Scholar] [CrossRef]
- Rohde, M.; Schwienbacher, M.; Nikolaus, T.; Heesemann, J.; Ebel, F. Detection of early phase specific surface appendages during germination of Aspergillus fumigatus conidia. FEMS Microbiol. Lett. 2002, 206, 99–105. [Google Scholar] [CrossRef]
- Rhodes, C.J. Aspergillus fumigatus: Growth and virulence. Med. Mycol. 2006, 44, 77–81. [Google Scholar] [CrossRef]
- Kubitschek-Barreira, P.H.; Curty, N.; Neves, G.W.; Gil, C.; Lopes-Bezerra, L.M. Differential proteomic analysis of Aspergillus fumigatus morphotypes reveals putative drug targets. J. Proteomics 2013, 78, 522–534. [Google Scholar] [CrossRef]
- Denikus, N.; Orfaniotou, F.; Wulf, G.; Lehmann, P.F.; Monod, M.; Reichard, U. Fungal antigens expressed during invasive aspergillosis. Infect. Immun. 2005, 73, 4704–4713. [Google Scholar] [CrossRef]
- Lamaris, G.A.; Chamilos, G.; Lewis, R.E.; Safdar, A.; Raad, II.; Kontoyiannis, D.P. Scedosporium infection in a tertiary care cancer center: A review of 25 cases from 1989–2006. Clin. Infect. Dis. 2006, 43, 1580–1584. [Google Scholar] [CrossRef]
- Cortez, K.J.; Roilides, E.; Quiroz-Telles, F.; Meletiadis, J.; Antachopoulos, C.; Knudsen, T.; Buchanan, W.; Milanovich, J.; Sutton, D.A.; Fothergill, A.; et al. Infections caused by Scedosporium spp. Clin. Microbiol. Rev. 2008, 21, 157–197. [Google Scholar] [CrossRef]
- Mori, Y.; Nagasaki, Y.; Kamezaki, K.; Takenaka, K.; Iwasaki, H.; Harada, N.; Miyamoto, T.; Abe, Y.; Shimono, N.; Akashi, K.; et al. High incidence of false-positive Aspergillus galactomannan test in multiple myeloma. Am. J. Hematol. 2010, 5, 449–451. [Google Scholar]
- Morrissey, C.O. Advancing the field: Evidence for new management strategies in invasive fungal infections. Curr. Fungal. Infect. Rep. 2013, 7, 51–58. [Google Scholar] [CrossRef]
- Vidovic, A.; Arsic-Arsenijevic, V.; Tomin, D.; Djunic, I.; Jakovic, R.; Loncar, Z.; Barac, A. Proven invasive pulmonary mucormycosis successfully treated with amphotericin B and surgery in patient with acute myeloblastic leukemia: A case report. J. Med. Case Rep. 2013, 7, 263. [Google Scholar] [CrossRef]
- Crešnar, B.; Petrič, S. Cytochrome P450 enzymes in the fungal kingdom. Biochim. Biophys. Acta 2010, 1814, 29–35. [Google Scholar]
- Ehrlich, K.C.; Chang, P.K.; Yu, J.; Cotty, P.J. Aflatoxin biosynthesis cluster gene cypA is required for G aflatoxin formation. Appl. Environ. Microbiol. 2004, 70, 6518–6524. [Google Scholar] [CrossRef]
- Wen, Y.; Hatabayashi, H.; Arai, H.; Kitamoto, H.K.; Yabe, K. Function of the cypX and moxY genes in aflatoxin biosynthesis in Aspergillus parasiticus. Appl. Environ. Microbiol. 2005, 71, 3192–3198. [Google Scholar] [CrossRef]
- Lepesheva, G.I.; Waterman, M.R. Sterol 14-α-demethylase cytochrome P450 (CYP51), a P450 in all biological kingdoms. Biochim. Biophys. Acta 2007, 1770, 467–477. [Google Scholar]
- Verweij, P.E.; Mellado, E.; Melchers, W. Multiple-triazole-resistant aspergillosis. N. Engl. J. Med. 2007, 356, 1481–1483. [Google Scholar] [CrossRef]
- Mavridou, E.; Bruggemann, R.J.; Melchers, W.J.; Verweij, P.E.; Mouton, J.W. Impact of cyp51A mutations on the pharmacokinetic and pharmacodynamics properties of voriconazole in a murine model of disseminated aspergillosis. Antimicrob. Agents Chemother. 2010, 54, 4758–4764. [Google Scholar] [CrossRef]
- Kuipers, S.; Brüggemann, R.J.; de Sévaux, R.G.; Heesakkers, J.P.; Melchers, W.J.; Mouton, J.W.; Verweij, P.E. Failure of posaconazole therapy in a renal transplant patient with invasive aspergillosis due to Aspergillus fumigatus with attenuated susceptibility to posaconazole. Antimicrob. Agents Chemother. 2011, 55, 3564–3566. [Google Scholar] [CrossRef]
- Van den Brink, H.M.; van Gorcom, R.F.; van den Hondel, C.A.; Punt, P.J. Cytochrome P450 enzyme systems in fungi. Fungal Genet. Biol. 1998, 23, 1–17. [Google Scholar] [CrossRef]
- Nelson, D.R. Cytochrome P450 and the individuality of species. Arch. Biochem. Biophys. 1999, 369, 1–10. [Google Scholar] [CrossRef]
- Nelson, D.R. Cytochrome P450 nomenclature. Methods Mol. Cell. Biol. 1998, 107, 15–24. [Google Scholar]
- Nelson, D.R. Metazoan cytochrome P450 evolution. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1998, 121, 15–22. [Google Scholar]
- Hinnebusch, A.G.; Dever, T.E.; Asano, K. Translational Control in Biology and Medicine; Sonenberg, N., Hershey, J.W.B., Mathews, M.B., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2007; pp. 225–268. [Google Scholar]
- Carvalho, M.D.; Carvalho, J.F.; Merrick, W.C. Biological characterization of various forms of elongation factor 1 from rabbit reticulocytes. Arch. Biochem. Biophys. 1984, 234, 603–611. [Google Scholar] [CrossRef]
- Skogerson, L.; Moldave, K. Characterization of the interaction of aminoacyltransferase II with ribosomes. Binding of transferase II and translocation of peptidyl transfer ribonucleic acid. J. Biol. Chem. 1968, 243, 5354–5360. [Google Scholar]
- Liu, M.; Gelli, A. Elongation factor 3, EF3, associates with the calcium channel Cch1 and targets Cch1 to the plasma membrane in Cryptococcus neoformans. Eukaryot. Cell 2008, 7, 1118–1126. [Google Scholar] [CrossRef]
- Hollenstein, K.; Dawson, R.J.; Locher, K.P. Structure and mechanism of ABC transporter proteins. Curr. Opin. Struct. Biol. 2007, 17, 412–418. [Google Scholar] [CrossRef]
- Uritani, M.; Miyazaki, M. Characterization of the ATPase and GTPase activities of the elongation factor 3 (EF-3) from yeasts. J. Biochem. 1988, 103, 522–530. [Google Scholar]
- Sasikumar, A.N.; Kinzy, T.G. Mutations in the chromodomain-like insertion of translation elongation factor 3 compromise protein synthesis through reduced ATPase activity. J. Biol. Chem. 2014, 289, 4853–4860. [Google Scholar] [CrossRef]
- López-Ribot, J.L.; Alloush, H.M.; Masten, B.J.; Chaffin, W.L. Evidence for presence in the cell wall of Candida albicans of a protein related to the hsp70 family. Infect. Immun. 1996, 64, 3333–3340. [Google Scholar]
- Yuan, S.Y. Protein kinase signalling in the modulation of microvascular permeability. Vascul. Pharmacol. 2002, 39, 213–223. [Google Scholar] [CrossRef]
- Pardo, M.; Ward, M.; Bains, S.; Molina, M.; Blackstock, W.; Gil, C.; Nombela, C. A proteomic approach for the study of Saccharomyces cerevisiae cell wall biogenesis. Electrophoresis 2000, 21, 3396–3410. [Google Scholar] [CrossRef]
- Pitarch, A.; Sanchez, M.; Nombela, C.; Gil, C. Sequential fractionation and two-dimensional gel analysis unravels the complexity of the dimorphic fungus Candida albicans cell wall proteome. Mol. Cell. Proteomics 2002, 1, 967–982. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Virginio, E.D.; Kubitschek-Barreira, P.H.; Batista, M.V.; Schirmer, M.R.; Abdelhay, E.; Shikanai-Yasuda, M.A.; Lopes-Bezerra, L.M. Immunoproteome of Aspergillus fumigatus Using Sera of Patients with Invasive Aspergillosis. Int. J. Mol. Sci. 2014, 15, 14505-14530. https://doi.org/10.3390/ijms150814505
Virginio ED, Kubitschek-Barreira PH, Batista MV, Schirmer MR, Abdelhay E, Shikanai-Yasuda MA, Lopes-Bezerra LM. Immunoproteome of Aspergillus fumigatus Using Sera of Patients with Invasive Aspergillosis. International Journal of Molecular Sciences. 2014; 15(8):14505-14530. https://doi.org/10.3390/ijms150814505
Chicago/Turabian StyleVirginio, Emylli D., Paula H. Kubitschek-Barreira, Marjorie Vieira Batista, Marcelo R. Schirmer, Eliana Abdelhay, Maria A. Shikanai-Yasuda, and Leila M. Lopes-Bezerra. 2014. "Immunoproteome of Aspergillus fumigatus Using Sera of Patients with Invasive Aspergillosis" International Journal of Molecular Sciences 15, no. 8: 14505-14530. https://doi.org/10.3390/ijms150814505