
Hair Dyes Resorcinol and Lawsone Reduce Production of Melanin in Melanoma Cells by Tyrosinase Activity Inhibition and Decreasing Tyrosinase and Microphthalmia-Associated Transcription Factor (MITF) Expression


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
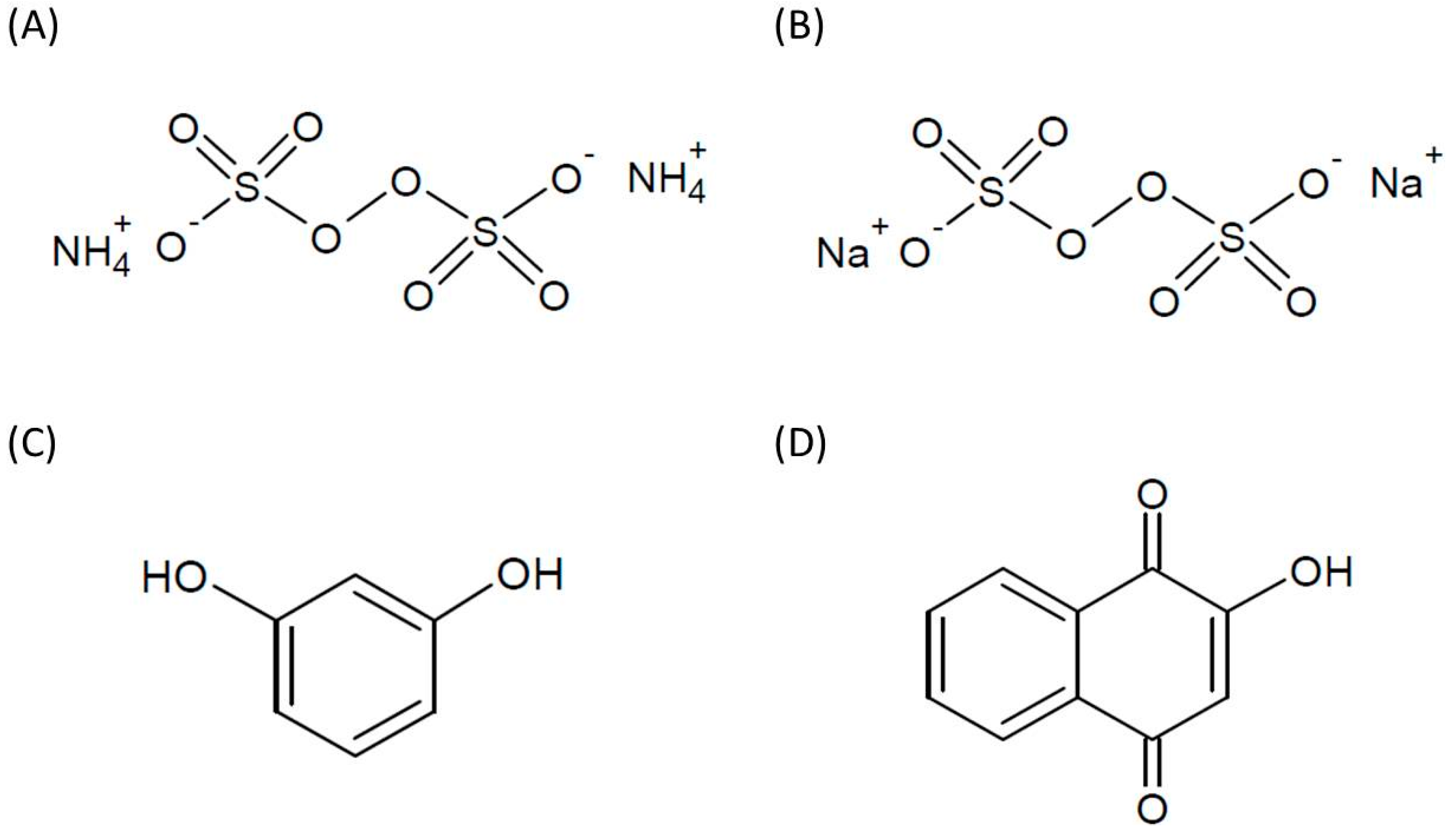
:1. Introduction

2. Results and Discussion
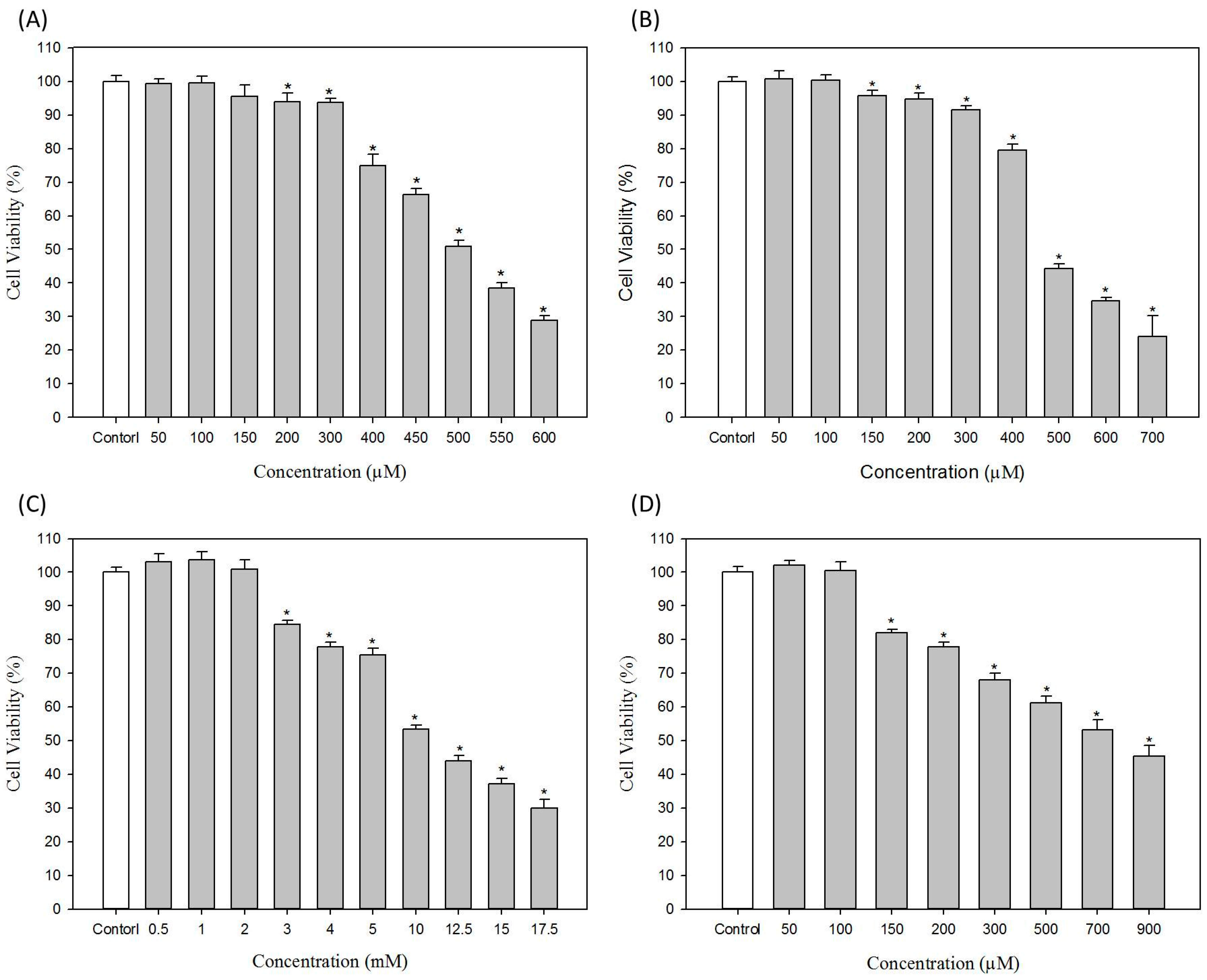
2.1. Effects of Hair Dyes on Cell Viability of B16-F10 Cells

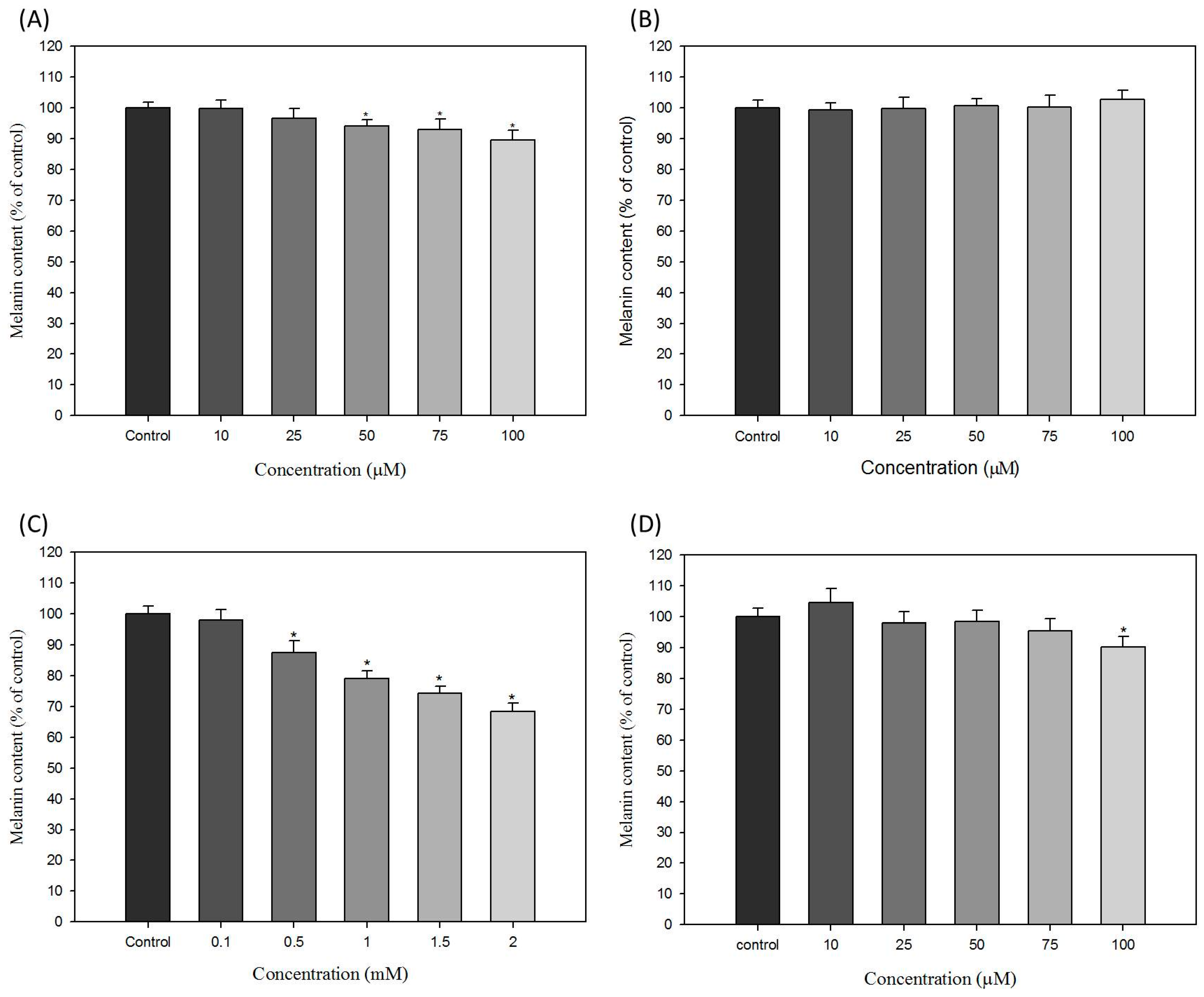
2.2. Effects of Hair Dyes on the Melanin Content of B16-F10 Cells

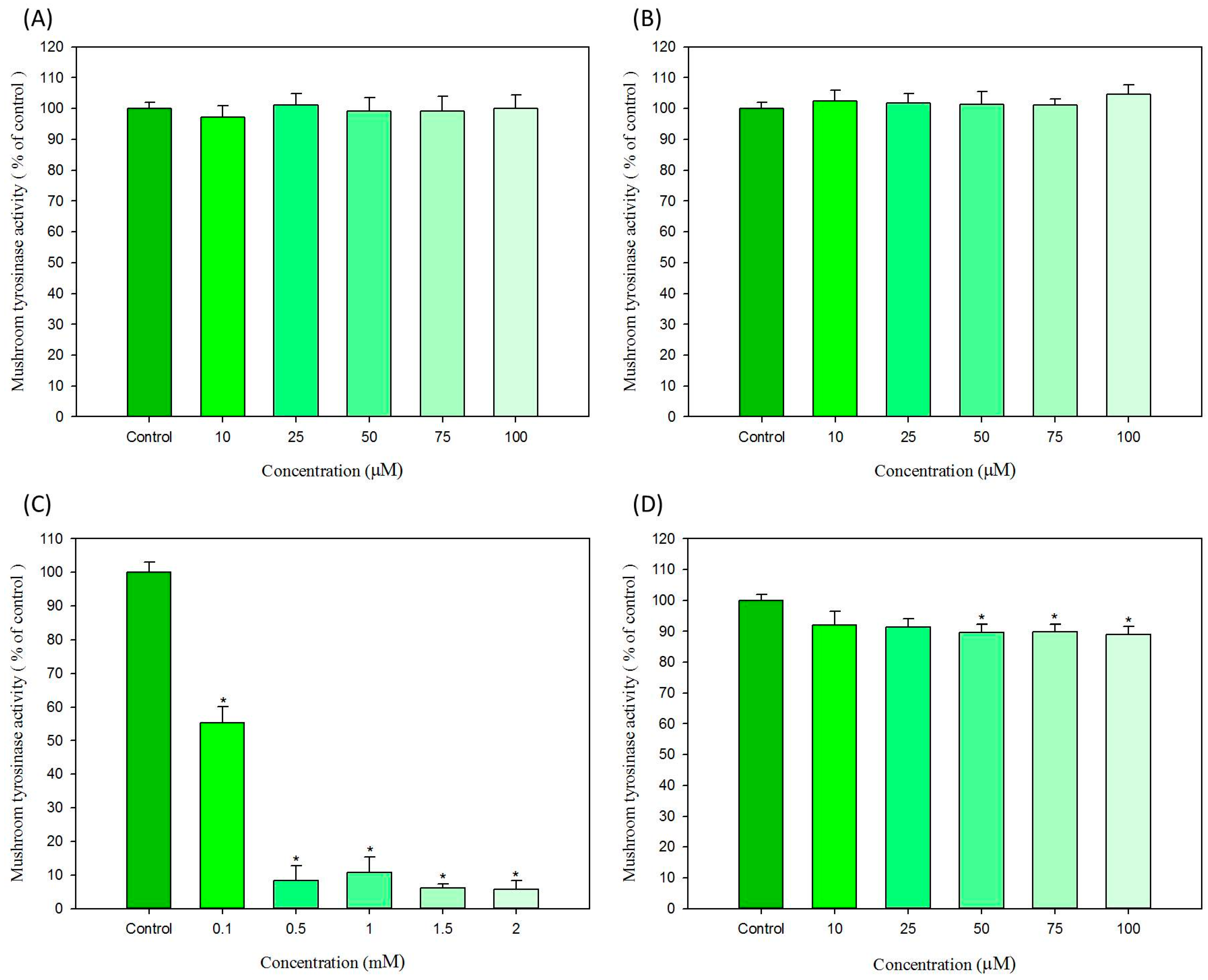
2.3. In Vitro Effects of Hair Dyes on the Activity of Mushroom Tyrosinase and B16-F10 Cellular Tyrosinase


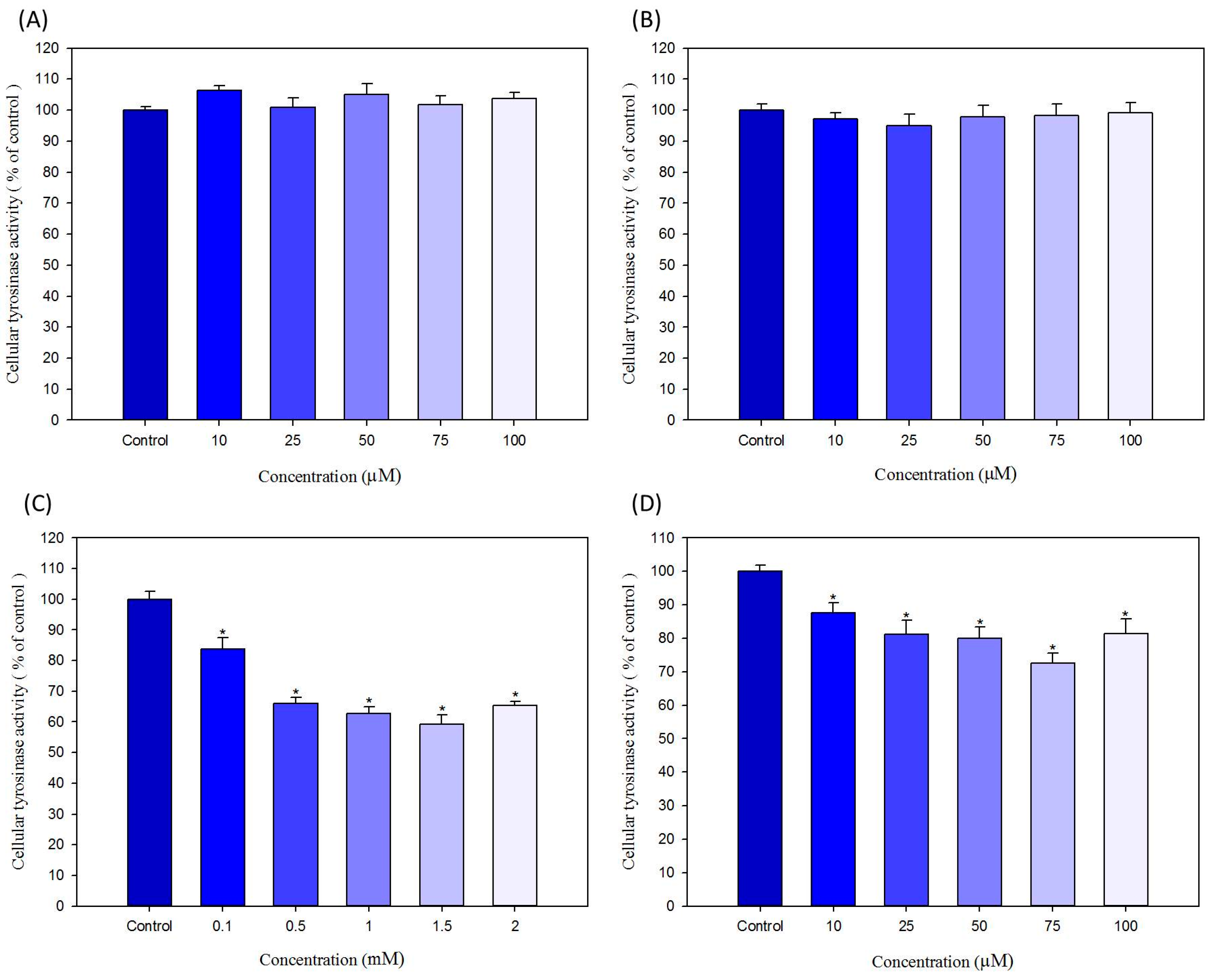
2.4. Effects of Hair Dyes on the Cellular Tyrosinase Activity in B16-F10 Cells

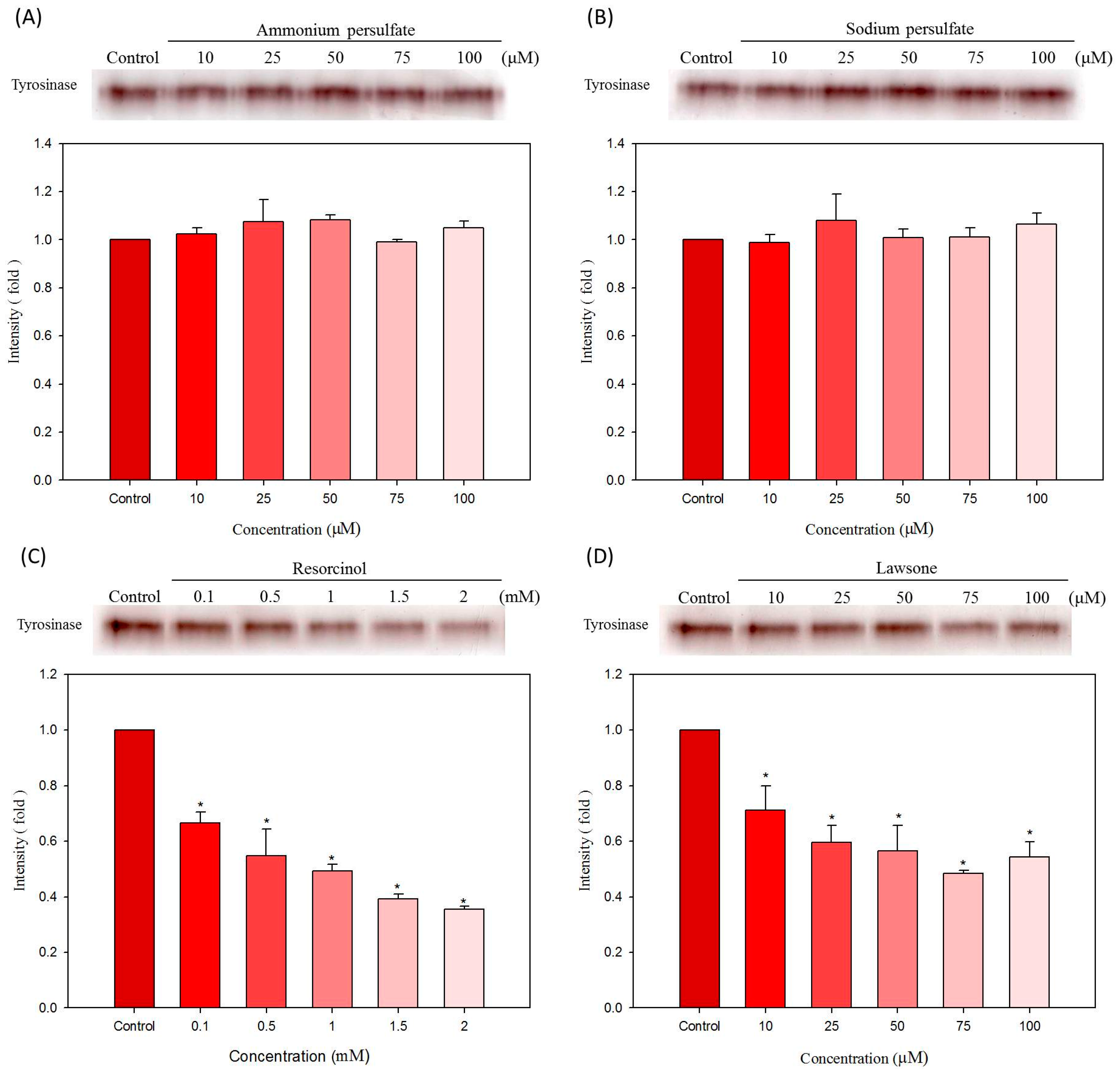
2.5. Effects of Hair Dyes on the Protein Levels of Tyrosinase and MITF in B16-F10 Cells

3. Experimental Section
3.1. Materials
3.2. Cell Line and Cell Culture
3.3. MTT Assay for Cell Viability
3.4. Melanin Content Analysis
3.5. In Vitro Tyrosinase Activity Assays
3.6. Zymographic Analysis for Cellular Tyrosinase
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bolduc, C.; Shapiro, J. Hair care products: Waving, straightening, conditioning, and coloring. Clin. Dermatol. 2001, 19, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.J.; Shin, H.; Paik, S.H.; Choi, J.W.; Lee, J.H.; Cho, S.; Kwon, O. The pattern of hair dyeing in koreans with gray hair. Ann. Dermatol. 2013, 25, 401–404. [Google Scholar] [CrossRef] [PubMed]
- AlGhamdi, K.M.; Moussa, N.A. Knowledge and practices of, and attitudes towards, the use of hair dyes among females visiting a teaching hospital in riyadh, saudi arabia. Ann. Saudi Med. 2011, 31, 613–619. [Google Scholar] [CrossRef] [PubMed]
- De Groot, A.C. Side-effects of henna and semi-permanent ‘black henna’ tattoos: A full review. Contact Dermat. 2013, 69, 1–25. [Google Scholar] [CrossRef]
- Draelos, Z.K. Hair cosmetics. Dermatol. Clin. 1991, 9, 19–27. [Google Scholar] [PubMed]
- Pang, S.; Fiume, M.Z. Final report on the safety assessment of ammonium, potassium, and sodium persulfate. Int. J. Toxicol. 2001, 20, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Hamann, D.; Yazar, K.; Hamann, C.R.; Thyssen, J.P.; Liden, C. P-phenylenediamine and other allergens in hair dye products in the united states: A consumer exposure study. Contact Dermat. 2014, 70, 213–218. [Google Scholar] [CrossRef]
- Park, H.Y.; Kosmadaki, M.; Yaar, M.; Gilchrest, B.A. Cellular mechanisms regulating human melanogenesis. Cell. Mol. Life Sci. 2009, 66, 1493–1506. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, K.U.; Ali, A.S.; Ali, S.A.; Naaz, I. Microbial tyrosinases: Promising enzymes for pharmaceutical, food bioprocessing, and environmental industry. Biochem. Res. Int. 2014, 2014. [Google Scholar] [CrossRef]
- Ito, S.; Wakamatsu, K. Human hair melanins: What we have learned and have not learned from mouse coat color pigmentation. Pigment Cell Melanoma Res. 2011, 24, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Chiarelli-Neto, O.; Ferreira, A.S.; Martins, W.K.; Pavani, C.; Severino, D.; Faiao-Flores, F.; Maria-Engler, S.S.; Aliprandini, E.; Martinez, G.R.; di Mascio, P.; et al. Melanin photosensitization and the effect of visible light on epithelial cells. PLoS One 2014, 9, e113266. [Google Scholar] [CrossRef] [PubMed]
- Videira, I.F.; Moura, D.F.; Magina, S. Mechanisms regulating melanogenesis. An. Bras. Dermatol. 2013, 88, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Cichorek, M.; Wachulska, M.; Stasiewicz, A.; Tyminska, A. Skin melanocytes: Biology and development. Postep. Dermatol. Alergol. 2013, 30, 30–41. [Google Scholar] [CrossRef]
- Seiberg, M. Age-induced hair greying—The multiple effects of oxidative stress. Int. J. Cosmet. Sci. 2013, 35, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, K.; Ogawa, M.; Sugibayashi, K.; Yamada, K.; Yamamoto, K. Relationship between tyrosinase inhibitory action and oxidation-reduction potential of cosmetic whitening ingredients and phenol derivatives. Arch. Pharmacal Res. 1999, 22, 335–339. [Google Scholar] [CrossRef]
- Khatib, S.; Nerya, O.; Musa, R.; Tamir, S.; Peter, T.; Vaya, J. Enhanced substituted resorcinol hydrophobicity augments tyrosinase inhibition potency. J. Med. Chem. 2007, 50, 2676–2681. [Google Scholar] [CrossRef] [PubMed]
- Stratford, M.R.; Ramsden, C.A.; Riley, P.A. Mechanistic studies of the inactivation of tyrosinase by resorcinol. Bioorg. Med. Chem. 2013, 21, 1166–1173. [Google Scholar] [CrossRef] [PubMed]
- Mensing, T.; Marek, W.; Raulf-Heimsoth, M.; Baur, X. Acute exposure to hair bleach causes airway hyperresponsiveness in a rabbit model. Eur. Respir. J. 1998, 12, 1371–1374. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.X.; Kubo, I. Kinetics of mushroom tyrosinase inhibition by quercetin. J. Agric. Food Chem. 2002, 50, 4108–4112. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Yang, C.H.; Chang, N.F.; Wu, P.S.; Chen, Y.S.; Lee, S.M.; Chen, C.W. Study on the stability of deoxyarbutin in an anhydrous emulsion system. Int. J. Mol. Sci. 2011, 12, 5946–5954. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Morita, M.; Ichikawa, C.; Takahashi, H.; Toriyama, M. Depigmenting mechanisms of all-trans retinoic acid and retinol on b16 melanoma cells. Biosci. Biotechnol. Biochem. 2008, 72, 2589–2597. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Lee, S.M.; Lin, C.C.; Liu, C.Y. Hispolon decreases melanin production and induces apoptosis in melanoma cells through the downregulation of tyrosinase and microphthalmia-associated transcription factor (MITF) expressions and the activation of caspase-3, -8 and -9. Int. J. Mol. Sci. 2014, 15, 1201–1215. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Sim, G.S.; Bae, J.T.; Oh, J.Y.; Lee, G.S.; Lee, D.H.; Lee, B.C.; Pyo, H.B. Synthesis and anti-melanogenic effects of lipoic acid-polyethylene glycol ester. J. Pharm. Pharmacol. 2008, 60, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.C.; Liu, K.C.; Chiou, Y.L.; Yang, C.H.; Chen, T.H.; Li, T.T.; Liu, L.L. Fenofibrate suppresses melanogenesis in b16-f10 melanoma cells via activation of the p38 mitogen-activated protein kinase pathway. Chem. Biol. Interact. 2013, 205, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.B.; Babiarz, L.; Liebel, F.; Roydon Price, E.; Kizoulis, M.; Gendimenico, G.J.; Fisher, D.E.; Seiberg, M. Modulation of microphthalmia-associated transcription factor gene expression alters skin pigmentation. J. Investig. Dermatol. 2002, 119, 1330–1340. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.A.; Cook, A.L.; Roberts, D.W.; Leonard, J.H.; Sturm, R.A. Post-transcriptional regulation of melanin biosynthetic enzymes by camp and resveratrol in human melanocytes. J. Investig. Dermatol. 2007, 127, 2216–2227. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Lee, S.M.; Lin, Y.J.; Chiang, S.H.; Lin, C.C. Effects of danshensu and salvianolic acid b from salvia miltiorrhiza bunge (lamiaceae) on cell proliferation and collagen and melanin production. Molecules 2014, 19, 2029–2041. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Chiang, S.H.; Wang, H.Y.; Wu, P.S.; Lin, C.C. Curcumin enhances the production of major structural components of elastic fibers, elastin, and fibrillin-1, in normal human fibroblast cells. Biosci. Biotechnol. Biochem. 2014. [Google Scholar] [CrossRef]
- Chen, Y.S.; Lee, S.M.; Lin, C.C.; Liu, C.Y.; Wu, M.C.; Shi, W.L. Kinetic study on the tyrosinase and melanin formation inhibitory activities of carthamus yellow isolated from carthamus tinctorius l. J. Biosci. Bioeng. 2013, 115, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Ahn, J.W.; Nam, S.H.; Yoon, C.S.; Shin, J.C.; Lee, S.H. S-(−)-10,11-dihydroxyfarnesoic acid methyl ester inhibits melanin synthesis in murine melanocyte cells. Int. J. Mol. Sci. 2014, 15, 12750–12763. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.Y.; Lai, C.C.; Lin, C.C.; Chou, S.T. Effect of vetiveria zizanioides essential oil on melanogenesis in melanoma cells: Downregulation of tyrosinase expression and suppression of oxidative stress. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-M.; Chen, Y.-S.; Lin, C.-C.; Chen, K.-H. Hair Dyes Resorcinol and Lawsone Reduce Production of Melanin in Melanoma Cells by Tyrosinase Activity Inhibition and Decreasing Tyrosinase and Microphthalmia-Associated Transcription Factor (MITF) Expression. Int. J. Mol. Sci. 2015, 16, 1495-1508. https://doi.org/10.3390/ijms16011495
Lee S-M, Chen Y-S, Lin C-C, Chen K-H. Hair Dyes Resorcinol and Lawsone Reduce Production of Melanin in Melanoma Cells by Tyrosinase Activity Inhibition and Decreasing Tyrosinase and Microphthalmia-Associated Transcription Factor (MITF) Expression. International Journal of Molecular Sciences. 2015; 16(1):1495-1508. https://doi.org/10.3390/ijms16011495
Chicago/Turabian StyleLee, Shu-Mei, Yi-Shyan Chen, Chih-Chien Lin, and Kuan-Hung Chen. 2015. "Hair Dyes Resorcinol and Lawsone Reduce Production of Melanin in Melanoma Cells by Tyrosinase Activity Inhibition and Decreasing Tyrosinase and Microphthalmia-Associated Transcription Factor (MITF) Expression" International Journal of Molecular Sciences 16, no. 1: 1495-1508. https://doi.org/10.3390/ijms16011495