
Transmembrane Signal Transduction in Oocyte Maturation and Fertilization: Focusing on Xenopus laevis as a Model Animal

{kind=link}
Abstract
:1. Oocyte “Cytoplasmic” Signaling Events Associated with Meiotic Maturation and Fertilization, Focusing on Protein Phosphorylation
1.1. Meiosis and Oocyte Cytoplasmic Maturation

1.2. Maturation-Promoting Factor
1.3. Cyclic Nucleotides and Oocyte Cytoplasmic Maturation
1.4. Mitogen-Activated Protein Kinase Cascade
1.5. Cytostatic Factor and Secondary Cell Cycle Arrest in Meiotic Oocyte Maturation
1.6. Sperm-Induced Resumption of Meiosis and Oocyte/Egg Activation
2. Plasma Membrane- and/or Egg Coat-Associated Events during Oocyte Maturation, Ovulation, and Fertilization
2.1. Mechanism of Progesterone and Its Receptor Interaction for Oocyte Maturation
2.2. Structure and Function of Vitelline Envelope/Membrane, and Other Egg Coats
2.3. Sperm Plasma Membrane-Associated Components
2.4. Egg/Oocyte Plasma Membrane-Associated Components
2.5. Physiological Function of Egg MD (Microdomain) and Its Acquisition during Oocyte Maturation
3. Future Perspective
Acknowledgments
Conflicts of Interest
References
- Kang, M.K.; Han, S.J. Post-transcriptional and post-translational regulation during mouse oocyte maturation. BMB Rep. 2011, 44, 147–157. [Google Scholar]
- Liang, C.G.; Su, Y.Q.; Fan, H.Y.; Schatten, H.; Sun, Q.Y. Mechanisms regulating oocyte meiotic resumption: Roles of mitogen-activated protein kinase. Mol. Endocrinol. 2007, 21, 2037–2055. [Google Scholar]
- Machaca, K. Ca2+ signaling differentiation during oocyte maturation. J. Cell Physiol. 2007, 213, 331–340. [Google Scholar]
- Madgwick, S.; Jones, K.T. How eggs arrest at metaphase II: MPF stabilisation plus APC/C inhibition equals Cytostatic Factor. Cell Div. 2007, 2. [Google Scholar] [CrossRef]
- Schmitt, A.; Nebreda, A.R. Signalling pathways in oocyte meiotic maturation. J. Cell Sci. 2002, 115, 2457–2459. [Google Scholar]
- Tripathi, A.; Kumar, K.V.; Chaube, S.K. Meiotic cell cycle arrest in mammalian oocytes. J. Cell. Physiol. 2010, 223, 592–600. [Google Scholar]
- Ferrell, J.E., Jr. Xenopus oocyte maturation: New lessons from a good egg. Bioessays 1999, 21, 833–842. [Google Scholar]
- Tunquist, B.J.; Maller, J.L. Under arrest: Cytostatic factor (CSF)-mediated metaphase arrest in vertebrate eggs. Genes Dev. 2003, 17, 683–710. [Google Scholar]
- Mehlmann, L.M. Stops and starts in mammalian oocytes: Recent advances in understanding the regulation of meiotic arrest and oocyte maturation. Reproduction 2005, 130, 791–799. [Google Scholar]
- Sirard, M.A. Resumption of meiosis: Mechanism involved in meiotic progression and its relation with developmental competence. Theriogenology 2001, 55, 1241–1254. [Google Scholar]
- Trounson, A.; Anderiesz, C.; Jones, G. Maturation of human oocytes in vitro and their developmental competence. Reproduction 2001, 121, 51–75. [Google Scholar]
- Wassmann, K.; Niault, T.; Maro, B. Metaphase I arrest upon activation of the Mad2-dependent spindle checkpoint in mouse oocytes. Curr. Biol. 2003, 13, 1596–1608. [Google Scholar]
- Maller, J.L.; Gautier, J.; Langan, T.A.; Lohka, M.J.; Shenoy, S.; Shalloway, D.; Nurse, P. Maturation-promoting factor and the regulation of the cell cycle. J. Cell Sci. Suppl. 1989, 12, 53–63. [Google Scholar]
- Maller, J.L. Xenopus oocytes and the biochemistry of cell division. Biochemistry 1990, 29, 3157–3166. [Google Scholar]
- Masui, Y. Towards understanding the control of the division cycle in animal cells. Biochem. Cell Biol. 1992, 70, 920–945. [Google Scholar]
- Clarke, P.R.; Karsenti, E. Regulation of p34cdc2 protein kinase: New insights into protein phosphorylation and the cell cycle. J. Cell Sci. 1991, 100, 409–414. [Google Scholar]
- Ledan, E.; Polanski, Z.; Terret, M.E.; Maro, B. Meiotic maturation of the mouse oocyte requires an equilibrium between cyclin B synthesis and degradation. Dev. Biol. 2001, 232, 400–413. [Google Scholar]
- Marangos, P.; Verschuren, E.W.; Chen, R.; Jackson, P.K.; Carroll, J. Prophase I arrest and progression to metaphase I in mouse oocytes are controlled by Emi1-dependent regulation of APCCdh1. J. Cell Biol. 2007, 176, 65–75. [Google Scholar]
- Norris, R.P.; Ratzan, W.J.; Freudzon, M.; Mehlmann, L.M.; Krall, J.; Movsesian, M.A.; Wang, H.; Ke, H.; Nikolaev, V.O.; Jaffe, L.A. Cyclic GMP from the surrounding somatic cells regulates cyclic AMP and meiosis in the mouse oocyte. Development 2009, 136, 1869–1878. [Google Scholar]
- Sirard, M.A.; Bilodeau, S. Granulosa cells inhibit the resumption of meiosis in bovine oocytes in vitro. Biol. Reprod. 1990, 43, 777–783. [Google Scholar]
- Sun, Q.Y.; Miao, Y.L.; Schatten, H. Towards a new understanding on the regulation of mammalian oocyte meiosis resumption. Cell Cycle 2009, 8, 2741–2747. [Google Scholar]
- Vaccari, S.; Horner, K.; Mehlmann, L.M.; Conti, M. Generation of mouse oocytes defective in cAMP synthesis and degradation: Endogenous cyclic AMP is essential for meiotic arrest. Dev. Biol. 2008, 316, 124–134. [Google Scholar]
- Mayes, M.A.; Sirard, M.A. Effect of type 3 and type 4 phosphodiesterase inhibitors on the maintenance of bovine oocytes in meiotic arrest. Biol. Reprod. 2002, 66, 180–184. [Google Scholar]
- Tsafriri, A.; Chun, S.Y.; Zhang, R.; Hsueh, A.J.; Conti, M. Oocyte maturation involves compartmentalization and opposing changes of cAMP levels in follicular somatic and germ cells: Studies using selective phosphodiesterase inhibitors. Dev. Biol. 1996, 178, 393–402. [Google Scholar]
- Pirino, G.; Wescott, M.P.; Donovan, P.J. Protein kinase A regulates resumption of meiosis by phosphorylation of Cdc25B in mammalian oocytes. Cell Cycle 2009, 8, 665–670. [Google Scholar]
- Han, S.J.; Conti, M. New pathways from PKA to the Cdc2/cyclin B complex in oocytes: Wee1B as a potential PKA substrate. Cell Cycle 2006, 5, 227–231. [Google Scholar]
- Stanford, J.S.; Ruderman, J.V. Changes in regulatory phosphorylation of Cdc25C Ser287 and Wee1 Ser549 during normal cell cycle progression and checkpoint arrests. Mol. Biol. Cell 2005, 16, 5749–5760. [Google Scholar]
- Solc, P.; Schultz, R.M.; Motlik, J. Prophase I arrest and progression to metaphase I in mouse oocytes: Comparison of resumption of meiosis and recovery from G2-arrest in somatic cells. Mol. Hum. Reprod. 2010, 16, 654–664. [Google Scholar]
- Potapova, T.A.; Daum, J.R.; Byrd, K.S.; Gorbsky, G.J. Fine tuning the cell cycle: Activation of the Cdk1 inhibitory phosphorylation pathway during mitotic exit. Mol. Biol. Cell 2009, 20, 1737–1748. [Google Scholar]
- Zhang, M.; Ouyang, H.; Xia, G. The signal pathway of gonadotrophins-induced mammalian oocyte meiotic resumption. Mol. Hum. Reprod. 2009, 15, 399–409. [Google Scholar]
- Tornell, J.; Billig, H.; Hillensjo, T. Regulation of oocyte maturation by changes in ovarian levels of cyclic nucleotides. Hum. Reprod. 1991, 6, 411–422. [Google Scholar]
- Wang, S.; Ning, G.; Chen, X.; Yang, J.; Ouyang, H.; Zhang, H.; Tai, P.; Mu, X.; Zhou, B.; Zhang, M.; et al. PDE5 modulates oocyte spontaneous maturation via cGMP-cAMP but not cGMP-PKG signaling. Front. Biosci. 2008, 13, 7087–7095. [Google Scholar]
- Mehlmann, L.M.; Jones, T.L.; Jaffe, L.A. Meiotic arrest in the mouse follicle maintained by a Gs protein in the oocyte. Science 2002, 297, 1343–1345. [Google Scholar]
- Conti, M.; Andersen, C.B.; Richard, F.; Mehats, C.; Chun, S.Y.; Horner, K.; Jin, C.; Tsafriri, A. Role of cyclic nucleotide signaling in oocyte maturation. Mol. Cell Endocrinol. 2002, 187, 153–159. [Google Scholar]
- Hupalowska, A.; Kalaszczynska, I.; Hoffmann, S.; Tsurumi, C.; Kubiak, J.Z.; Polanski, Z.; Ciemerych, M.A. Metaphase I arrest in LT/Sv mouse oocytes involves the spindle assembly checkpoint. Biol. Reprod. 2008, 79, 1102–1110. [Google Scholar]
- Li, M.; Li, S.; Yuan, J.; Wang, Z.B.; Sun, S.C.; Schatten, H.; Sun, Q.Y. Bub3 is a spindle assembly checkpoint protein regulating chromosome segregation during mouse oocyte meiosis. PLoS One 2009, 4, e7701. [Google Scholar]
- Niault, T.; Hached, K.; Sotillo, R.; Sorger, P.K.; Maro, B.; Benezra, R.; Wassmann, K. Changing Mad2 levels affects chromosome segregation and spindle assembly checkpoint control in female mouse meiosis I. PLoS One 2007, 2, e1165. [Google Scholar]
- Brunet, S.; Maro, B. Cytoskeleton and cell cycle control during meiotic maturation of the mouse oocyte: Integrating time and space. Reproduction 2005, 130, 801–811. [Google Scholar]
- Homer, H. New insights into the genetic regulation of homologue disjunction in mammalian oocytes. Cytogenet. Genome Res. 2011, 133, 209–222. [Google Scholar]
- Madgwick, S.; Nixon, V.L.; Chang, H.Y.; Herbert, M.; Levasseur, M.; Jones, K.T. Maintenance of sister chromatid attachment in mouse eggs through maturation-promoting factor activity. Dev. Biol. 2004, 275, 68–81. [Google Scholar]
- Choi, T.; Rulong, S.; Resau, J.; Fukasawa, K.; Matten, W.; Kuriyama, R.; Mansour, S.; Ahn, N.; Vande Woude, G.F. Mos/mitogen-activated protein kinase can induce early meiotic phenotypes in the absence of maturation-promoting factor: A novel system for analyzing spindle formation during meiosis I. Proc. Natl. Acad. Sci. USA 1996, 93, 4730–4735. [Google Scholar]
- Verlhac, M.H.; Kubiak, J.Z.; Weber, M.; Geraud, G.; Colledge, W.H.; Evans, M.J.; Maro, B. Mos is required for MAP kinase activation and is involved in microtubule organization during meiotic maturation in the mouse. Development 1996, 122, 815–822. [Google Scholar]
- Zhou, R.P.; Oskarsson, M.; Paules, R.S.; Schulz, N.; Cleveland, D.; Vande Woude, G.F. Ability of the c-mos product to associate with and phosphorylate tubulin. Science 1991, 251, 671–675. [Google Scholar]
- Masui, Y.; Markert, C.L. Cytoplasmic control of nuclear behavior during meiotic maturation of frog oocytes. J. Exp. Zool. 1971, 177, 129–145. [Google Scholar]
- Wu, J.Q.; Kornbluth, S. Across the meiotic divide—CSF activity in the post-Emi2/XErp1 era. J. Cell Sci. 2008, 121, 3509–3514. [Google Scholar]
- Perry, A.C.; Verlhac, M.H. Second meiotic arrest and exit in frogs and mice. EMBO Rep. 2008, 9, 246–251. [Google Scholar]
- Shoji, S.; Yoshida, N.; Amanai, M.; Ohgishi, M.; Fukui, T.; Fujimoto, S.; Nakano, Y.; Kajikawa, E.; Perry, A.C. Mammalian Emi2 mediates cytostatic arrest and transduces the signal for meiotic exit via Cdc20. EMBO J. 2006, 25, 834–845. [Google Scholar]
- Schmidt, A.; Rauh, N.R.; Nigg, E.A.; Mayer, T.U. Cytostatic factor: An activity that puts the cell cycle on hold. J. Cell Sci. 2006, 119, 1213–1218. [Google Scholar]
- Tang, W.; Wu, J.Q.; Guo, Y.; Hansen, D.V.; Perry, J.A.; Freel, C.D.; Nutt, L.; Jackson, P.K.; Kornbluth, S. Cdc2 and Mos regulate Emi2 stability to promote the meiosis I-meiosis II transition. Mol. Biol. Cell 2008, 19, 3536–3543. [Google Scholar]
- Ohsumi, K.; Koyanagi, A.; Yamamoto, T.M.; Gotoh, T.; Kishimoto, T. Emi1-mediated M-phase arrest in Xenopus eggs is distinct from cytostatic factor arrest. Proc. Natl. Acad. Sci. USA 2004, 101, 12531–12536. [Google Scholar]
- Inoue, D.; Ohe, M.; Kanemori, Y.; Nobui, T.; Sagata, N. A direct link of the Mos-MAPK pathway to Erp1/Emi2 in meiotic arrest of Xenopus laevis eggs. Nature 2007, 446, 1100–1104. [Google Scholar]
- Nishiyama, T.; Ohsumi, K.; Kishimoto, T. Phosphorylation of Erp1 by p90rsk is required for cytostatic factor arrest in Xenopus laevis eggs. Nature 2007, 446, 1096–1099. [Google Scholar]
- Mochida, S.; Hunt, T. Calcineurin is required to release Xenopus egg extracts from meiotic M phase. Nature 2007, 449, 336–340. [Google Scholar]
- Nishiyama, T.; Yoshizaki, N.; Kishimoto, T.; Ohsumi, K. Transient activation of calcineurin is essential to initiate embryonic development in Xenopus laevis. Nature 2007, 449, 341–345. [Google Scholar]
- Hansen, D.V.; Tung, J.J.; Jackson, P.K. CaMKII and polo-like kinase 1 sequentially phosphorylate the cytostatic factor Emi2/XErp1 to trigger its destruction and meiotic exit. Proc. Natl. Acad. Sci. USA 2006, 103, 608–613. [Google Scholar]
- Liu, J.; Maller, J.L. Calcium elevation at fertilization coordinates phosphorylation of XErp1/Emi2 by Plx1 and CaMKII to release metaphase arrest by cytostatic factor. Curr. Biol. 2005, 15, 1458–1468. [Google Scholar]
- Rauh, N.R.; Schmidt, A.; Bormann, J.; Nigg, E.A.; Mayer, T.U. Calcium triggers exit from meiosis II by targeting the APC/C inhibitor XErp1 for degradation. Nature 2005, 437, 1048–1052. [Google Scholar]
- Liu, J.; Grimison, B.; Lewellyn, A.L.; Maller, J.L. The anaphase-promoting complex/cyclosome inhibitor Emi2 is essential for meiotic but not mitotic cell cycles. J. Biol. Chem. 2006, 281, 34736–34741. [Google Scholar]
- Castro, A.; Peter, M.; Magnaghi-Jaulin, L.; Vigneron, S.; Galas, S.; Lorca, T.; Labbe, J.C. Cyclin B/cdc2 induces c-Mos stability by direct phosphorylation in Xenopus oocytes. Mol. Biol. Cell 2001, 12, 2660–2671. [Google Scholar]
- Madgwick, S.; Hansen, D.V.; Levasseur, M.; Jackson, P.K.; Jones, K.T. Mouse Emi2 is required to enter meiosis II by reestablishing cyclin B1 during interkinesis. J. Cell Biol. 2006, 174, 791–801. [Google Scholar]
- Maller, J.L.; Schwab, M.S.; Roberts, B.T.; Gross, S.D.; Taieb, F.E.; Tunquist, B.J. The pathway of MAP kinase mediation of CSF arrest in Xenopus oocytes. Biol. Cell 2001, 93, 27–33. [Google Scholar]
- Ducibella, T. The cortical reaction and development of activation competence in mammalian oocytes. Hum. Reprod. Update 1996, 2, 29–42. [Google Scholar]
- Ducibella, T.; Fissore, R. The roles of Ca2+, downstream protein kinases, and oscillatory signaling in regulating fertilization and the activation of development. Dev. Biol. 2008, 315, 257–279. [Google Scholar]
- Schultz, R.M.; Kopf, G.S. Molecular basis of mammalian egg activation. Curr. Top. Dev. Biol. 1995, 30, 21–62. [Google Scholar]
- Gilkey, J.C.; Jaffe, L.F.; Ridgway, E.B.; Reynolds, G.T. A free calcium wave traverses the activating egg of the medaka, Oryzias latipes. J. Cell Biol. 1978, 76, 448–466. [Google Scholar]
- Miyazaki, S.; Ito, M. Calcium signals for egg activation in mammals. J. Pharmacol. Sci. 2006, 100, 545–552. [Google Scholar]
- Runft, L.L.; Jaffe, L.A.; Mehlmann, L.M. Egg activation at fertilization: Where it all begins. Dev. Biol. 2002, 245, 237–254. [Google Scholar]
- Steinhardt, R.A.; Epel, D. Activation of sea-urchin eggs by a calcium ionophore. Proc. Natl. Acad. Sci. USA 1974, 71, 1915–1919. [Google Scholar]
- Stricker, S.A. Comparative biology of calcium signaling during fertilization and egg activation in animals. Dev. Biol. 1999, 211, 157–176. [Google Scholar]
- Whitaker, M. Calcium at fertilization and in early development. Physiol. Rev. 2006, 86, 25–88. [Google Scholar]
- Mazia, D. The release of calcium in Arbacia eggs upon fertilization. J. Cell Comp. Phys. 1937, 10, 291–304. [Google Scholar]
- Ajduk, A.; Malagocki, A.; Maleszewski, M. Cytoplasmic maturation of mammalian oocytes: Development of a mechanism responsible for sperm-induced Ca2+ oscillations. Reprod. Biol. 2008, 8, 3–22. [Google Scholar]
- Horner, V.L.; Wolfner, M.F. Transitioning from egg to embryo: Triggers and mechanisms of egg activation. Dev. Dyn. 2008, 237, 527–544. [Google Scholar]
- Swann, K.; Saunders, C.M.; Rogers, N.T.; Lai, F.A. PLCz: A sperm protein that triggers Ca2+ oscillations and egg activation in mammals. Semin. Cell Dev. Biol. 2006, 17, 264–273. [Google Scholar]
- Townley, I.K.; Roux, M.M.; Foltz, K.R. Signal transduction at fertilization: The Ca2+ release pathway in echinoderms and other invertebrate deuterostomes. Semin. Cell Dev. Biol. 2006, 17, 293–302. [Google Scholar]
- Chaube, S.K.; Dubey, P.K.; Mishra, S.K.; Shrivastav, T.G. Verapamil reversibly inhibits spontaneous parthenogenetic activation in aged rat eggs cultured in vitro. Cloning Stem Cells 2007, 9, 608–617. [Google Scholar]
- Galat, V.; Zhou, Y.; Taborn, G.; Garton, R.; Iannaccone, P. Overcoming MIII arrest from spontaneous activation in cultured rat oocytes. Cloning Stem Cells 2007, 9, 303–314. [Google Scholar]
- Vincent, C.; Cheek, T.R.; Johnson, M.H. Cell cycle progression of parthenogenetically activated mouse oocytes to interphase is dependent on the level of internal calcium. J. Cell Sci. 1992, 103, 389–396. [Google Scholar]
- Zernicka-Goetz, M. Spontaneous and induced activation of rat oocytes. Mol. Reprod. Dev. 1991, 28, 169–176. [Google Scholar]
- Kubiak, J.Z.; Weber, M.; Geraud, G.; Maro, B. Cell cycle modification during the transitions between meiotic M-phases in mouse oocytes. J. Cell Sci. 1992, 102, 457–467. [Google Scholar]
- Du Pasquier, D.; Dupre, A.; Jessus, C. Unfertilized Xenopus eggs die by bad-dependent apoptosis under the control of Cdk1 and JNK. PLoS One 2011, 6, e23672. [Google Scholar]
- Iguchi, S.; Iwasaki, T.; Fukami, Y.; Tokmakov, A.A. Unlaid Xenopus eggs degrade by apoptosis in the genital tract. BMC Cell Biol. 2013, 14. [Google Scholar] [CrossRef]
- Tokmakov, A.A.; Iguchi, S.; Iwasaki, T.; Fukami, Y. Unfertilized frog eggs die by apoptosis following meiotic exit. BMC Cell Biol. 2011, 12. [Google Scholar] [CrossRef]
- Bayaa, M.; Booth, R.A.; Sheng, Y.; Liu, X.J. The classical progesterone receptor mediates Xenopus oocyte maturation through a nongenomic mechanism. Proc. Natl. Acad. Sci. USA 2000, 97, 12607–12612. [Google Scholar]
- Tian, J.; Kim, S.; Heilig, E.; Ruderman, J.V. Identification of XPR-1, a progesterone receptor required for Xenopus oocyte activation. Proc. Natl. Acad. Sci. USA 2000, 97, 14358–14363. [Google Scholar]
- Josefsberg Ben-Yehoshua, L.; Lewellyn, A.L.; Thomas, P.; Maller, J.L. The role of Xenopus membrane progesterone receptor b in mediating the effect of progesterone on oocyte maturation. Mol. Endocrinol. 2007, 21, 664–673. [Google Scholar]
- Maller, J.L. The elusive progesterone receptor in Xenopus oocytes. Proc. Natl. Acad. Sci. USA 2001, 98, 8–10. [Google Scholar]
- Martinez, S.; Pastén, P.; Suarez, K.; García, A.; Nualart, F.; Montecino, M.; Hinrichs, M.V.; Olate, J. Classical Xenopus laevis progesterone receptor associates to the plasma membrane through its ligand-binding domain. J. Cell Physiol. 2007, 211, 560–567. [Google Scholar]
- Martinez, S.; Grandy, R.; Pasten, P.; Montecinos, H.; Montecino, M.; Olate, J.; Hinrichs, M.V. Plasma membrane destination of the classical Xenopus laevis progesterone receptor accelerates progesterone-induced oocyte maturation. J. Cell Biochem. 2006, 99, 853–859. [Google Scholar]
- Lutz, L.B.; Cole, L.M.; Gupta, M.K.; Kwist, K.W.; Auchus, R.J.; Hammes, S.R. Evidence that androgens are the primary steroids produced by Xenopus laevis ovaries and may signal through the classical androgen receptor to promote oocyte maturation. Proc. Natl. Acad. Sci. USA 2001, 98, 13728–13733. [Google Scholar]
- Yang, W.H.; Lutz, L.B.; Hammes, S.R. Xenopus laevis ovarian CYP17 is a highly potent enzyme expressed exclusively in oocytes. Evidence that oocytes play a critical role in Xenopus ovarian androgen production. J. Biol. Chem. 2003, 278, 9552–9559. [Google Scholar]
- Wessel, G.M.; Brooks, J.M.; Green, E.; Haley, S.; Voronina, E.; Wong, J.; Zaydfudim, V.; Conner, S. The biology of cortical granules. Int. Rev. Cytol. 2001, 209, 117–206. [Google Scholar]
- Wessel, G.M.; Wong, J.L. Cell surface changes in the egg at fertilization. Mol. Reprod. Dev. 2009, 76, 942–953. [Google Scholar]
- Hedrick, J.L. Anuran and pig egg zona pellucida glycoproteins in fertilization and early development. Int. J. Dev. Biol. 2008, 52, 683–701. [Google Scholar]
- Goudet, G.; Mugnier, S.; Callebaut, I.; Monget, P. Phylogenetic analysis and identification of pseudogenes reveal a progressive loss of zona pellucida genes during evolution of vertebrates. Biol. Reprod. 2008, 78, 796–806. [Google Scholar]
- Gerton, G.L. Biochemical studies of the envelope transformations in Xenopus laevis eggs. Adv. Exp. Med. Biol. 1986, 207, 133–149. [Google Scholar]
- Yoshizaki, N. Comparison of two lectins isolated from Xenopus cortical granules. Zool. Sci. 1989, 6, 507–514. [Google Scholar]
- Ueda, Y.; Kubo, H.; Iwao, Y. Characterization of the acrosome reaction-inducing substance in Xenopus (ARISX) secreted form the oviductal pars recta onto the vitelline envelope. Dev. Biol. 2003, 264, 289–298. [Google Scholar]
- Guerardel, Y.; Kol, O.; Maes, E.; Lefebvre, T.; Boilly, B.; Davril, M.; Strecker, G. O-glycan variability of egg-jelly mucins from Xenopus laevis: Characterization of four phenotypes that differ by the terminal glycosylation of their mucins. Biochem. J. 2000, 352, 449–463. [Google Scholar]
- Katagiri, C. Chemical analysis of toad egg-jelly in relation to its “sperm-capacitating” activity. Dev. Growth Differ. 1973, 15, 81–92. [Google Scholar]
- Al-Anzi, B.; Chandler, D.E. A sperm chemoattractant is released from Xenopus egg jelly during spawning. Dev. Biol. 1998, 198, 366–375. [Google Scholar]
- Olson, J.H.; Xiang, X.; Ziegert, T.; Kittelson, A.; Rawis, A.; Bieber, A.L.; Chandler, D.E. Allurin, a 21-kDa sperm chemoattractant from Xenopus egg jelly, is related to mammalian sperm-binding proteins. Proc. Natl. Acad. Sci. USA 2001, 98, 11205–11210. [Google Scholar]
- Nagai, K.; Ishida, T.; Hashimoto, T.; Harada, Y.; Ueno, S.; Ueda, Y.; Kubo, H.; Iwao, Y. The sperm-surface glycoprotein, SGP, is necessary for fertilization in the frog, Xenopus laevis. Dev. Growth Differ. 2009, 51, 499–510. [Google Scholar]
- Kubo, H.; Shiga, K.; Harada, Y.; Iwao, Y. Analysis of a sperm surface molecule that binds to a vitelline envelope component of Xenopus laevis eggs. Mol. Reprod. Dev. 2010, 77, 728–735. [Google Scholar]
- Tian, J.M.; Gong, H.; Thomsen, G.H.; Lennarz, W.J. Gamete interactions in Xenopus laevis: Identification of sperm binding glycoproteins in the egg vitelline envelope. J. Cell Biol. 1997, 136, 1099–1108. [Google Scholar]
- Tian, J.M.; Gong, H.; Thomsen, G.H.; Lennarz, W.J. Xenopus laevis sperm-egg adhesion is regulated by modifications in the sperm receptor and the egg vitelline envelope. Dev. Biol. 1997, 187, 143–153. [Google Scholar]
- Vo, L.H.; Yen, T.Y.; Macher, B.A.; Hedrick, J.L. Identification of the ZPC oligosaccharide ligand involved in sperm binding and the glycan structures of Xenopus laevis vitelline envelope glycoproteins. Biol. Reprod. 2003, 69, 1822–1830. [Google Scholar]
- Iwao, Y.; Shiga, K.; Shiroshita, A.; Yoshikawa, T.; Sakiie, M.; Ueno, T.; Ueno, S.; Ijiri, T.W.; Sato, K. The need of MMP-2 on the sperm surface for Xenopus fertilization: Its role on a fast electrical block to polyspermy. Mech. Dev. 2014, 134, 80–95. [Google Scholar]
- Iwao, Y. Fertilization in amphibians. In Fertilization in Protozoa and Metazoan Animal; Tarin, J.J., Cano, A., Eds.; Springer-Verlag: Berlin, Germany, 2000; pp. 147–191. [Google Scholar]
- Iwao, Y. Egg activation and physiological polyspermy. Reproduction 2012, 144, 11–22. [Google Scholar]
- Iwao, Y.; Fujimura, T. Activation of Xenopus eggs by RGD-containing peptides accompanied by intracellular Ca2+ release. Dev. Biol. 1996, 209, 308–320. [Google Scholar]
- Sato, K.; Iwao, Y.; Fujimura, T.; Tamaki, I.; Ogawa, K.; Iwasaki, T.; Tokmakov, A.A.; Hatano, O.; Fukami, Y. Evidence for the involvement of a Src-related tyrosine kinase in Xenopus egg activation. Dev. Biol. 1999, 209, 308–320. [Google Scholar]
- Shilling, F.M.; Krätzschmar, J.; Cai, H.; Weskamp, G.; Gayko, U.; Leibow, J.; Myles, D.G.; Nuccitelli, R.; Blobel, C.P. Identification of metalloproteinase/disintegrins in Xenopus laevis testis with a potential role in fertilization. Dev. Biol. 1997, 186, 155–164. [Google Scholar]
- Shilling, F.M.; Magie, C.R.; Nuccitelli, R. Voltage-dependent activation of frog eggs by a sperm surface disintegrin peptide. Dev. Biol. 1998, 202, 113–124. [Google Scholar]
- Sato, K.; Aoto, M.; Mori, K.; Akasofu, S.; Tokmakov, A.A.; Sahara, S.; Fukami, Y. Purification and characterization of a Src-related p57 protein-tyrosine kinase from Xenopus oocytes. Isolation of an inactive form of the enzyme and its activation and translocation upon fertilization. J. Biol. Chem. 1996, 271, 13250–13257. [Google Scholar]
- Sato, K.; Tokmakov, A.A.; Iwasaki, T.; Fukami, Y. Tyrosine kinase-dependent activation of phospholipase Cg is required for calcium transient in Xenopus egg fertilization. Dev. Biol. 2000, 224, 453–469. [Google Scholar]
- Sato, K.; Tokmakov, A.A.; Fukami, Y. Fertilization signalling and protein-tyrosine kinases. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2000, 126, 129–148. [Google Scholar]
- Sato, K.; Ogawa, K.; Tokmakov, A.A.; Iwasaki, T.; Fukami, Y. Hydrogen peroxide induces Src family tyrosine kinase-dependent activation of Xenopus eggs. Dev. Growth Differ. 2001, 43, 55–72. [Google Scholar]
- Sato, K.; Tokmakov, A.A.; He, C.L.; Kurokawa, M.; Iwasaki, T.; Shirouzu, M.; Fissore, R.A.; Yokoyama, S.; Fukami, Y. Reconstitution of Src-dependent phospholipase Cgamma phosphorylation and transient calcium release by using membrane rafts and cell-free extracts from Xenopus eggs. J. Biol. Chem. 2003, 278, 38413–38420. [Google Scholar]
- Sato, K.; Fukami, Y.; Stith, B.J. Signal transduction pathways leading to Ca2+ release in a vertebrate model system: Lessons from Xenopus eggs. Semin. Cell Dev. Biol. 2006, 17, 285–292. [Google Scholar]
- Iwasaki, T.; Koretomo, Y.; Fukuda, T.; Paronetto, M.P.; Sette, C.; Fukami, Y.; Sato, K. Expression, phosphorylation, and mRNA-binding of heterogenous nuclear ribonucleoprotein K in Xenopus oocytes, eggs, and early embryos. Dev. Growth Differ. 2008, 50, 23–40. [Google Scholar]
- Aoto, M.; Sato, K.; Takeba, S.; Horiuchi, Y.; Iwasaki, T.; Tokmakov, A.A.; Fukami, Y. A 58-kDa Shc protein is present in Xenopus eggs and is phosphorylated on tyrosine residues upon egg activation. Biochem. Biophys. Res. Commun. 1999, 258, 265–270. [Google Scholar]
- Mammadova, G.; Iwasaki, T.; Tokmakov, A.A.; Fukami, Y.; Sato, K. Evidence that phosphatidylinositol 3-kinase is involved in sperm-induced tyrosine kinase signaling in Xenopus egg fertilization. BMC Dev. Biol. 2009, 9. [Google Scholar] [CrossRef]
- Sato, K.; Iwasaki, T.; Hirahara, S.; Nishihira, Y.; Fukami, Y. Molecular dissection of egg fertilization signaling with the aid of tyrosine kinase-specific inhibitor and activator strategies. Biochim. Biophys. Acta 2004, 1697, 103–121. [Google Scholar]
- Abassi, Y.A.; Carroll, D.J.; Giusti, A.F.; Belton, R.J., Jr.; Foltz, K.R. Evidence that Src-type tyrosine kinase activity is necessary for initiation of calcium release at fertilization in sea urchin eggs. Dev. Biol. 2000, 218, 206–219. [Google Scholar]
- Carroll, D.J.; Albay, D.T.; Terasaki, M.; Jaffe, L.A.; Foltz, K.R. Identification of PLCγ-dependent and -independent events during fertilization of sea urchin eggs. Dev. Biol. 1999, 206, 232–247. [Google Scholar]
- Giusti, A.F.; Carroll, D.J.; Abassi, Y.A.; Foltz, K.R. Evidence that a starfish egg Src family tyrosine kinase associates with PLC-g1 SH2 domains at fertilization. Dev. Biol. 1999, 208, 189–199. [Google Scholar]
- Giusti, A.F.; Carroll, D.J.; Abassi, Y.A.; Terasaki, M.; Foltz, K.R.; Jaffe, L.A. Requirement of a Src family kinase for initiating calcium release at fertilization in starfish eggs. J. Biol. Chem. 1999, 274, 29318–29322. [Google Scholar]
- Kinsey, W.H.; Shen, S.S. Role of the Fyn kinase in calcium release during fertilization of the sea urchin egg. Dev. Biol. 2000, 225, 253–264. [Google Scholar]
- Talmor, A.; Kinsey, W.H.; Shalgi, R. Expression and immunolocalization of p59c−fyn tyrosine kinase in rat eggs. Dev. Biol. 1998, 194, 38–46. [Google Scholar]
- Runft, L.L.; Jaffe, L.A. Sperm extract injection into ascidian eggs signals Ca2+ release by the same pathway as fertilization. Development 2000, 127, 3227–3236. [Google Scholar]
- Rice, A.; Parrington, J.; Jones, K.T.; Swann, K. Mammalian sperm contain a Ca2+-sensitive phospholipase C activity that can generate InsP3 from PIP2 associated with intracellular organelles. Dev. Biol. 2000, 228, 125–135. [Google Scholar]
- Kurokawa, M.; Sato, K.; Smyth, J.; Wu, H.; Fukami, K.; Takenawa, T.; Fissore, R.A. Evidence that activation of Src family kinase is not required for fertilization-associated [Ca2+]i oscillations in mouse eggs. Reproduction 2004, 127, 441–454. [Google Scholar]
- Mori, T.; Wu, G.M.; Mori, E. Expression of CD4-like structure on murine egg vitelline membrane and its signal transductive roles through p56lck in fertilization. Am. J. Reprod. Immunol. 1991, 26, 97–103. [Google Scholar]
- Rogers, N.T.; Hobson, E.; Pickering, S.; Lai, F.A.; Braude, P.; Swann, K. Phospholipase Cz causes Ca2+ oscillations and parthenogenetic activation of human oocytes. Reproduction 2004, 128, 697–702. [Google Scholar]
- Escoffier, J.; Yassine, S.; Lee, H.C.; Martinez, G.; Delaroche, J.; Coutton, C.; Karaouzène, T.; Zouari, R.; Metzler-Guillemain, C.; Pernet-Gallay, K.; et al. Subcellular localization of phospholipase Cζ in human sperm and its absence in DPY19L2-deficient sperm are consistent with its role in oocyte activation. Mol. Hum. Reprod. 2014. [Google Scholar] [CrossRef]
- Malcuit, C.; Knott, J.G.; He, C.; Wainwright, T.; Parys, J.B.; Robl, J.M.; Fissore, R.A. Fertilization and inositol 1,4,5-trisphosphate (IP3)-induced calcium release in type-1 inositol 1,4,5-trisphosphate receptor down-regulated bovine eggs. Biol. Reprod. 2005, 73, 2–13. [Google Scholar]
- Saunders, C.M.; Larman, M.G.; Parrington, J.; Cox, L.J.; Royse, J.; Blayney, L.M.; Swann, K.; Lai, F.A. PLCz: A sperm-specific trigger of Ca2+ oscillations in eggs and embryo development. Development 2002, 129, 3533–3544. [Google Scholar]
- Sette, C.; Bevilacqua, A.; Bianchini, A.; Mangia, F.; Geremia, R.; Rossi, P. Parthenogenetic activation of mouse eggs by microinjection of a truncated c-kit tyrosine kinase present in spermatozoa. Development 1997, 124, 2267–2274. [Google Scholar]
- Aarabi, M.; Balakier, H.; Bashar, S.; Moskovtsev, S.I.; Sutovsky, P.; Librach, C.L.; Oko, R. Sperm-derived WW domain-binding protein, PAWP, elicits calcium oscillations and oocyte activation in humans and mice. FASEB J. 2014, 28, 4434–4440. [Google Scholar]
- Harada, Y.; Matsumoto, T.; Hirahara, S.; Nakashima, A.; Ueno, S.; Oda, S.; Miyazaki, S.; Iwao, Y. Characterization of a sperm factor for egg activation at fertilization of the newt Cynops pyrrhogaster. Dev. Biol. 2007, 306, 797–808. [Google Scholar]
- Harada, Y.; Kawazoe, M.; Eto, Y.; Ueno, S.; Iwao, Y. The Ca2+ increase by the sperm factor in physiologically polyspermic newt fertilization: Its signaling mechanism in egg cytoplasm and the species-specificity. Dev. Biol. 2011, 351, 266–276. [Google Scholar]
- Mizushima, S.; Hiyama, G.; Shiba, K.; Inaba, K.; Dohra, H.; Ono, T.; Shimada, K.; Sasanami, T. The birth of quail chicks after intracytoplasmic sperm injection. Development 2014, 141, 3799–3806. [Google Scholar]
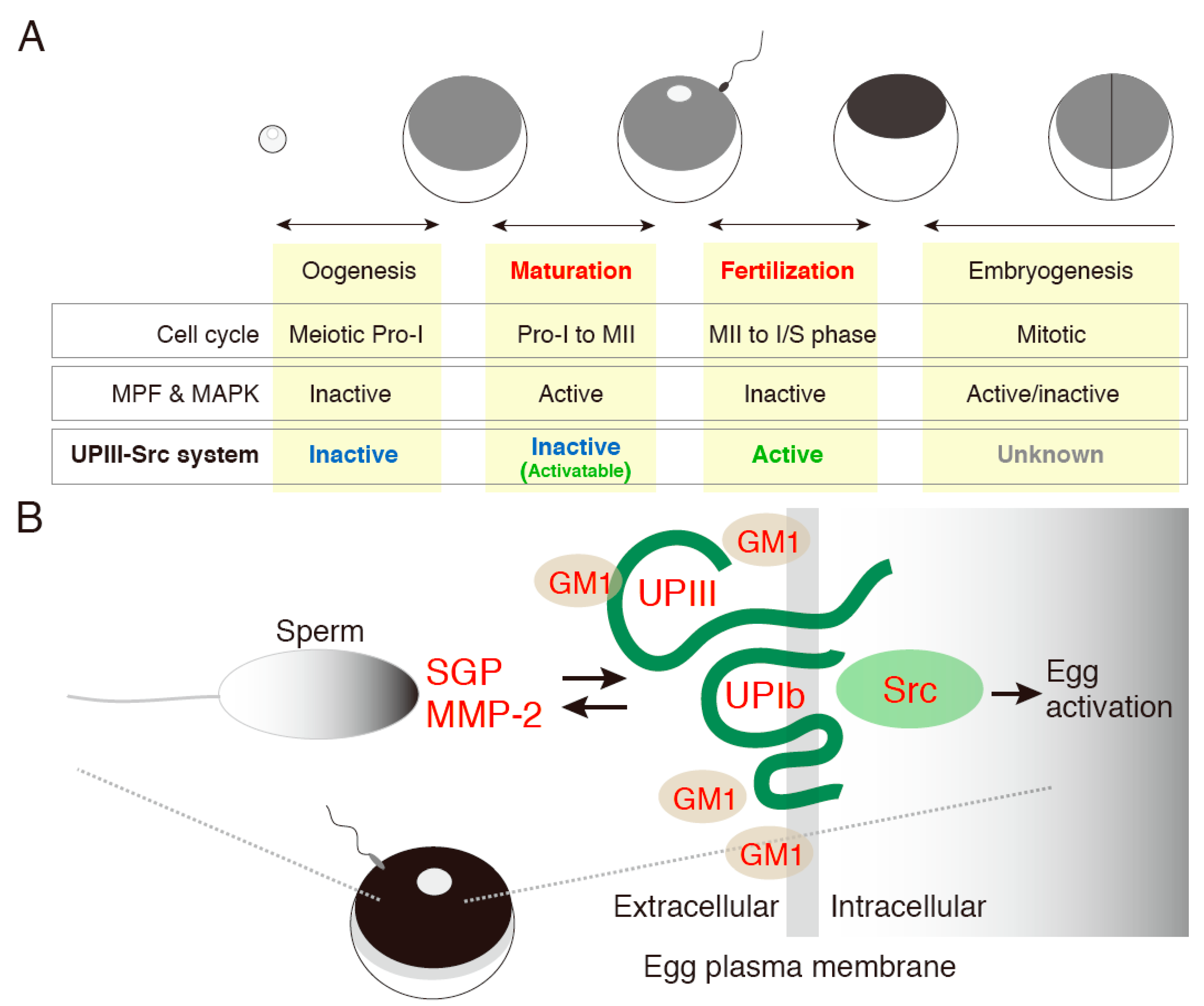
- Sakakibara, K.; Sato, K.; Yoshino, K.; Oshiro, N.; Hirahara, S.; Mahbub Hasan, A.K.; Iwasaki, T.; Ueda, Y.; Iwao, Y.; Yonezawa, K.; et al. Molecular identification and characterization of Xenopus egg uroplakin III, an egg raft-associated transmembrane protein that is tyrosine-phosphorylated upon fertilization. J. Biol. Chem. 2005, 280, 15029–15037. [Google Scholar]
- Mahbub Hasan, A.K.; Fukami, Y.; Sato, K.I. Gamete membrane microdomains and their associated molecules in fertilization signaling. Mol. Reprod. Dev. 2011, 78, 814–830. [Google Scholar]
- Sato, K.; Iwasaki, T.; Ogawa, K.; Konishi, M.; Tokmakov, A.A.; Fukami, Y. Low density detergent-insoluble membrane of Xenopus eggs: Subcellular microdomain for tyrosine kinase signaling in fertilization. Development 2002, 129, 885–896. [Google Scholar]
- Sato, K.; Yoshino, K.; Tokmakov, A.A.; Iwasaki, T.; Yonezawa, K.; Fukami, Y. Studying fertilization in cell-free extracts: Focusing on membrane/lipid raft functions and proteomics. Methods Mol. Biol. 2006, 322, 395–411. [Google Scholar]
- Mahbub Hasan, A.K.; Sato, K.; Sakakibara, K.; Ou, Z.; Iwasaki, T.; Ueda, Y.; Fukami, Y. Uroplakin III, a novel Src substrate in Xenopus egg rafts, is a target for sperm protease essential for fertilization. Dev. Biol. 2005, 286, 483–492. [Google Scholar]
- Mizote, A.; Okamoto, S.; Iwao, Y. Activation of Xenopus eggs by proteases: Possible involvement of a sperm protease in fertilization. Dev. Biol. 1999, 208, 79–92. [Google Scholar]
- Mahbub Hasan, A.K.; Ou, Z.; Sakakibara, K.; Hirahara, S.; Iwasaki, T.; Sato, K.; Fukami, Y. Characterization of Xenopus egg membrane microdomains containing uroplakin Ib/III complex: Roles of their molecular interactions for subcellular localization and signal transduction. Genes Cells 2007, 12, 251–267. [Google Scholar]
- Tu, L.; Sun, T.T.; Kreibich, G. Specific heterodimer formation is a prerequisite for uroplakins to exit from the endoplasmic reticulum. Mol. Biol. Cell 2002, 13, 4221–4230. [Google Scholar]
- Sabri, A.; Guo, J.; Elouardighi, H.; Darrow, A.L.; Andrade-Gordon, P.; Steinberg, S.F. Mechanisms of protease-activated receptor-4 actions in cardiomyocytes. Role of Src tyrosine kinase. J. Biol. Chem. 2003, 278, 11714–11720. [Google Scholar]
- Vu, T.K.; Hung, D.T.; Wheaton, V.I.; Coughlin, S.R. Molecular cloning of a functional thrombin receptor reveals a novel proteolytic mechanism of receptor activation. Cell 1991, 64, 1057–1068. [Google Scholar]
- Fortini, M.E. γ-secretase-mediated proteolysis in cell-surface-receptor signalling. Nat. Rev. Mol. Cell Biol. 2002, 3, 673–684. [Google Scholar]
- Nagano, O.; Saya, H. Mechanism and biological significance of CD44 cleavage. Cancer Sci. 2004, 95, 930–935. [Google Scholar]
- Bates, R.C.; Fees, C.P.; Holland, W.L.; Winger, C.C.; Batbayer, K.; Ancar, R.; Bergren, T.; Petcoff, D.; Stith, B.J. Activation of Src and release of intracellular calcium by phosphatidic acid during Xenopus laevis fertilization. Dev. Biol. 2014, 386, 165–180. [Google Scholar]
- Mahbub Hasan, A.K.; Hashimoto, A.; Maekawa, Y.; Matsumoto, T.; Kushima, S.; Ijiri, T.W.; Fukami, Y.; Sato, K. The egg membrane microdomain-associated uroplakin III-Src system becomes functional during oocyte maturation and is required for bidirectional gamete signaling at fertilization in Xenopus laevis. Development 2014, 141, 1705–1714. [Google Scholar]
- Anifandis, G.; Messini, C.; Dafopoulos, K.; Sotiriou, S.; Messinis, I. Molecular and cellular mechanisms of sperm-oocyte interactions opinions relative to in vitro fertilization (IVF). Int. J. Mol. Sci. 2014, 15, 12972–12997. [Google Scholar]
- Okabe, M. The cell biology of mammalian fertilization. Development 2013, 140, 4471–4479. [Google Scholar]
- Kaji, K.; Oda, S.; Shikano, T.; Ohnuki, T.; Uematsu, Y.; Sakagami, J.; Tada, N.; Miyazaki, S.; Kudo, A.; et al. The gamete fusion process is defective in eggs of CD9-deficient mice. Nat. Genet. 2000, 24, 279–282. [Google Scholar]
- Le Naour, F.; Rubinstein, E.; Jasmin, C.; Prenant, M.; Boucheix, C. Severely reduced female fertility in CD9-deficient mice. Science 2000, 287, 319–321. [Google Scholar]
- Miyado, K.; Yamada, G.; Yamada, S.; Hasuwa, H.; Nakamura, Y.; Ryu, F.; Suzuki, K.; Kosai, K.; Inoue, K.; Ogura, A.; et al. Requirement of CD9 on the egg plasma membrane for fertilization. Science 2000, 287, 321–324. [Google Scholar]
- Barraud-Lange, V.; Naud-Barriant, N.; Bomsel, M.; Wolf, J.P.; Ziyyat, A. Transfer of oocyte membrane fragments to fertilizing spermatozoa. FASEB J. 2007, 21, 3446–3449. [Google Scholar]
- Ziyyat, A.; Rubinstein, E.; Monier-Gavelle, F.; Barraud, V.; Kulski, O.; Prenant, M.; Boucheix, C.; Bomsel, M.; Wolf, J.P. CD9 controls the formation of clusters that contain tetraspanins and the integrin a6b1, which are involved in human and mouse gamete fusion. J. Cell Sci. 2006, 119, 416–424. [Google Scholar]
- Miyado, K.; Yoshida, K.; Yamagata, K.; Sakakibara, K.; Okabe, M.; Wang, X.; Miyamoto, K.; Akutsu, H.; Kondo, T.; Takahashi, Y.; et al. The fusing ability of sperm is bestowed by CD9-containing vesicles released from eggs in mice. Proc. Natl. Acad. Sci. USA 2008, 105, 12921–12926. [Google Scholar]
- Barraud-Lange, V.; Chalas Boissonnas, C.; Serres, C.; Auer, J.; Schmitt, A.; Lefèvre, B.; Wolf, J.P.; Ziyyat, A. Membrane transfer from oocyte to sperm occurs in two CD9-independent ways that do not supply the fertilising ability of CD9-deleted oocytes. Reproduction 2012, 144, 53–66. [Google Scholar]
- Gupta, S.; Primakoff, P.; Myles, D.G. Can the presence of wild-type oocytes during insemination rescue the fusion defect of CD9 null oocytes? Mol. Reprod. Dev. 2009, 76. [Google Scholar] [CrossRef]
- Jégou, A.; Ziyyat, A.; Barraud-Lange, V.; Perez, E.; Wolf, J.P.; Pincet, F.; Gourier, C. CD9 tetraspanin generates fusion competent sites on the egg membrane for fertilization. Proc. Natl. Acad. Sci. USA 2011, 108, 10946–10951. [Google Scholar]
- Inoue, N.; Ikawa, M.; Isotani, A.; Okabe, M. The immunoglobulin superfamily protein Izumo is required for sperm to fuse with eggs. Nature 2005, 434, 234–238. [Google Scholar]
- Inoue, N.; Hamada, D.; Kamikubo, H.; Hirata, K.; Kataoka, M.; Yamamoto, M.; Ikawa, M.; Okabe, M.; Hagihara, Y. Molecular dissection of IZUMO1, a sperm protein essential for sperm-egg fusion. Development 2013, 140, 3221–3229. [Google Scholar]
- Bianchi, E.; Doe, B.; Goulding, D.; Wright, G.J. Juno is the egg Izumo receptor and is essential for mammalian fertilization. Nature 2014, 508, 483–487. [Google Scholar]
- Sprunck, S.; Rademacher, S.; Vogler, F.; Gheyselinck, J.; Grossniklaus, U.; Dresselhaus, T. Egg cell-secreted EC1 triggers sperm cell activation during double fertilization. Science 2012, 338, 1093–1097. [Google Scholar]
- Mori, T.; Kuroiwa, H.; Higashiyama, T.; Kuroiwa, T. GENERATIVE CELL SPECIFIC 1 is essential for angiosperm fertilization. Nat. Cell Biol. 2006, 8, 64–71. [Google Scholar]
- Lei, Y.; Guo, X.; Liu, Y.; Cao, Y.; Deng, Y.; Chen, X.; Cheng, C.H.; Dawid, I.B.; Chen, Y.; Zhao, H. Efficient targeed gene disruption in Xenopus embryos using engineered transcription activator-like effector nucleases (TALEs). Proc. Natl. Acad. Sci. USA 2013, 109, 17484–17489. [Google Scholar]
- Nakajima, K.; Nakai, Y.; Okada, M.; Yaoita, Y. Targeted gene disruption in the Xenopus tropicalis using designed TALE nucleases. Zool. Sci. 2013, 30, 455–460. [Google Scholar]
- Suzuki, K.T.; Isoyama, Y.; Kashiwagi, K.; Sakuma, T.; Ochiai, H.; Sakamoto, N.; Furuno, N.; Kashiwagi, A.; Yamamoto, T. High efficiency TALEs enable F0 functional analysis by targeted gene disruption in Xenopus laevis embryos. Biol. Open 2013, 2, 448–452. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, K.-i. Transmembrane Signal Transduction in Oocyte Maturation and Fertilization: Focusing on Xenopus laevis as a Model Animal. Int. J. Mol. Sci. 2015, 16, 114-134. https://doi.org/10.3390/ijms16010114
Sato K-i. Transmembrane Signal Transduction in Oocyte Maturation and Fertilization: Focusing on Xenopus laevis as a Model Animal. International Journal of Molecular Sciences. 2015; 16(1):114-134. https://doi.org/10.3390/ijms16010114
Chicago/Turabian StyleSato, Ken-ichi. 2015. "Transmembrane Signal Transduction in Oocyte Maturation and Fertilization: Focusing on Xenopus laevis as a Model Animal" International Journal of Molecular Sciences 16, no. 1: 114-134. https://doi.org/10.3390/ijms16010114