Multi-Step Pathogenesis and Induction of Local Immune Response by Systemic Candida Albicans Infection in an Intravenous Challenge Mouse Model

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
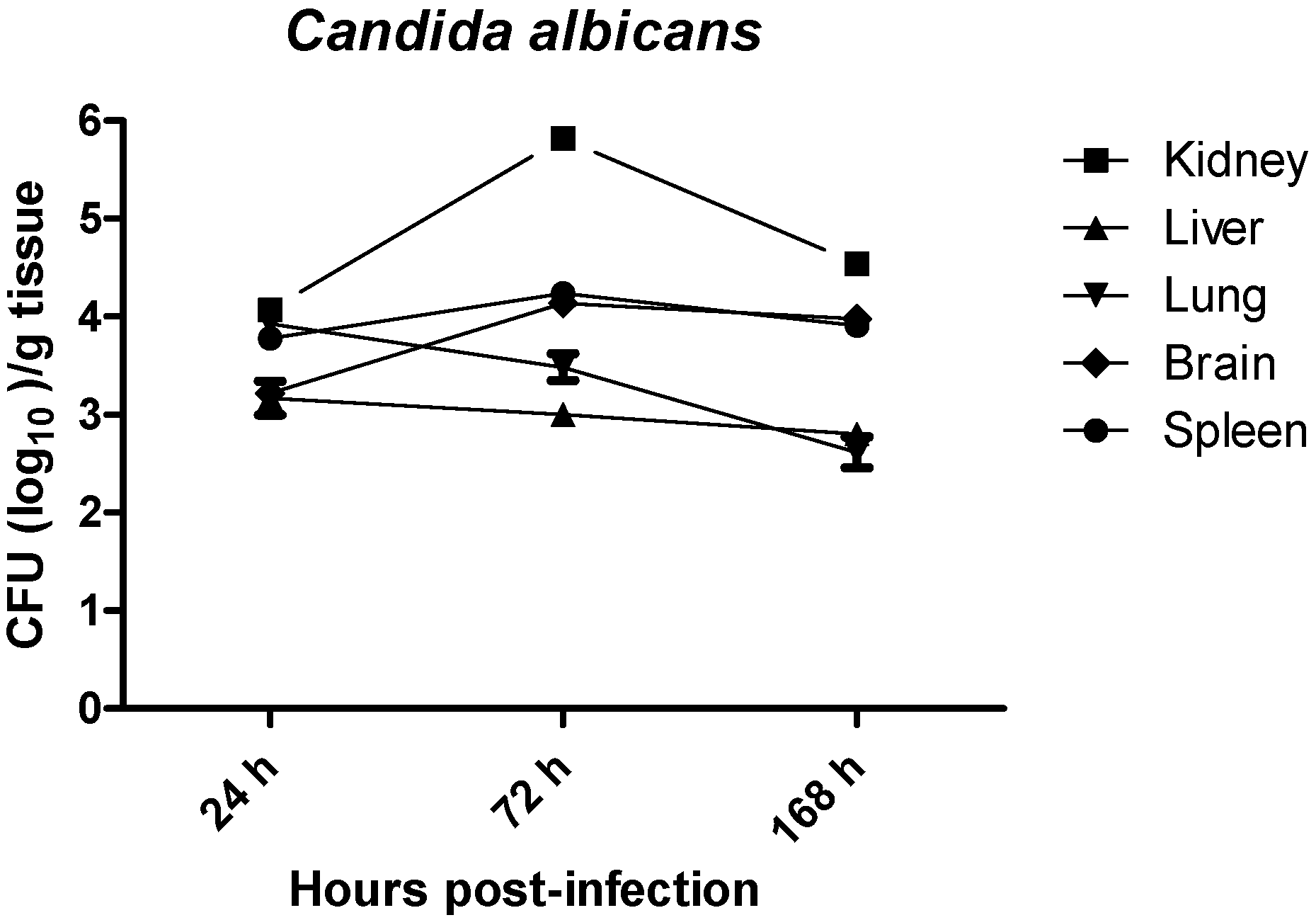
2.1. Quantitative Yeast Count
2.2. In Vitro Haemolysin Production by C. albicans
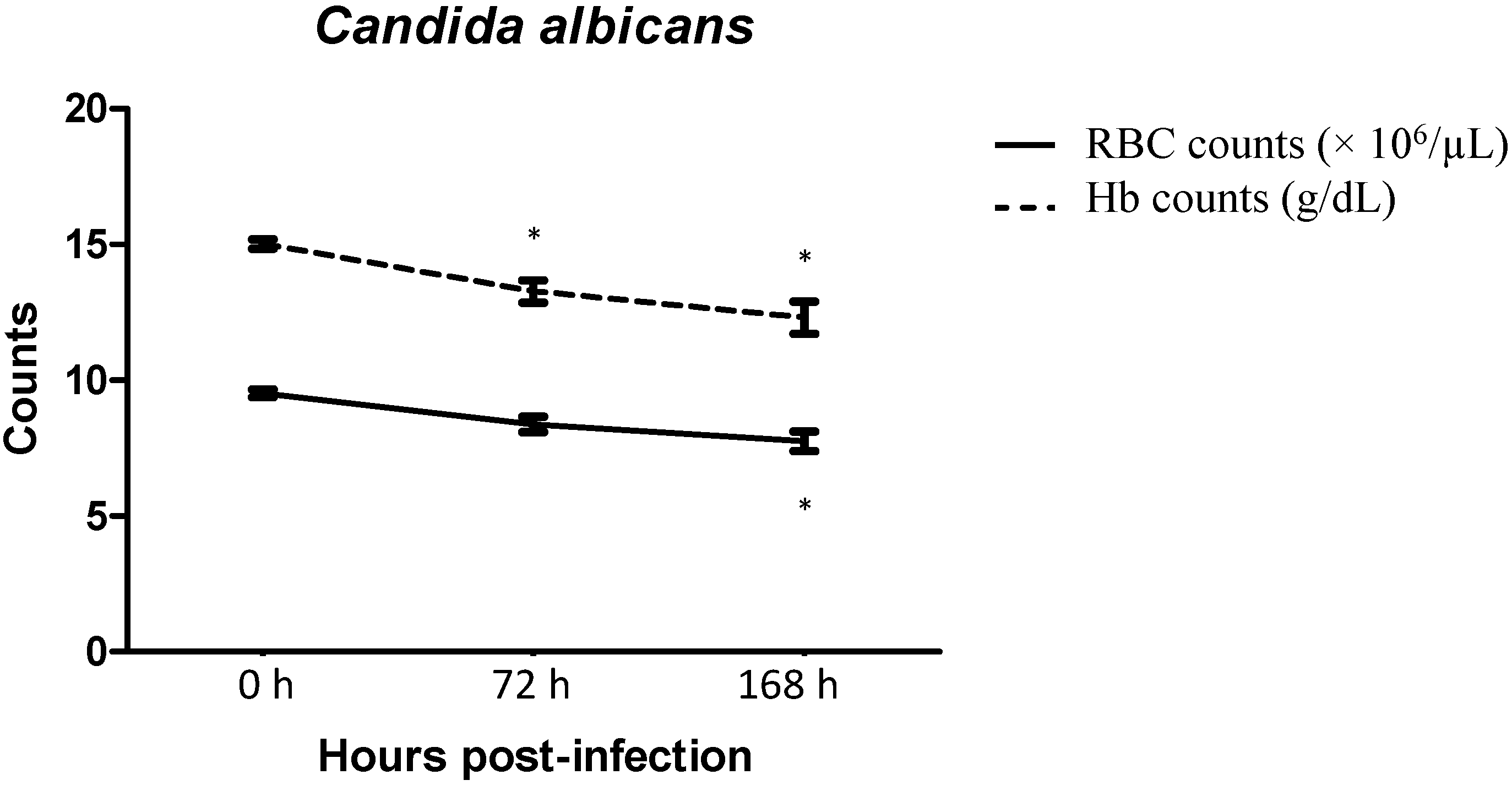
2.3. Red Blood Cells and Haemoglobin Counts


2.4. Murine Erythropoietin Expression

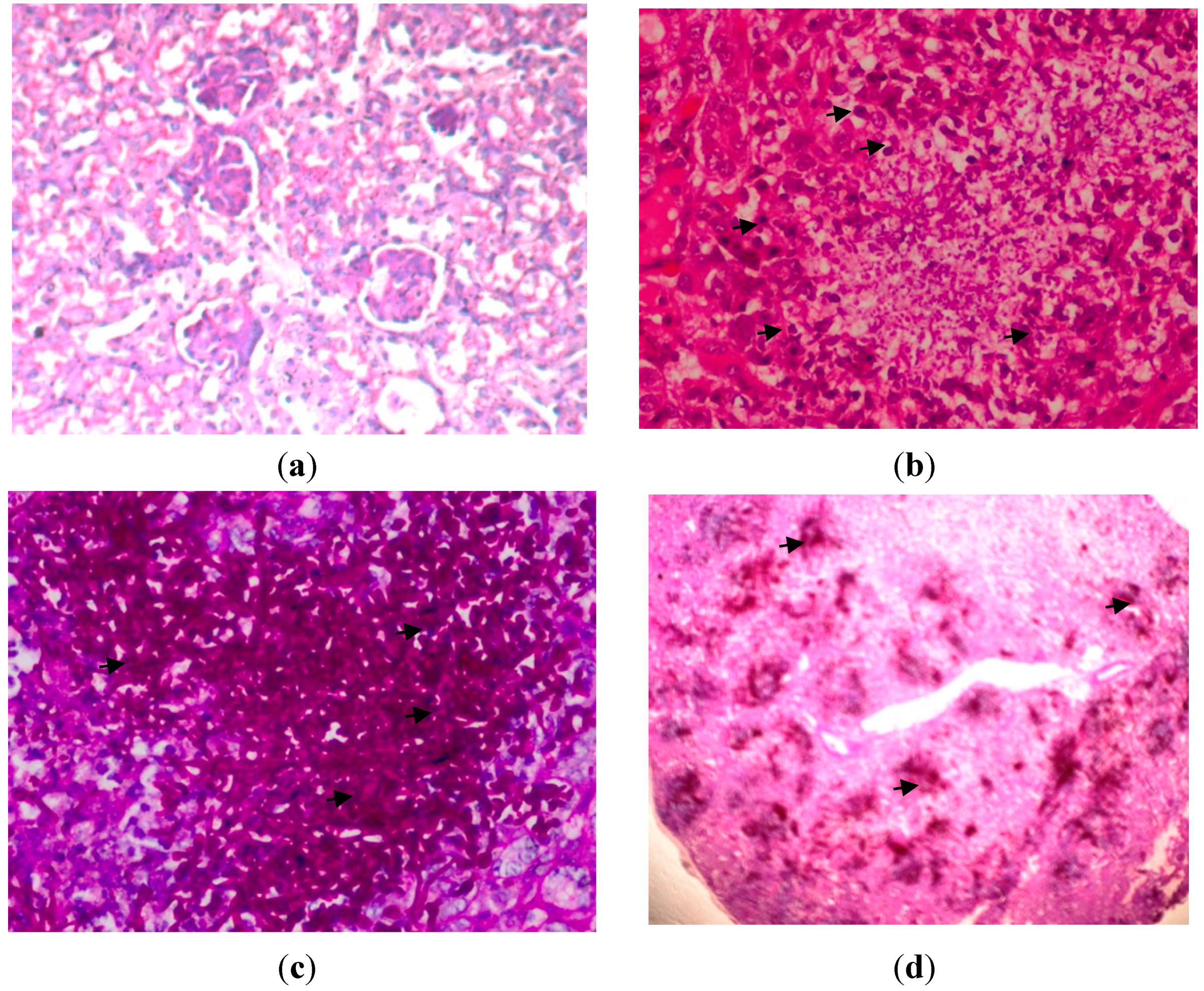
2.5. Histopathology



2.6. Pattern Recognition Receptor (PRR) Gene Expression during Infection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Kidney | Brain | Spleen | |||
|---|---|---|---|---|---|---|
| Gene | Time | |||||
| 24 h Mean Fold Changes/p Values | 72 h Mean Fold Changes/p Values | 24 h Mean Fold Changes/p Values | 72 h Mean Fold Changes/p Values | 24 h Mean Fold Changes/p Values | 72 h Mean Fold Changes/p Values | |
| Tlr2 | 13.03/0.0022 | 10.58/0.0002 | 3.71/0.0092 | 7.18/0.00007 | 1.96/0.023 | 1.19/0.194 |
| Tlr4 | 1.51/0.108 | 3.94/0.006 | 1.27/0.087 | 3.11/0.0012 | −1.01/0.88 | 1.09/0.589 |
| Tlr9 | 2.11/0.145 | 1.01/0.807 | −1.61/0.123 | −4.78/0.012 | −1.59/0.002 | −3.77/0.008 |
| Mrc1 | −2.39/0.208 | 2.00/0.235 | −1.50/0.097 | −1.92/0.04 | −1.22/0.146 | 3.15/0.00002 |
| Mrc2 | −2.53/0.0073 | 1.62/0.053 | 2.09/0.264 | −1.33/0.291 | −1.44/0.188 | 1.03/0.924 |
| Clec7a | 6.48/0.0061 | 4.18/0.04 | 1.66/0.410 | 1.95/0.148 | −1.17/0.251 | −2.45/0.035 |
| Clec4n | 18.89/0.0018 | 40.63/0.036 | 4.48/0.0004 | 17.12/0.00009 | 2.40/0.0004 | −1.04/0.933 |
| Clec4e | 24.41/0.00181 | 75.51/0.0017 | 8.02/0.08 | 20.93/0.00011 | 6.75/0.003 | 6.63/0.00074 |
2.7. Cytokine Gene Expression during Infection
| Organ | Kidney | Brain | Spleen | |||
|---|---|---|---|---|---|---|
| Gene | Time | |||||
| 24 h Mean Fold Changes/p Values | 72 h Mean Fold Changes/p Values | 24 h Mean Fold Changes/p Values | 72 h Mean Fold Changes/p Values | 24 h Mean Fold Changes/p Values | 72 h Mean Fold Changes/p Values | |
| Tgfbi | 3.62/0.0012 | 21.71/0.001 | 2.21/0.016 | 7.90/0.00011 | 1.81/0.0006 | 2.08/0.051 |
| Il10 | 6.23/0.0019 | 121.66/0.048 | 6.82/0.18 | 46.35/0.00002 | 6.44/0.0018 | 21.16/0.031 |
| Tnf | 65.04/0.0006 | 36.93/0.002 | 12.61/0.055 | 58.13/0.0005 | −1.57/0.173 | −3.02/0.012 |
| Il13 | 1.35/0.8851 | 1.49/0.734 | 3.90/0.227 | 1.12/0.863 | 2.22/0.355 | 1.24/0.853 |
| Ifng | 12.32/0.1147 | −2.14/0.059 | 6.71/0.202 | 1.99/0.447 | −1.02/0.829 | −8.40/0.00002 |
| Il23a | 8.07/0.0013 | 21.51/0.0006 | 3.32/0.245 | 3.19/0.008 | 4.26/0.021 | 1.21/0.818 |
| Il6 | 35.17/0.016 | 49.15/0.056 | 15.78/0.005 | 83.93/0.0001 | 1.36/0.466 | −1.65/0.918 |
| Il4 | −1.54/0.545 | −1.00/0.923 | 5.66/0.121 | −1.04/0.776 | −1.15/0.689 | −2.54/0.0011 |
| Il17a | 1.08/0.980 | 1.43/0.446 | 10.80/0.168 | 3.10/0.029 | 7.10/0.008 | −1.19/0.527 |
| Il12a | 20.87/0.0037 | 80.26/0.017 | 1.77/0.174 | 5.06/0.0008 | −1.35/0.120 | 1.08/0.671 |
2.8. Chemokine Gene Expression during Infection
| Organ | Kidney | Brain | Spleen | |||
|---|---|---|---|---|---|---|
| Gene | Time | |||||
| 24 h Mean Fold Changes/p Values | 72 h Mean Fold Changes/p Values | 24 h Mean Fold Changes/p Values | 72 h Mean Fold Changes/p Values | 24 h Mean Fold Changes/p Values | 72 h Mean Fold Changes/p Values | |
| Cxcl1 | 104.21/0.0235 | 126.02/0.0007 | 64.59/0.077 | 27.81/0.0002 | 6.89/0.001 | −1.43/0.950 |
| Cxcl10 | 76.82/0.024 | −1.03/0.951 | 6.63/0.015 | 1.28/0.361 | 2.57/0.00006 | −17.84/0.003 |
| Ccl5 | 17.43/0.016 | 1.30/0.651 | −13.36/0.047 | −8.22/0.049 | −1.77/0.00006 | −5.45/0.0005 |
| Cxcl9 | 29.72/0.140 | −2.14/0.322 | 1.49/0.442 | −3.69/0.051 | 3.08/0.0005 | −13.24/0.004 |
| Csf2 | 1.21/0.583 | 3.53/0.356 | −1.02/0.473 | −1.88/0.338 | −2.04/0.730 | 1.18/0.487 |
| Cxcl11 | 56.75/0.019 | 1.95/0.100 | 6.25/0.036 | 2.44/0.10 | 1.50/0.304 | −2.93/0.123 |
| Ccl12 | 11.71/0.074 | −1.20/0.644 | 32.37/0.022 | −13.63/0.04 | 23.22/0.006 | 2.02/0.235 |
2.9. Discussion
3. Experimental Section
3.1. Ethical Statement
3.2. Mice
3.3. Fungal Inoculum and Animal Inoculation
3.4. Quantitative Yeast Count
3.5. Determination of Haemolysin Activity
3.6. Red Blood Cells and Haemoglobin Counts
3.7. Erythropoietin Expression
3.8. Pathology
3.9. Local Host Immune Response
3.10. Data and Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ozcan, S.K.; Budak, F.; Willke, A.; Filiz, S.; Costur, P.; Dalcik, H. Efficacies of caspofungin and a combination of caspofungin and merepenem in the treatment of murine disseminated candidiasis. APMIS 2006, 114, 829–836. [Google Scholar]
- Pedraz, J.; Delgado-Jiménez, Y.; Pérez-Gala, S.; Nam-Cha, S.; Fernández-Herrera, J.; García-Diez, A. Cutaneous expression of systemic candidiasis. Clin. Exp. Dermatol. 2009, 34, 106–110. [Google Scholar]
- Perlroth, J.; Choi, B.; Spellberg, B. Nosocomial fungal infections: Epidemiology, diagnosis, and treatment. Med. Mycol. 2007, 45, 321–346. [Google Scholar] [CrossRef]
- Spellberg, B.; Ibrahim, A.S.; Edwards, J.E., Jr.; Filler, S.G. Mice with disseminated candidiasis die of progressive sepsis. J. Infect. Dis. 2005, 192, 336–343. [Google Scholar]
- Tuite, A.; Elias, M.; Picard, S.; Mullick, A.; Gros, P. Genetic control of susceptibility to Candida albicans in susceptible A/J and resistant C57BL/6J mice. Genes Immun. 2005, 6, 672–682. [Google Scholar]
- Pfaller, M.A.; Pappas, P.G.; Wingard, J.R. Invasive fungal pathogens: Current epidemiological trends. Clin. Infect. Dis. 2006, 43, S3–S14. [Google Scholar]
- Arendup, M.; Horn, T.; Frimodt-Moller, N. In vivo pathogenicity of eight medically relevant Candida species in an animal model. Infection 2002, 30, 286–291. [Google Scholar] [CrossRef]
- MacCallum, D.M. Massive induction of innate immune response to Candida albicans in the kidney in a murine intravenous challenge model. FEMS Yeast 2009, 9, 1111–1122. [Google Scholar] [CrossRef]
- Ashman, R.B.; Fulurija, A.; Papadimitriou, J.M. Strain-dependent differences in host response to Candida albicans infection in mice are related to organ susceptibility and infectious load. Infect. Immun. 1996, 64, 1866–1869. [Google Scholar]
- Brieland, J.; Essig, D.; Jackson, C.; Frank, D.; Loebenberg, D.; Menzel, F.; Arnold, B.; DiDomenico, B.; Hare, R. Comparison of pathogenesis and host immune responses to Candida glabrata and Candida albicans in systemically infected immunocompetent mice. Infect. Immun. 2001, 69, 5046–5055. [Google Scholar] [CrossRef]
- Westenfelder, C.; Biddle, D.L.; Baranowski, R.L. Human, rat, and mouse kidney cells express functional erythropoietin receptors. Kidney Int. 1999, 55, 808–820. [Google Scholar] [CrossRef]
- Maxwell, P.H.; Osmond, M.K.; Pugh, C.W.; Heryet, A.; Nicholls, L.G.; Tan, C.C.; Doe, B.G.; Ferguson, D.J.; Johnson, M.H.; Ratcliffe, P.J. Identification of the renal erythropoietin-producing cells using transgenic mice. Kidney Int. 1993, 44, 1149–1162. [Google Scholar]
- Pan, X.; Suzuki, N.; Hirano, I.; Yamazaki, S.; Minegishi, N.; Yamamoto, M. Isolation and characterization of renal erythropoietin producing cells from genetically produced anemia mice. PLoS One 2011. [Google Scholar] [CrossRef]
- Chasis, J.A.; Mohandas, N. Erythroblastic islands: Niches for erythropoiesis. Blood 2008, 112, 470–478. [Google Scholar]
- Fradin, C.; de Groot, P.; MacCallum, D.; Schaller, M.; Klis, F.; Odds, F.C.; Hube, B. Granulocytes govern the transcriptional response, morphology and proliferation of Candida albicans in human blood. Mol. Microbiol. 2005, 56, 397–415. [Google Scholar]
- Fradin, C.; Mavor, A.L.; Weindl, G.; Schaller, M.; Hanke, K.; Kaufmann, S.H.; Mollenkopf, H.; Hube, B. The early transcriptional response of human granulocytes to infection with Candida albicans is not essential for killing, but reflects cellular communications. Infect. Immun. 2006, 75, 1493–1501. [Google Scholar]
- Muller, V.; Viemann, D.; Schmidt, M.; Endres, N.; Ludwig, S.; Leverkus, M.; Roth, J.; Goebeler, M. Candida albicans triggers activation of distinct signaling pathways to establish a proinflammatory gene expression program in primary human endothelial cells. J. Immunol. 2007, 179, 8435–8445. [Google Scholar] [CrossRef]
- Barker, K.S.; Park, H.; Phan, Q.T.; Xu, L.; Homayouni, R.; Rogers, P.D.; Filler, S.G. Transcriptome profile of the vascular endothelial cell response to Candida albicans. J. Infect. Dis. 2008, 198, 193–202. [Google Scholar]
- Prigneau, O.; Porta, A.; Poudrier, J.A.; Colonna-Romano, S.; Noel, T.; Maresca, B. Genes involved in beta-oxidation, energy metabolism and glyoxylate cycle are induced by Candida albicans during macrophage infection. Yeast 2003, 20, 723–730. [Google Scholar]
- Lorenz, M.C.; Bender, J.A.; Fink, G.R. Transcriptional response of Candida albicans upon internalization by macrophages. Eukaryot. Cell 2004, 3, 1076–1087. [Google Scholar] [CrossRef]
- Wolfs, T.G.; Buurman, W.A.; van Schadewijk, A.; de Vries, B.; Daemen, M.A.; Hiemstra, P.S.; van ’t Veer, C. In vivo expression of Toll-like receptor 2 and 4 by renal epithelial cells: IFN-gamma and TNF-alpha mediated upregulation during inflammation. J. Immunol. 2002, 168, 1286–1293. [Google Scholar] [CrossRef]
- Shigeoka, A.A.; Holscher, T.D.; King, A.J.; Hall, F.W.; Kiosses, W.B.; Tobias, P.S.; Mackman, N.; McKay, D.B. TLR2 is constitutively expressed within the kidney and participates in ischemic renal injury through both MyD88-dependent and -independent pathways. J. Immunol. 2007, 178, 6252–6258. [Google Scholar]
- Shah, V.B.; Huang, Y.; Keshwara, R.; Ozment-Skelton, T.; Williams, D.L.; Keshvara, L. Beta-glucan activates microglia without inducing cytokine production in Dectin-1-dependent manner. J. Immunol. 2008, 180, 2777–2785. [Google Scholar]
- Shah, V.B.; Williams, D.L.; Keshvara, L. b-Glucan attenuates TLR2- and TLR4-mediated cytokine production by microglia. Neurosci. Lett. 2009, 458, 111–115. [Google Scholar]
- Ishibashi, K.; Miura, N.N.; Adachi, Y.; Ogura, N.; Tamura, H.; Tanaka, S.; Ohno, N. DNA array analysis of altered gene expression in human leukocytes stimulated with soluble and particulate forms of Candida cell wall beta-glucan. Int. Immunopharmacol. 2004, 4, 387–401. [Google Scholar]
- Sarafi, M.N.; Garcia-Zepeda, E.A.; MacLean, J.A.; Charo, I.F.; Luster, A.D. Murine monocyte chemoattractant protein (MCP)-5: A novel CC chemokine that is a structural and functional homologue of human MCP-1. J. Exp. Med. 1997, 185, 99–109. [Google Scholar]
- Strieter, R.M.; Kunkel, S.L.; Standiford, T.J. Chemokines in the Lung; Dekker: New York, NY, USA, 2003. [Google Scholar]
- Roson, M.I.; Cavallero, S.; della Penna, S.; Cao, G.; Gorzalczany, S.; Pandolfo, M.; Kuprewicz, A.; Canessa, O.; Toblli, J.E.; Fernández, B.E. Acute sodium overload produces renal tubulointerstitial inflammation in normal rats. Kidney Int. 2006, 70, 1439–1446. [Google Scholar]
- Hertting, O.; Khalil, A.; Jaremko, G.; Chromek, M.; Li, Y.H.; Bakhiet, M.; Bartfai, T.; Tullus, K.; Brauner, A. Enhanced chemokine response in experimental acute Escherichia colipyelonephritis in IL-1β-deficient mice. Clin. Exp. Immunol. 2003, 131, 225–233. [Google Scholar] [CrossRef]
- Hardison, J.L.; Wrightsman, R.A.; Carpenter, P.M.; Kuziel, W.A.; Lane, T.E.; Manning, J.E. The CC chemokine receptor 5 is important in control of parasite replication and acute cardiac inflammation following infection with trypanosoma cruzi. Infect. Immun. 2006, 74, 135–143. [Google Scholar]
- Yang, D.; Chen, Q.; Hoover, D.M.; Staley, P.; Tucker, K.D.; Lubkowski, J.; Oppenheim, J.J. Many chemokines including CCL20/MIP-3alpha display antimicrobial activity. J. Leukoc. Biol. 2003, 74, 448–455. [Google Scholar]
- Söbirk, S.K.; Mörgelin, M.; Egesten, A.; Bates, P.; Shannon, O.; Collin, M. Human chemokines as antimicrobial peptides with direct parasiticidal effect on Leishmania. mexicana in vitro. PLoS One 2013, 8, e58129. [Google Scholar] [CrossRef] [Green Version]
- Romani, L.; Howard, D.H. Mechanisms of resistance to fungal infection. Curr. Opin. Immunol. 1995, 7, 517–523. [Google Scholar]
- Romani, L.; Mencacci, A.; Tonnetti, L.; Spaccapelo, R.; Cenci, E.; Wolf, S.; Puccetti, P.; Bistoni, F. Interleukin-12 but not interferon-gamma production correlates with induction of T-helper type-1 phenotype in murine candidiasis. Eur. J. Immunol. 1994, 24, 909–915. [Google Scholar]
- Harrington, L.E.; Hatton, R.D.; Mangan, P.R.; Turner, H.; Murphy, T.L.; Murphy, K.M.; Weaver, C.T. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat. Immunol. 2005, 6, 1123–1132. [Google Scholar]
- Tan, Z.Y.; Bealgey, K.W.; Fang, Y.; Gong, Y.M.; Bao, S. Interleukin-23: Immunological roles and clinical implications. Int. J. Biochem. Cell Biol. 2009, 41, 733–735. [Google Scholar]
- Netea, M.G.; van Tits, L.J.; Curfs, J.H.; Amiot, F.; Meis, J.F.; van der Meer, J.W.; Kullberg, B.J. Increased susceptibility of TNF-alpha lymphotoxin-alpha double knockout mice to systemic candidiasis through impaired recruitment of neutrophils and phagocytosis of Candida albicans. J. Immunol. 1999, 163, 1498–1505. [Google Scholar]
- Kunze, R.; Schutz, B.; Schrodl, W.; Bernhardt, H. Candida albicans clinical isolates inactivated by formalin with different adherence to buccal epithelial cells induce proinflammatory and regulatory cytokines in human peripheral blood mononuclear cells. Mycoses 1999, 42, 529–538. [Google Scholar] [CrossRef]
- Netea, M.G.; van Der Graaf, C.A.; Vonk, A.G.; Verschueren, I.; van der Meer, J.W.; Kullberg, B.J. The role of toll-like receptor (TLR) 2 and TLR4 in the host defense against disseminated candidiasis. J. Infect. Dis. 2002, 185, 1483–1489. [Google Scholar]
- Sherry, R.M.; Cue, J.I.; Goddard, J.K.; Parramore, J.B.; DiPiro, J.T. Interleukin-10 is associated with the development of sepsis in trauma patients. J. Trauma 1996, 40, 613–617. [Google Scholar]
- Hildebrand, F.; Pape, H.C.; Hoevel, P.; Krettek, C.; van Griensven, M. The importance of systemic cytokines in the pathogenesis of polymicrobial sepsis and dehydroepiandrosterone treatment in a rodent model. Shock 2003, 20, 338–346. [Google Scholar]
- Cassatella, M.A. The neutrophil: One of the cellular targets of interleukin-10. Int. J. Clin. Lab. Res. 1998, 28, 148–161. [Google Scholar]
- Cassatella, M.A.; Meda, L.; Bonora, S.; Ceska, M.; Constantin, G. Interleukin 10 (IL-10) inhibits the release of proinflammatory cytokines from human polymorphonuclear leukocytes. Evidence for an autocrine role of tumor necrosis factor and IL-1 beta in mediating the production of IL-8 triggered by lipopolysaccharide. J. Exp. Med. 1993, 178, 2207–2211. [Google Scholar] [CrossRef]
- Cassatella, M.A.; Meda, L.; Gasperini, S.; Calzetti, F.; Bonora, S. Interleukin 10 (IL-10) upregulatesIL-1 receptor antagonist production from lipopolysaccharide-stimulated human polymorphonuclear leukocytes by delaying mRNA degradation. J. Exp. Med. 1994, 179, 1695–1699. [Google Scholar]
- Netea, M.G.; Sutmuller, R.; Hermann, C.; van der Graaf, C.A.; van der Meer, J.W.; van Krieken, J.H.; Hartung, T.; Adema, G.; Kullberg, B.J. Toll-like receptor 2 suppresses immunity against Candida albicans through induction of IL-10 and regulatory T cells. J. Immunol. 2004, 172, 3712–3718. [Google Scholar]
- De Luca, A.; Montagnoli, C.; Zelante, T.; Bonifazi, P.; Bozza, S.; Moretti, S.; D’Angelo, C.; Vacca, C.; Boon, L.; Bistoni, F.; et al. Functional yet balanced reactivity to Candida albicans requires TRIF, MyD88, and IDO-dependent inhibition of Rorc. J. Immunol. 2007, 179, 5999–6008. [Google Scholar] [CrossRef]
- Luo, G.; Samaranayake, L.P.; Yau, J.Y.Y. Candida species exhibit differential in vitro hemolytic activities. J. Clin. Microbiol. 2001, 39, 2971–2974. [Google Scholar] [CrossRef]
- RT2 Profiler PCR Array Data Analysis Version 3.5. Available online: http://pcrdataanalysis.sabiosciences.com/pcr/arrayanalysis.php (accessed on 20 March 2014).
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chin, V.-K.; Foong, K.-J.; Maha, A.; Rusliza, B.; Norhafizah, M.; Chong, P.P. Multi-Step Pathogenesis and Induction of Local Immune Response by Systemic Candida Albicans Infection in an Intravenous Challenge Mouse Model. Int. J. Mol. Sci. 2014, 15, 14848-14867. https://doi.org/10.3390/ijms150814848
Chin V-K, Foong K-J, Maha A, Rusliza B, Norhafizah M, Chong PP. Multi-Step Pathogenesis and Induction of Local Immune Response by Systemic Candida Albicans Infection in an Intravenous Challenge Mouse Model. International Journal of Molecular Sciences. 2014; 15(8):14848-14867. https://doi.org/10.3390/ijms150814848
Chicago/Turabian StyleChin, Voon-Kin, Kuan-Jeang Foong, Abdullah Maha, Basir Rusliza, Mohtarrudin Norhafizah, and Pei Pei Chong. 2014. "Multi-Step Pathogenesis and Induction of Local Immune Response by Systemic Candida Albicans Infection in an Intravenous Challenge Mouse Model" International Journal of Molecular Sciences 15, no. 8: 14848-14867. https://doi.org/10.3390/ijms150814848