Farm Animal Serum Proteomics and Impact on Human Health

Abstract
:
1. Introduction

Animal Genome Databases and Annotation of Gene Functions

{kind=link}
{kind=link}
| GO Annotation | Number of Associations | Number of Distinct Proteins |
|---|---|---|
| Human | 403,735 | 46,159 |
| Chicken | 93,784 | 14,238 |
| Cow | 128,684 | 20,032 |
| Pig | 107,097 | 19,723 |
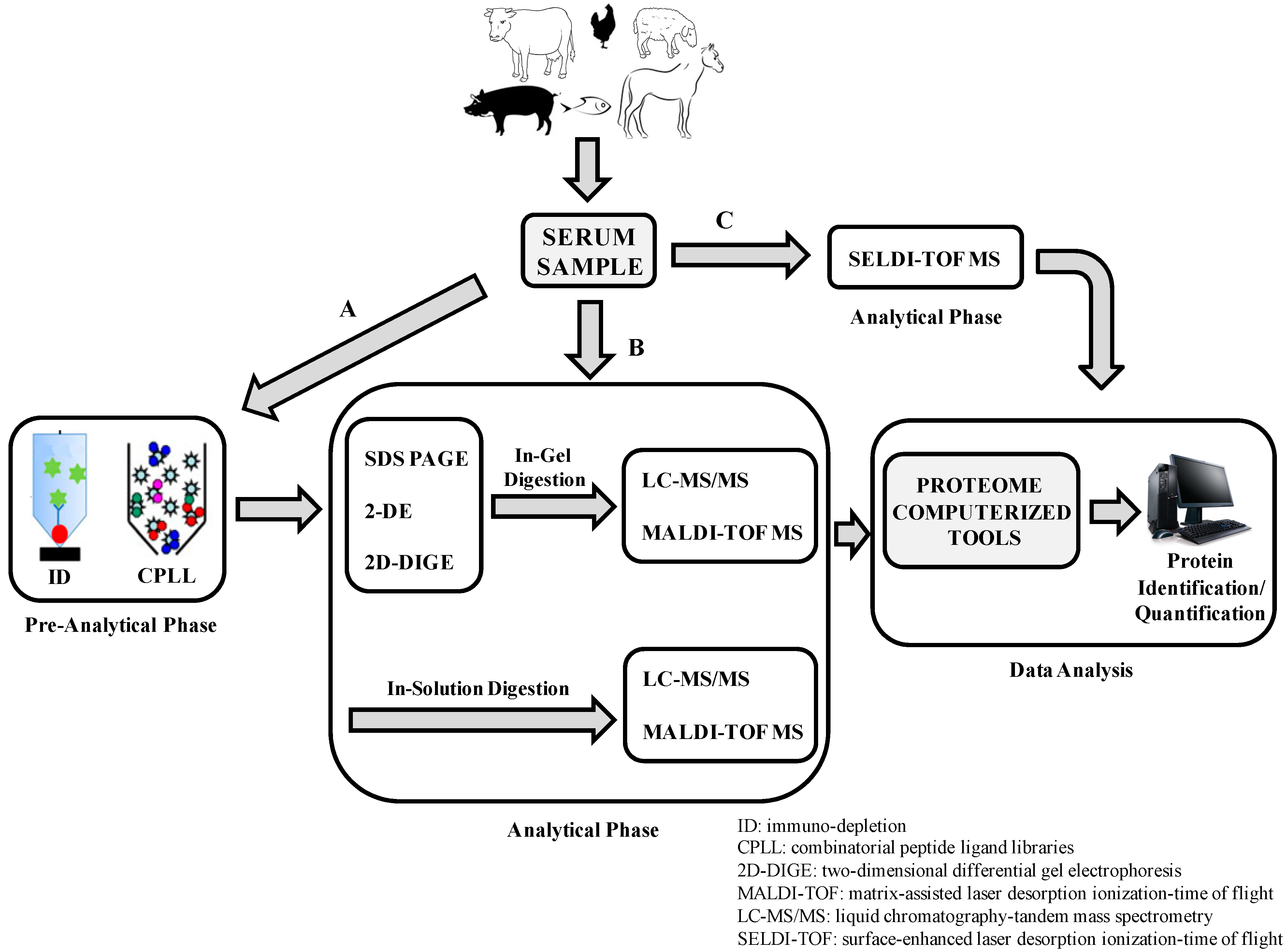
2. Animal Serum Proteomic Applications
| Farm Animals | Proteomic Techniques | Serum Proteomics Study | Reference |
|---|---|---|---|
 Bovine | 2D-DIGE MALDI-TOF-MS MS/MS | Serum proteomic analysis of Bruna cows to identify novel potential biomarkers for stress and welfare | [22] |
| MALDI-TOF-MS | Production of reproducible protein mass profiles able to identify calves undergoing illicit treatments | [23] | |
| 2D-DIGE MALDI-TOF-MS | Investigation of new pathophysiological modulations in the plasma proteome of cows affected by milk fever | [24] | |
| 2-DE MS | Study of serum proteome modulations of heifers during the last phase of pregnancy and early postpartum as a tool for the most accurate management of the peripartum period of these animals | [25] | |
| 1-DE MALDI-TOF-MS | Identification of 480 bovine plasma proteins | [26] | |
| MALDI-TOF-MS 2-DE | Serum proteomics evaluation of cows for a better understanding of mastitis pathophysiology and an early diagnosis of the disease | [27,28] | |
| iTRAQ MS | Study of cattle serum samples infected with either Mycobacterium bovis or Mycobacterium paratuberculosis for discovery of potential biomarkers of infection and progression of disease | [29] | |
| CPLL 2D-DIGE LC–MS/MS | Early diagnosis of bovine Johne’s disease | [30] | |
| MALDI-TOF-MS 2-DE | Proteome and immunome of the tachyzoite stage of Besnoitia besnoiti in bovine besnoitiosis to study significant biological processes and host immune response associated with parasite infection | [31] | |
 Ovine | 2-DE MALDI-TOF-MS | Identification of novel staphylococcal antigens for rational vaccine design | [32] |
| 2-DE | Serological proteome analysis for the identification of Staphylococcus aureus antigens produced in the sheep immune response during mastitis | [33] | |
| SELDI-TOF-MS 1-DE LC–MS/MS | Identification of biomarkers in sheep paratuberculosis | [34] | |
| SELDI-TOF-MS | Detection of a panel of 4 putative serum protein biomarkers as immunological responses in infectious diseases in sheep | [35] | |
| 2-DE MALDI-TOF-MS | Application of proteomics in putative biomarker discovery for early diagnosis as well as for monitoring the physiological and metabolic situations critical for ovine welfare | [36] | |
 Swine | 2D-DIGE MS | Evaluation of the validity of traditional stress biomarkers in pigs housed at different densities and identification of new potential stress biomarkers | [37] |
| 2-D-DIGE MALDI-TOF-MS LC–MS/MS | Identification of 10 differentially expressed protein spots by analysis of the serum proteome modulations in pigs infected by CSF virus versus uninfected controls | [38] | |
| LC–MS/MS | Identification of altered pigs serum proteome with FMD virus infection by analyzing the pigs before and after infection | [39] | |
| LC–MS/MS | Assessment of serum proteome profiles modulation between Ossabaw pigs and human NAFLD | [40,41] | |
 Poultry | 2-DE MALDI-TOF-MS | Analysis of serum proteome of hens at different developmental stages as a base to explore the physiology of growth or reproduction of laying hens | [42] |
| 2-DE LC–MS/MS | Evaluation of APEC proteome changes after exposure to chicken serum to characterize specific protein molecules that may be involved in serum resistance of APEC isolates | [43] | |
| 1-DE MALDI-TOF-MS | Evaluation of APEC growth in different host species requires different survival strategies | [44] | |
| 2-DE, MALDI-TOF-MS | Evaluation of the serum proteome from broilers inoculated with one Eimeria species | [45] | |
 Equine | 1-DE MALDI-TOF-MS MS/MS | Serum proteome evaluation of the effects of Senecio jacobea ingestion by horses | [46] |
| MALDI-TOF-MS LC–MS/MS | Characterization of 29 different serum proteins to establish a horse serum protein database and acquire a better knowledge on equine proteome modulation during diseases | [47] | |
| SELDI-TOF-MS MALDI-TOF-MS | Evaluation of differences in serum proteome profiles of horses, donkeys and mules to determine possible metabolism-related differences | [48] | |
| 2D-DIGE MALDI-TOF-MS | Discovery of 7 potential biomarker candidates to improve diagnosis and therapy of autoimmune uveitis | [49] | |
 Fish | Capillary Electrophoresis–MS | Description of the N-glycans level in serum of salmon (Salmo salar) exposed to long-term handling stress | [50] |
| 2-DE LC–MS/MS | Assessment of the increase of lysozyme and angiotensin carboxypeptidase activities in plasma of fish transferred from hypoosmotic to hyperosmotic solution | [51,52] | |
| 2-DE MALDI-TOF-MS | Determination of protein profiles of alterations in serum of rainbow trout (Oncorhynchus mykiss, Walbaum) as a measure of the acute phase response to the probiotics present in the feed | [53] | |
| 2-DE MS/MS | Characterization of the alterations in serum acute phase response-related proteins with low molecular weight from loach after injury | [54] | |
| 1-DE MALDI-TOF-MS | The first heterogeneous interactome between shrimp serum proteins and Vibrio parahaemolyticus outer membrane proteins | [55] |
2.1. Monitoring of Animal Welfare
2.2. Monitoring of Diseases
2.2.1. Bovine Serum Proteomics
2.2.2. Ovine Serum Proteomics
2.2.3. Swine Serum Proteomics
2.2.4. Poultry Serum Proteomics
2.2.5. Equine Serum Proteomics
2.2.6. Fish Serum Proteomics
3. Conclusions and Future Perspectives
Supplementary Materials
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Doherty, M.K.; Beynon, R.J.; Whitfield, P.D. Proteomics and naturally occurring animal diseases: Opportunities for animal and human medicine. Proteomics Clin. Appl. 2008, 2, 135–141. [Google Scholar] [CrossRef]
- Ceciliani, F.; Eckersall, D.; Burchmore, R.; Lecchi, C. Proteomics in veterinary medicine: Applications and trends in disease pathogenesis and diagnostics. Vet. Pathol. 2014, 51, 351–362. [Google Scholar] [CrossRef]
- Anderson, N.L.; Anderson, N.G. The human plasma proteome: History, character, and diagnostic prospects. Mol. Cell Proteomics 2002, 1, 845–867. [Google Scholar] [CrossRef]
- Introduction to Mass Spectrometry. Available online: http://www.medicine.virginia.edu/research/cores/biomolec/ProteinAnalysisbyMassSpectrometry/Intro%20to%20Mass%20Spec%20Course%20UVa%2020100518.pdf (accessed on 1 September 2014).
- Fang, X.; Zhang, W.W. Affinity separation and enrichment methods in proteomic analysis. J. Proteomics 2008, 71, 284–303. [Google Scholar] [CrossRef]
- Righetti, P.G.; Boschetti, E. The ProteoMiner and the FortyNiners: Searching for gold nuggets in the proteomic arena. Mass Spec. Rev. 2008, 27, 596–608. [Google Scholar] [CrossRef]
- Boschetti, E.; Lomas, L.; Citterio, A.; Righetti, P.G. Romancing the “hidden proteome”, anno domini two zero zero seven. J. Chromatogr. A 2007, 1153, 277–290. [Google Scholar] [CrossRef]
- Boschetti, E.; Righetti, P.G. The art of observing rare protein species in proteomes with peptide ligand libraries. Proteomics 2009, 9, 1492–1510. [Google Scholar] [CrossRef]
- Gundry, R.L.; White, M.Y.; Nogee, J.; Tchernyshyov, I.; van Eyk, J.E. Assessment of albumin removal from an immunoaffinity spin column: Critical implications for proteomic examination of the albuminome and albumin-depleted samples. Proteomics 2009, 9, 2021–2028. [Google Scholar] [CrossRef]
- Zhi, W.; Purohit, S.; Carey, C.; Wang, M.; She, J.X. Proteomic technologies for the discovery of type 1 diabetes biomarkers. J. Diabetes Sci. Technol. 2010, 4, 993–1002. [Google Scholar] [CrossRef]
- Walther, T.C.; Mann, M. Mass spectrometry-based proteomics in cell biology. J. Cell Biol. 2010, 190, 491–500. [Google Scholar] [CrossRef]
- Pan, C.; Xu, S.; Zhou, H.; Fu, Y.; Ye, M.; Zou, H. Recent developments in methods and technology for analysis of biological samples by MALDI-TOF-MS. Anal. Bioanal. Chem. 2007, 387, 193–204. [Google Scholar]
- Rodríguez-Suárez, E.; Whetton, A.D. The application of quantification techniques in proteomics for biomedical research. Mass Spectrom. Rev. 2013, 32, 1–26. [Google Scholar] [CrossRef]
- Unlü, M.; Morgan, M.E.; Minden, J.S. Difference gel electrophoresis: A single gel method for detecting changes in protein extracts. Electrophoresis 1997, 18, 2071–2077. [Google Scholar] [CrossRef]
- Gygi, S.P.; Rist, B.; Gerber, S.A.; Turecek, F.; Gelb, M.H.; Aebersold, R. Quantitative analysis of complex protein mixtures using isotope-coded affinity tags. Nat. Biotechnol. 1999, 17, 994–999. [Google Scholar] [CrossRef]
- Ross, P.L.; Huang, Y.N.; Marchese, J.N.; Williamson, B.; Parker, K.; Hattan, S.; Khainovski, N.; Pillai, S.; Dey, S.; Daniels, S.; et al. Multiplexed protein quantitation in Saccharomyces cerevisiae using amine-reactive isobaric tagging reagents. Mol. Cell. Proteomics 2004, 3, 1154–1169. [Google Scholar] [CrossRef]
- Old, W.M.; Meyer-Arendt, K.; Aveline-Wolf, L.; Pierce, K.G.; Mendoza, A.; Sevinsky, J.R.; Resing, K.A.; Ahn, N.G. Comparison of label-free methods for quantifying human proteins by shotgun proteomics. Mol. Cell Proteomics 2005, 4, 1487–1502. [Google Scholar] [CrossRef]
- Issaq, H.J.; Veenstra, T.D.; Conrads, T.P.; Felschow, D. The SELDI-TOF MS approach to proteomics: Protein profiling and biomarker identification. Biochem. Biophys. Res. Commun. 2002, 292, 587–592. [Google Scholar] [CrossRef]
- UniProt-GOA. Available online: http://www.ebi.ac.uk/GOA/newto (accessed on 27 August 2014).
- Balakrishnan, R.; Harris, M.A.; Huntley, R.; van Auken, K.; Cherry, J.M. A guide to best practices for Gene Ontology (GO) manual annotation. Database 2013, 9, bat054. [Google Scholar]
- Gene Ontology Consortium. Available online: http://beta.geneontology.org/ (accessed on 27 August 2014).
- Marco-Ramell, A.; Arroyo, L.; Saco, Y.; García-Heredia, A.; Camps, J.; Fina, M.; Piedrafita, J.; Bassols, A.J. Proteomic analysis reveals oxidative stress response as the main adaptative physiological mechanism in cows under different production systems. Proteomics 2012, 75, 4399–4411. [Google Scholar] [CrossRef]
- Della Donna, L.; Ronci, M.; Sacchetta, P.; Di Ilio, C.; Biolatti, B.; Federici, G.; Nebbia, C.; Urbani, A. A food safety control low mass-range proteomics platform for the detection of illicit treatments in veal calves by MALDI-TOF-MS serum profiling. J. Biotechnol. 2009, 4, 1596–1609. [Google Scholar] [CrossRef]
- Xia, C.; Zhang, H.Y.; Wu, L.; Xu, C.; Zheng, J.S.; Yan, Y.J.; Yang, L.J.; Shu, S. Proteomic analysis of plasma from cows affected with milk fever using two-dimensional differential in-gel electrophoresis and mass spectrometry. Res. Vet. Sci. 2012, 93, 857–861. [Google Scholar] [CrossRef]
- Cairoli, F.; Battocchio, M.; Veronesi, M.C.; Brambilla, D.; Conserva, F.; Eberini, I.; Wait, R.; Gianazza, E. Serum protein pattern during cow pregnancy: Acute-phase proteins increase in the peripartum period. Electrophoresis 2006, 27, 1617–1625. [Google Scholar] [CrossRef]
- Henning, A.K.; Groschup, M.H.; Mettenleiter, T.C.; Karger, A. Analysis of the bovine plasma proteome by matrix-assisted laser desorption/ionisation time-of-flight tandem mass spectrometry. Vet. J. 2014, 199, 175–80. [Google Scholar] [CrossRef]
- Turk, R.; Piras, C.; Kovačić, M.; Samardžija, M.; Ahmed, H.; de Canio, M.; Urbani, A.; Meštrić, Z.F.; Soggiu, A.; Bonizzi, L.; et al. Proteomics of inflammatory and oxidative stress response in cows with subclinical and clinical mastitis. Proteomics 2012, 75, 4412–4428. [Google Scholar] [CrossRef]
- Alonso-Fauste, I.; Andrés, M.; Iturralde, M.; Lampreave, F.; Gallart, J.; Alava, M.A.J. Proteomic characterization by 2-DE in bovine serum and whey from healthy and mastitis affected farm animals. Proteomics 2012, 75, 3015–3030. [Google Scholar]
- Seth., M.; Lamont, E.A.; Janagama, H.K.; Widdel, A.; Vulchanova, L.; Stabel, J.R.; Waters, W.R.; Palmer, M.V.; Sreevatsan, S. Biomarker discovery in subclinical mycobacterial infections of cattle. PLoS One 2009, 4, e5478. [Google Scholar] [CrossRef]
- You, Q.; Verschoor, C.P.; Pant, S.D.; Macri, J.; Kirby, G.M.; Karrow, N.A. Proteomic analysis of plasma from Holstein cows testing positive for Mycobacterium avium subsp. paratuberculosis (MAP). Vet. Immunol. Immunopathol. 2012, 148, 243–251. [Google Scholar]
- García-Lunar, P.; Regidor-Cerrillo, J.; Gutiérrez-Expósito, D.; Ortega-Mora, L.; Alvarez-García, G. First 2-DE approach towards characterising the proteome and immunome of Besnoitia besnoiti in the tachyzoite stage. Vet. Parasitol. 2013, 195, 24–34. [Google Scholar] [CrossRef]
- Vytvytska, O.; Nagy, E.; Blüggel, M.; Meyer, H.E.; Kurzbauer, R.; Huber, L.A.; Klade, C.S. Identification of vaccine candidate antigens of Staphylococcus aureus by serological proteome analysis. Proteomics 2002, 2, 580–590. [Google Scholar] [CrossRef]
- Le Maréchal, C.; Jan, G.; Even, S.; McCulloch, J.A.; Azevedo, V.; Thiéry, R.; Vautor, E.; le Loir, Y.J. Development of serological proteome analysis of mastitis by Staphylococcus aureus in ewes. Methods Microbiol. 2009, 79, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.; Taylor, D.; Begg, D.J.; Whittington, R.J. Biomarker discovery for ovine paratuberculosis (Johne’s disease) by proteomic serum profiling. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 315–326. [Google Scholar] [CrossRef]
- Zhong, L.; Taylor, D.L.; Whittington, R.J. Proteomic profiling of ovine serum by SELDI-TOF MS: Optimisation, reproducibility and feasibility of biomarker discovery using routinely collected samples. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 47–63. [Google Scholar] [CrossRef]
- Chiaradia, E.; Avellini, L.; Tartaglia, M.; Gaiti, A.; Just, I.; Scoppetta, F.; Czentnar, Z.; Pich, A. Proteomic evaluation of sheep serum proteins. BMC Vet. Res. 2012, 25, 66. [Google Scholar]
- Marco-Ramell, A.; Pato, R.; Peña, R.; Saco, Y.; Manteca, X.; Ruiz de la Torre, J.L.; Bassols, A. Identification of serum stress biomarkers in pigs housed at different stocking densities. Vet. J. 2011, 190, e66–e71. [Google Scholar] [CrossRef]
- Sun, J.F.; Shi, Z.X.; Guo, H.C.; Li, S.; Tu, C.C. Proteomic analysis of swine serum following highly virulent classical swine fever virus infection. Virol. J. 2011, 8, 107. [Google Scholar]
- Liu, Y.; Zhang, K.; Zheng, H.; Shang, Y.; Guo, J.; Tian, H.; Lu, G.; Jin, Y.; He, J.; Cai, X.; et al. Proteomics analysis of porcine serum proteins by LC–MS/MS after foot-and-mouth disease virus (FMDV) infection. J. Vet. Med. Sci. 2011, 73, 1569–1572. [Google Scholar] [CrossRef]
- Lee, L.; Alloosh, M.; Saxena, R.; van Alstine, W.; Watkins, B.A.; Klaunig, J.E.; Sturek, M.; Chalasani, N. Nutritional model of steatohepatitis and metabolic syndrome in the Ossabaw miniature swine. Hepatology 2009, 50, 56–67. [Google Scholar]
- Bell, L.N.; Lee, L.; Saxena, R.; Bemis, K.G.; Wang, M.; Theodorakis, J.L.; Vuppalanchi, R.; Alloosh, M.; Sturek, M.; Chalasani, N. Serum proteomic analysis of diet-induced steatohepatitis and metabolic syndrome in the Ossabaw miniature swine. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, 746–754. [Google Scholar] [CrossRef]
- Huang, S.Y.; Lin, J.H.; Chen, Y.H.; Chuang, C.K.; Chiu, Y.F.; Chen, M.Y.; Chen, H.H.; Lee, W.C. Analysis of chicken serum proteome and differential protein expression during development in single-comb White Leghorn hens. Proteomics 2006, 6, 2217–2224. [Google Scholar] [CrossRef]
- Tyler, C.D.; Lichti, C.F.; Diekman, A.B.; Foley, S.L. Evaluation of differentially expressed proteins following serum exposure in avian pathogenic Escherichia coli. Avian Dis. 2008, 52, 23–27. [Google Scholar]
- Li, G.; Cai, W.; Hussein, A.; Wannemuehler, Y.M.; Logue, C.M.; Nolan, L.K.J. Proteome response of an extraintestinal pathogenic Escherichia coli strain with zoonotic potential to human and chicken sera. Proteomics 2012, 75, 4853–4862. [Google Scholar] [CrossRef]
- Gilbert, E.R.; Cox, C.M.; Williams, P.M.; McElroy, A.P.; Dalloul, R.A.; Ray, W.K.; Barri, A.; Emmerson, D.A.; Wong, E.A.; Webb, K.E. Eimeria species and genetic background influence the serum protein profile of broilers with coccidiosis. PLoS One 2011, 6, e14636. [Google Scholar] [CrossRef]
- Moore, R.E.; Knottenbelt, D.; Matthews, J.B.; Beynon, R.J.; Whitfield, P.D. Biomarkers for ragwort poisoning in horses: Identification of protein targets. BMC Vet. Res. 2008, 8, 4–30. [Google Scholar]
- Miller, I.; Friedlein, A.; Tsangaris, G.; Maris, A.; Fountoulakis, M.; Gemeiner, M. The serum proteome of Equus caballus. Proteomics 2004, 4, 3227–3234. [Google Scholar] [CrossRef]
- Henze, A.; Aumer, F.; Grabner, A.; Raila, J.; Schweigert, F.J. Genetic differences in the serum proteome of horses, donkeys and mules are detectable by protein profiling. J. Nutr. 2011, 1, 170–173. [Google Scholar]
- Zipplies, J.K.; Hauck, S.M.; Schoeffmann, S.; Amann, B.; Stangassinger, M.; Ueffing, M.; Deeg, C.A. Serum PEDF levels are decreased in a spontaneous animal model for human autoimmune uveitis. J. Proteome Res. 2009, 8, 992–998. [Google Scholar] [CrossRef]
- Liu, X.; Afonso, L.; Altman, E.; Johnson, S.; Brown, L.; Li, J. O-acetylation of sialic acids in N-glycans of Atlantic salmon (Salmo salar) serum is altered by handling stress. Proteomics 2008, 8, 2849–2857. [Google Scholar] [CrossRef]
- Jiang, I.F.; Kumar, V.B.; Weng, C.F. Acute osmotic stress affects tilapia (Oreochromis mossambicus) innate immune responses. Fish Shellfish Immunol. 2008, 25, 841–846. [Google Scholar] [CrossRef]
- Kumar, V.B.; Jiang, I.F.; Yang, H.H.; Weng, C.F. Effects of serum on phagocytic activity and proteomic analysis of tilapia (Oreochromis mossambicus) serum after acute osmotic stress. Fish Shellfish Immunol. 2009, 26, 760–767. [Google Scholar] [CrossRef]
- Brunt, J.; Hansen, R.; Jamieson, D.J.; Austin, B. Proteomic analysis of rainbow trout (Oncorhynchus mykiss, Walbaum) serum after administration of probiotics in diets. Vet. Immunol. Immunopathol. 2008, 121, 199–205. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, S.; Peng, X. Serum acute phase response (APR)-related proteome of loach to trauma. Fish Shellfish Immunol. 2004, 16, 381–389. [Google Scholar]
- Liu, X.; She, X.T.; Zhu, Q.F.; Li, H.; Peng, X.X. Heterogeneous interactome between Litopenaeus vannamei plasma proteins and Vibrio parahaemolyticus outer membrane proteins. Fish Shellfish Immunol. 2013, 34, 192–198. [Google Scholar] [CrossRef]
- Kornegay, E.T.; Nolter, D.R. Effect of floor space and number of pigs per pen on performance. Pig News Inf. 1984, 5, 23–33. [Google Scholar]
- Kuhlers, D.L.; Jungst, S.B.; Marple, D.N.; Rahe, C.H. The effect of pen density during rearing on subsequent reproductive performance in gilts. J. Anim. Sci. 1985, 61, 1066–1069. [Google Scholar]
- Meunier-Salaun, M.C.; Vantrimponte, M.N.; Raab, A.; Dantzer, R. Effect of floor area restriction upon performance, behavior and physiology of growing-finishing pigs. J. Anim. Sci. 1987, 64, 1371–1377. [Google Scholar]
- Angulo, P. Nonalcoholic fatty liver disease. N. Engl. J. Med. 2002, 346, 1221–1231. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Di Girolamo, F.; D'Amato, A.; Lante, I.; Signore, F.; Muraca, M.; Putignani, L. Farm Animal Serum Proteomics and Impact on Human Health. Int. J. Mol. Sci. 2014, 15, 15396-15411. https://doi.org/10.3390/ijms150915396
Di Girolamo F, D'Amato A, Lante I, Signore F, Muraca M, Putignani L. Farm Animal Serum Proteomics and Impact on Human Health. International Journal of Molecular Sciences. 2014; 15(9):15396-15411. https://doi.org/10.3390/ijms150915396
Chicago/Turabian StyleDi Girolamo, Francesco, Alfonsina D'Amato, Isabella Lante, Fabrizio Signore, Marta Muraca, and Lorenza Putignani. 2014. "Farm Animal Serum Proteomics and Impact on Human Health" International Journal of Molecular Sciences 15, no. 9: 15396-15411. https://doi.org/10.3390/ijms150915396