
Activation of the Ubiquitin Proteasome Pathway by Silk Fibroin Modified Chitosan Nanoparticles in Hepatic Cancer Cells


Abstract
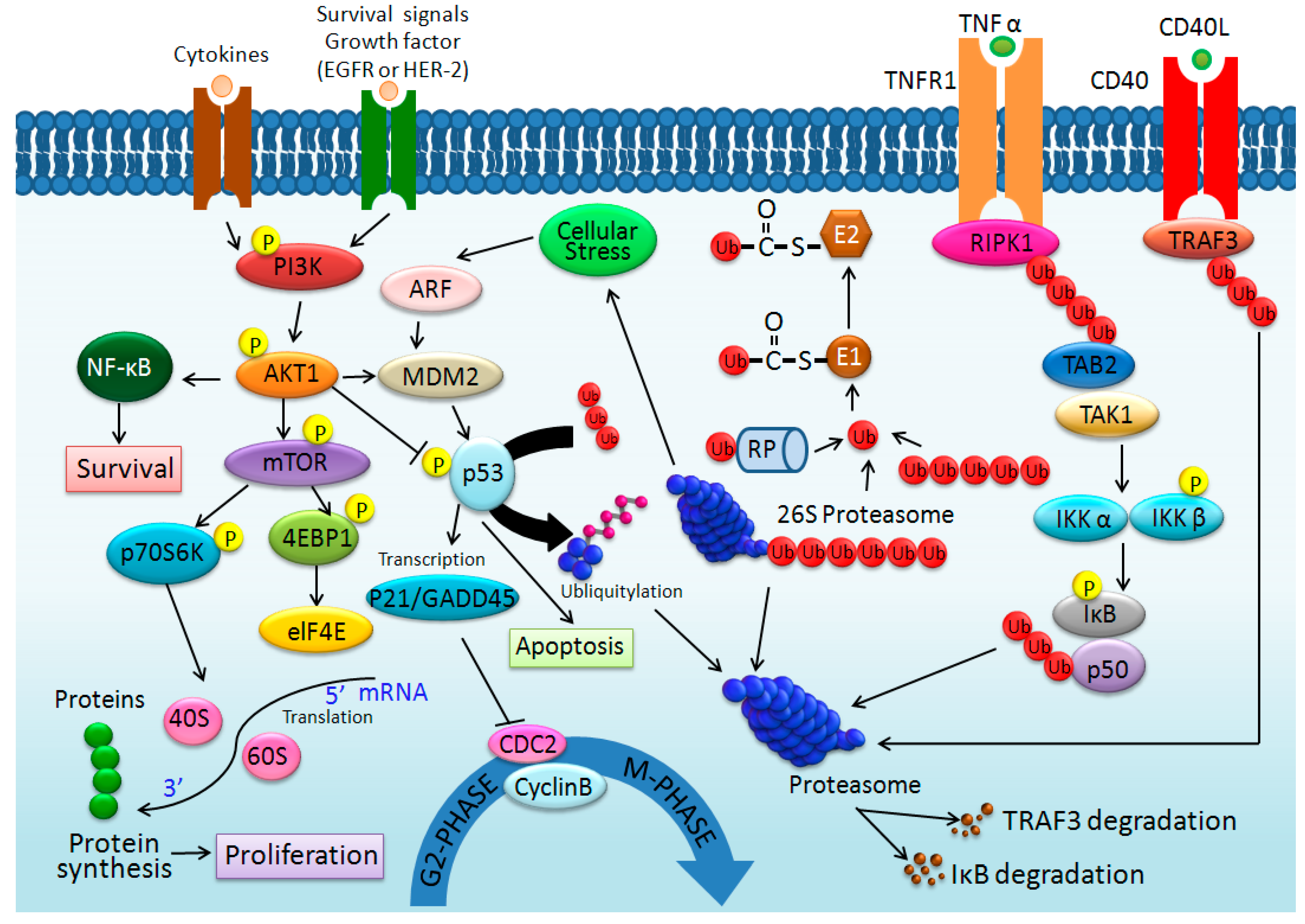
:1. Introduction
2. Results and Discussions
2.1. Size, Zeta Potential, and Morphology of Silk Fibroin Modified Chitosan Nanoparticles (SF-CSNPs)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nanoparticle | Concentration (μg/mL) | Particle Size (nm) # | Zeta Potential (mV) # |
|---|---|---|---|
| CSNP | N.D. | 239.3 ± 7.3 | 15.13 ± 0.6 |
| CSNP coating SF | 6.27 ± 0.17 ** | 311.9 ± 10.7 ** | 13.33 ± 0.3 * |
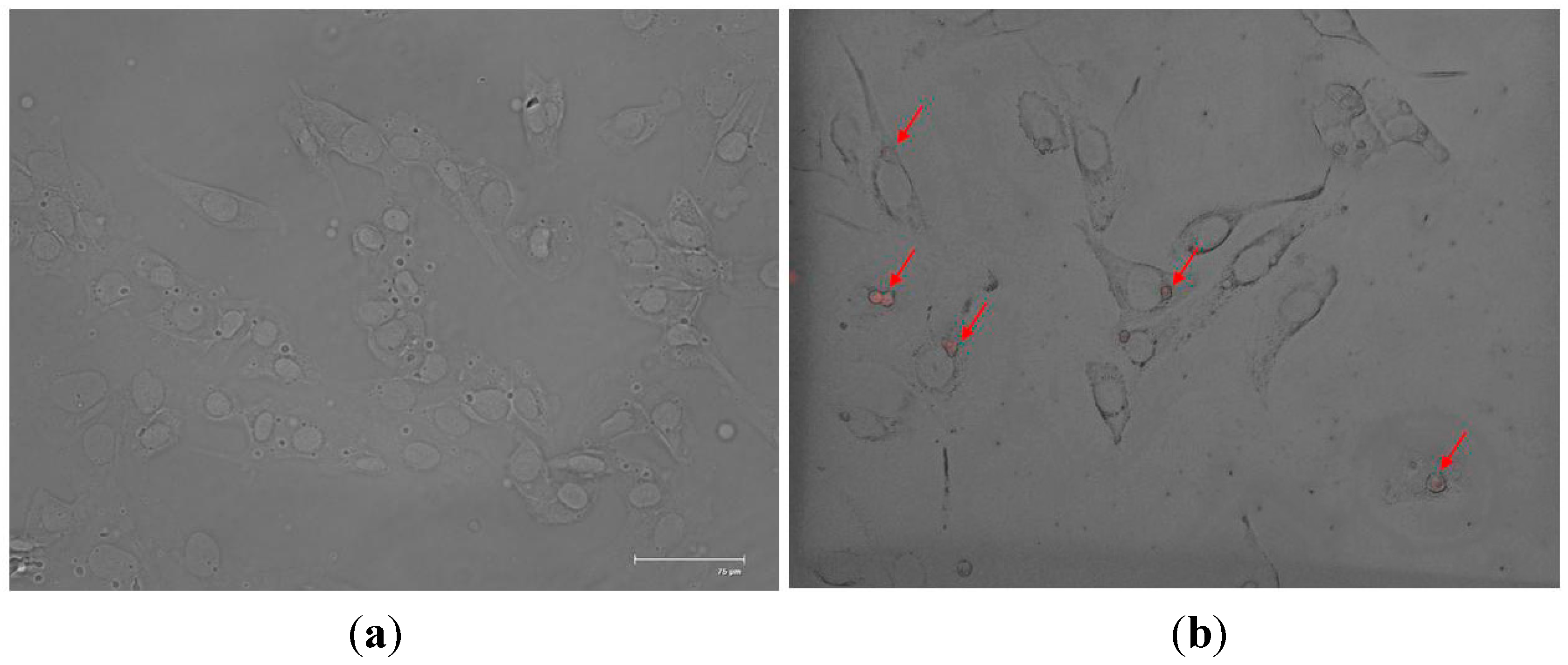
2.2. In Vitro SF-CSNP Uptake

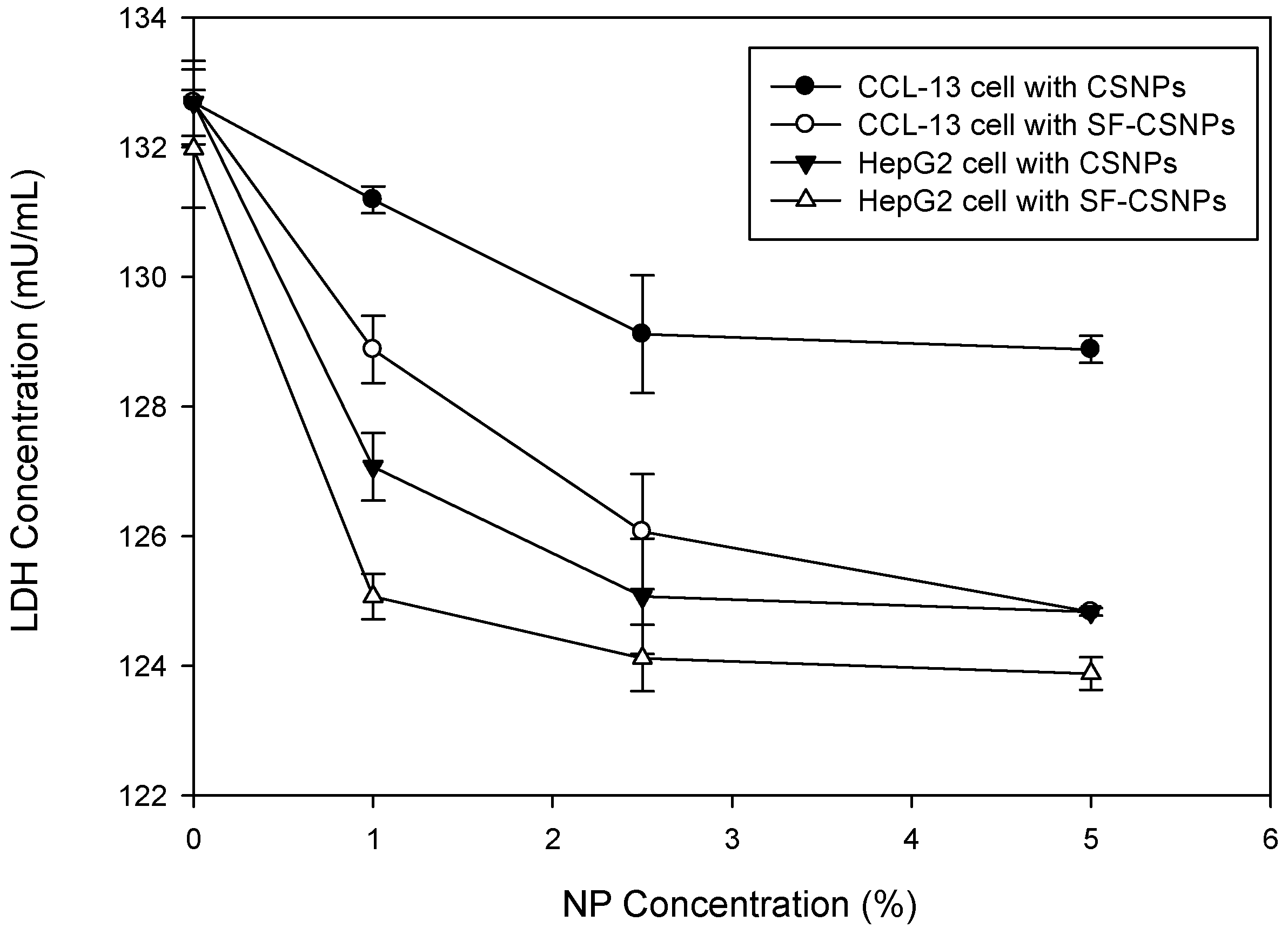
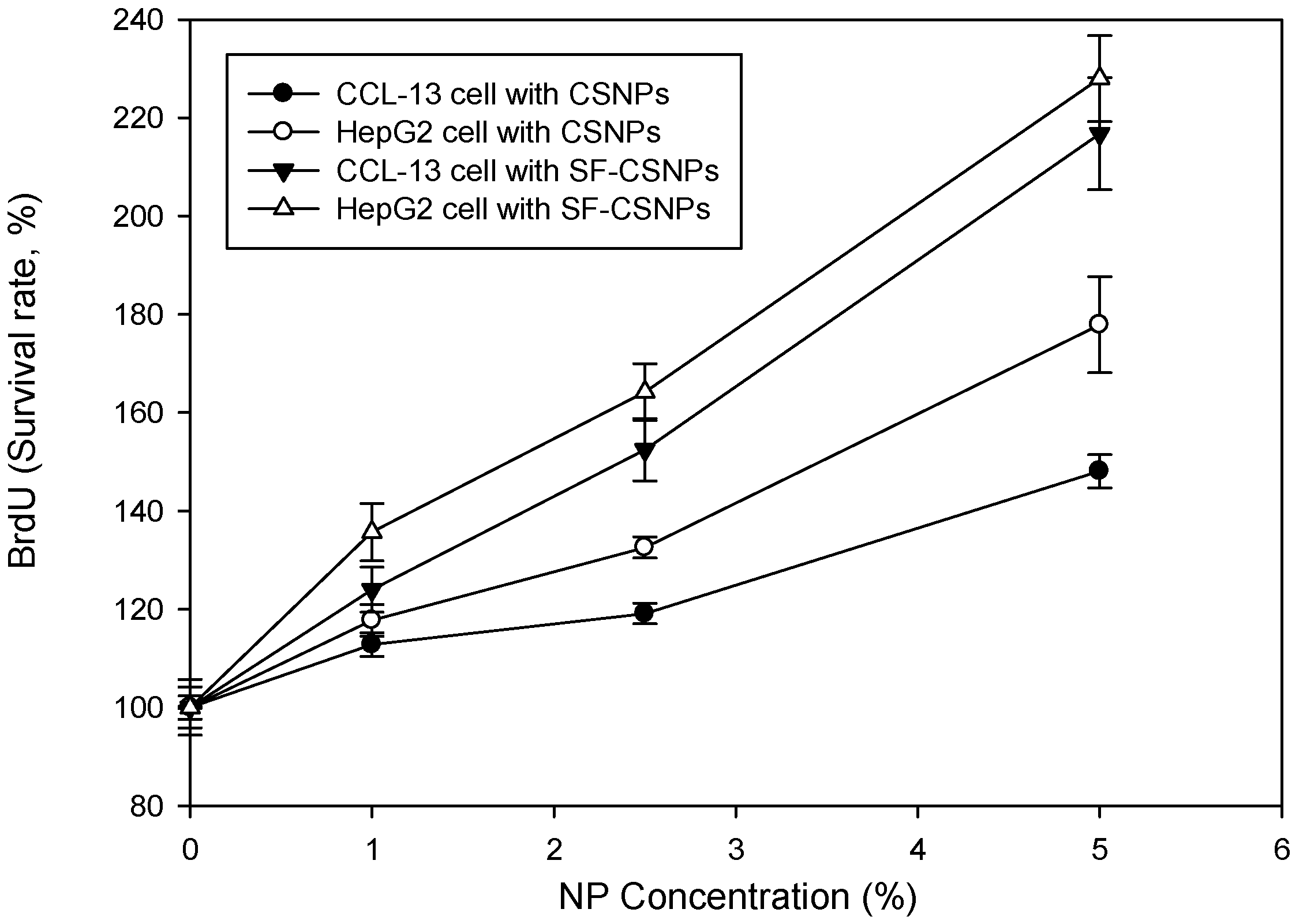
2.3. Cytotoxicity of SF-CSNP


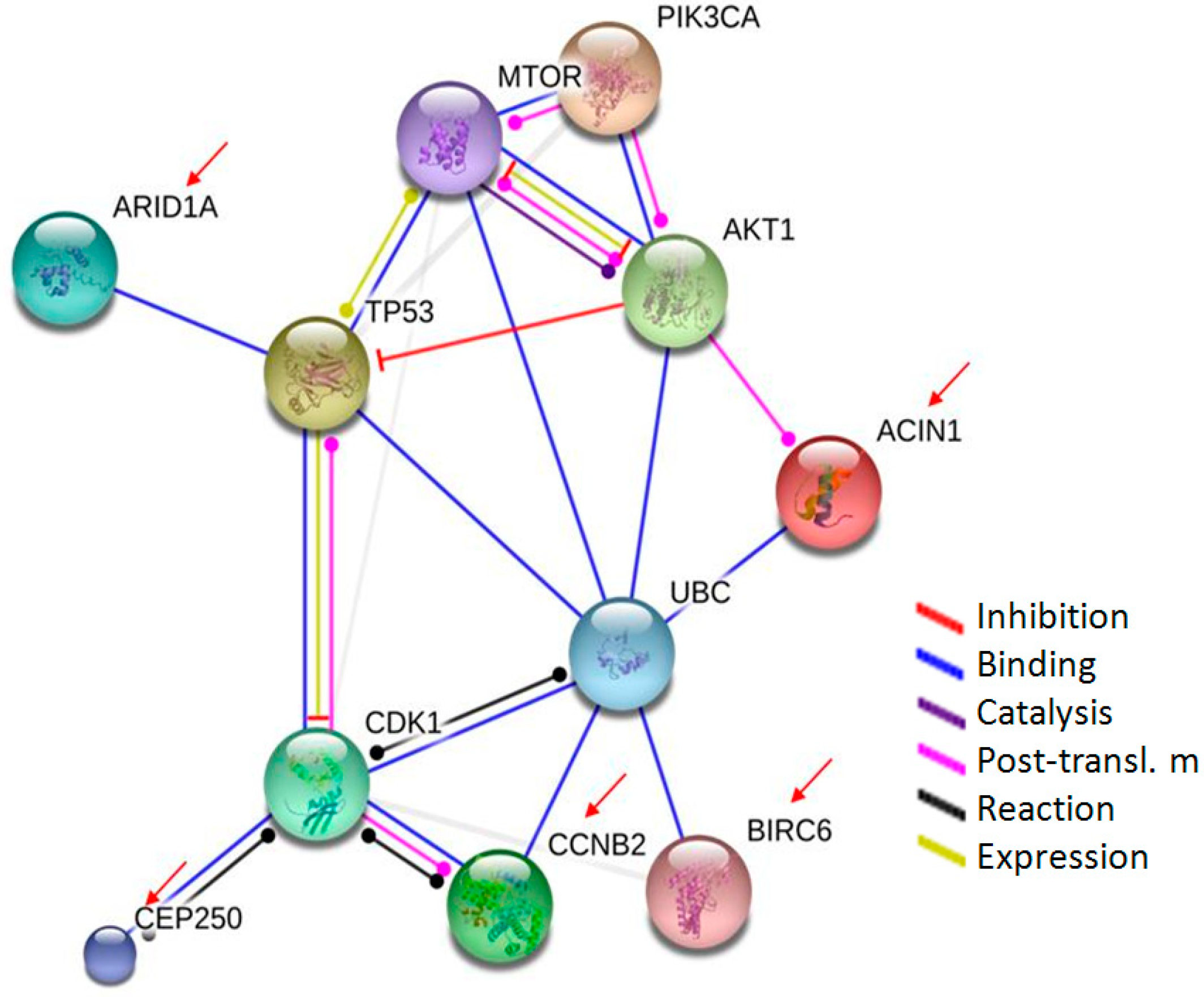
2.4. Proteomic Analysis of Cell Response to SF-CSNPs
| Short Name | Swiss-Prot No. | Protein Name | MW (Da) | PI | Subcellular Location | Biological Process | Molecular Function | Peptide |
|---|---|---|---|---|---|---|---|---|
| SULT1A3/1A4 | P50224 | Sulfotransferase 1A3/1A4 | 34,174 | 5.68 | Cytoplasm | Catecholamine metabolism; Lipid metabolism; Steroid metabolism | Transferase | R.LIKSHLPLALLPQTLLDQK.V; R.LIKSHLPLALLPQTLLDQK.V + Deamidated (NQ); R.LIKSHLPLALLPQTLLDQK.V + Deamidated (NQ); R.LIKSHLPLALLPQTLLDQK.V + 2 Deamidated (NQ) |
| ACIN1 | Q9UKV3 | Apoptotic chromatin condensation inducer in the nucleus | 151,771 | 6.08 | Nucleus; Nucleoplasm | Apoptosis; mRNA processing; mRNA splicing | ATPase activity; enzyme binding; nucleic acid binding; nucleotide binding; poly(A) RNA binding | R.EREMER.R; R.TSTSSSSVQAR.R + 7 Phospho (ST); K.QSADSSSSRSSSSSSSSSR.S + Deamidated (NQ); 9 Phospho (ST); K.QSADSSSSRSSSSSSSSSR.S + Deamidated (NQ); 10 Phospho (ST) |
| ProSAPiP1 | O60299 | ProSAP-interacting protein 1 | 71,747 | 7.56 | Cytoplasm; Cytoskeleton | K.SRTMTPAGGSGSGLSDSGR.N + Oxidation (M); K.SRTMTPAGGSGSGLSDSGR.N + Oxidation (M); 4 Phospho (ST); R.IGTASYGSGSGGSSGGGSGYQDLGTSDSGR.A + 4 Phospho (ST); Phospho (Y); K.SRTMTPAGGSGSGLSDSGR.N + Oxidation (M); 4 Phospho (ST); K.QLQLSYVEMYQRNQQLER.R + 3 Deamidated (NQ); Phospho (Y) | ||
| PDE5A | O76074 | cGMP-specific 3',5'-cyclic phosphodiesterase | 99,921 | 5.74 | blood coagulation; cGMP catabolic process; negative regulation of T cell proliferation; negative regulation of cardiac muscle contraction; positive regulation of MAP kinase activity; positive regulation of cardiac muscle hypertrophy; positive regulation of oocyte development; relaxation of cardiac muscle; signal transduction | Hydrolase | R.WILSVKKNYR.K + Phospho (ST); K.KIAATIISFMQVQK.C + Oxidation (M); Phospho (ST); K.ELNIEPTDLMNREKK.N + Deamidated (NQ); K.TQSILCMPIKNHREEVVGVAQAINK.K + 4 Deamidated (NQ); 2 Phospho (ST); R.GHTESCSCPLQQSPRADNSAPGTPTRK.I + 2 Deamidated (NQ); 5 Phospho (ST) | |
| ARID1A | O14497 | AT-rich interactive domain-containing protein 1A | 241,892 | 6.24 | Nucleus | Neurogenesis; Transcription; Transcription regulation | Chromatin regulator | K.SKKSSSSTTTNEK.I + Deamidated (NQ); 6 Phospho (ST); K.HPGLLLILGKLILLHHK.H; R.NSMTPNPGYQPSMNTSDMMGR.M + 2 Deamidated (NQ); Oxidation (M); 2 Phospho (ST); R.EMAVVLLANLAQGDSLAARAIAVQK.G + Deamidated (NQ); Oxidation (M); R.EMAVVLLANLAQGDSLAARAIAVQK.G + Deamidated (NQ); Oxidation (M); R.ITATMDDMLSTRSSTLTEDGAK.S + 2 Oxidation (M); 4 Phospho (ST); K.APGSDPFMSSGQGPNGGMGDPYSR.A + 4 Phospho (ST); Phospho (Y); R.GYMQRNPQMPQYSSPQPGSALSPR.Q + Oxidation (M); Phospho (ST); K.RNSMTPNPGYQPSMNTSDMMGR.M + Deamidated (NQ); Oxidation (M); 3 Phospho (ST); Phospho (Y) |
| BIRC6 | Q9NR09 | Baculoviral IAP repeat-containing protein 6 | 529,919 | 5.67 | Golgi apparatus; Cytoplasm; Cytoskeleton | Apoptosis; Cell cycle; Cell division; Mitosis; Ubl conjugation pathway | Ligase; Protease inhibitor; Thiol protease inhibitor | K.KTSISKER.V + 2 Phospho (ST); R.YGSTNARAK.I + Deamidated (NQ); 2 Phospho (ST); R.SRGTPSGTQSSR.E + Deamidated (NQ); 3 Phospho (ST); K.MKTCVDTYTNR.L + Deamidated (NQ); Phospho (ST); Phospho (Y); R.TIPDKIGSTSGAEAANK.I + Deamidated (NQ); R.QLQDRLTPMEALLQTR.Y + Deamidated (NQ); Oxidation (M); R.GRTIPDKIGSTSGAEAANK.I + Phospho (ST); K.EKSSNVKNENTSGTR.K + 3 Deamidated (NQ); 5 Phospho (ST); K.LVNILVQLPLSGNREYSAR.V; K.WNSVFPKPGTLVQCLRLPK.F + Carbamidomethyl (C); Deamidated (NQ); K.VNYHYMSQVKNANDANSAAR.A + 5 Deamidated (NQ); Phospho (ST); R.LAQEAVTLSTSLPLSSSSSVFVR.C + Deamidated (NQ); Phospho (ST); R.GTEEICNGGMRPVVRLPSLKHQSNK.G + Carbamidomethyl (C); Deamidated (NQ); R.SFLIHVKAVNERGTEEICNGGMRPVVR.L + Deamidated (NQ); Phospho (ST) |
| CCNB2 | O95067 | G2/mitotiC-specific cyclin-B2 | 45,253 | 9 | Cell cycle; Cell division; Mitosis | Cyclin | R.KKLQLVGITALLLASK.Y; K.VPVQPTKTTNVNKQLKPTASVKPVQMEK.L + Deamidated (NQ); Oxidation (M); Phospho (ST); K.AQNTKVPVQPTKTTNVNK.Q + 3 Deamidated (NQ); 2 Phospho (ST) | |
| CEP250 | Q9BV73 | Centrosome-associated protein CEP250 | 280,967 | 5 | Cytoplasm; Cytoskeleton | Cell cycle | protein C-terminus binding; protein kinase binding | R.EPAQLLLLLAK.T; K.GQLEVQIQTVTQAK.E + 4 Deamidated (NQ); Phospho (ST); R.QLMQERAEEGKGPSK.A + 2 Deamidated (NQ); K.ELSAQMELLRQEVK.E + Oxidation (M); Phospho (ST); R.GLHQSVRELQLTLAQK.E + Deamidated (NQ); Phospho (ST); R.DQELEALQQEQQQAQGQEER.V + 5 Deamidated (NQ); K.CVAELQKEVVLLQAQLTLERK.Q + Deamidated (NQ); K.AEHVRLSGSLLTCCLRLTVGAQSR.E; R.QEQTEVTAALAR.A + 2 Deamidated (NQ); R.RVNVELQLQGDSAQGQK.E + 2 Deamidated (NQ); Phospho (ST); K.AEHVRLSGSLLTCCLRLTVGAQSR.E; R.SLFKRGPLLTALSAEAVASALHK.L + 3 Phospho (ST) |
| AGBL1 | Q96MI9 | Cytosolic carboxypeptidase 4 | 120,204 | 6.85 | Cytoplasm; Cytosol | C-terminal protein deglutamylation; protein side chain deglutamylation | Carboxypeptidase; Hydrolase; Metalloprotease; Protease | R.MSASFSNSTRTR.E + Deamidated (NQ); 4 Phospho (ST); K.LAPAFTMSSCSFLVEKSR.A; R.SYTMESSYCGCNQGPYQCTQR.L + Deamidated (NQ); Oxidation (M); 4 Phospho (ST); 2 Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + Deamidated (NQ); Oxidation (M); 4 Phospho (ST); 2 Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + 2 Deamidated (NQ); Oxidation (M); 4 Phospho (ST); 2 Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + 3 Deamidated (NQ); Oxidation (M); 4 Phospho (ST); 2 Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + 2 Carbamidomethyl (C); 2 Deamidated (NQ); 3 Phospho (ST); 2 Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + 2 Carbamidomethyl (C); 2 Deamidated (NQ); Oxidation (M); 4 Phospho (ST); Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + 2 Carbamidomethyl (C); 2 Deamidated (NQ); Oxidation (M); 4 Phospho (ST); Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + 2 Carbamidomethyl (C); 2 Deamidated (NQ); Oxidation (M); 4 Phospho (ST); Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + 2 Carbamidomethyl (C); 2 Deamidated (NQ); Oxidation (M); 3 Phospho (ST); 2 Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + 2 Carbamidomethyl (C); 2 Deamidated (NQ); Oxidation (M); 3 Phospho (ST); 2 Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + 2 Carbamidomethyl (C); 2 Deamidated (NQ); Oxidation (M); 3 Phospho (ST); 2 Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + 2 Carbamidomethyl (C); 3 Deamidated (NQ); Oxidation (M); 2 Phospho (ST); 3 Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + 2 Carbamidomethyl (C); 3 Deamidated (NQ); Oxidation (M); 4 Phospho (ST); Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + 2 Carbamidomethyl (C); 3 Deamidated (NQ); Oxidation (M); 4 Phospho (ST); Phospho (Y); R.SYTMESSYCGCNQGPYQCTQR.L + 2 Carbamidomethyl (C); Oxidation (M); 5 Phospho (ST); Phospho (Y); R.EMGVSRSYTMESSYCGCNQGPYQCTQR.L + 2 Carbamidomethyl (C); Oxidation (M); 2 Phospho (ST); 2 Phospho (Y) |


3. Experimental Section
3.1. Silk Fibroin Nanoparticle Preparation
3.2. Characterization of Silk Fibroin Nanoparticles
3.3. Cell Culture
3.4. Bromodeoxyuridine (5-Bromo-2'-deoxyuridine, BrdU) and Lactate Dehydrogenase (LDH) Assay
3.5. Cell Morphology
3.6. Protein Sample Preparation
3.7. Proteomic Analysis
3.8. Database Search
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ayoub, N.A.; Garb, J.E.; Tinghitella, R.M.; Collin, M.A.; Hayashi, C.Y. Blueprint for a high-performance biomaterial: Full-length spider dragline silk genes. PLoS One 2007, 2, e514. [Google Scholar] [CrossRef] [PubMed]
- Hakimi, O.; Knight, D.P.; Vollrath, F.; Vadgama, P. Spider and mulberry silkworm silks as compatible biomaterials. Composites 2007, 38, 324–337. [Google Scholar] [CrossRef]
- Dyakonov, T.; Yang, C.H.; Bush, D.; Gosangari, S.; Majuru, S.; Fatmi, A. Design and characterization of a silk-fibroin-based drug delivery platform using naproxen as a model drug. J. Drug Deliv. 2012, 2012, 490514. [Google Scholar] [CrossRef] [PubMed]
- Chi, N.H.; Yang, M.C.; Chung, T.W.; Chou, N.K.; Wang, S.S. Cardiac repair using chitosan-hyaluronan/silk fibroin patches in a rat heart model with myocardial infarction. Carbohydr. Polym. 2013, 92, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Um, I.C.; Kweon, H.; Park, Y.H.; Hudson, S. Structural characteristics and properties of the regenerated silk fibroin prepared from formic acid. Int. J. Biol. Macromol. 2001, 29, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Reagan, M.R.; Kaplan, D.L. Electrospun silk biomaterial scaffolds for regenerative medicine. Adv. Drug Deliv. Rev. 2009, 61, 988–1006. [Google Scholar] [CrossRef] [PubMed]
- Guziewicz, N.; Best, A.; Perez-Ramirez, B.; Kaplan, D.L. Lyophilized silk fibroin hydrogels for the sustained local delivery of therapeutic monoclonal antibodies. Biomaterials 2011, 32, 2642–2650. [Google Scholar] [CrossRef] [PubMed]
- Rajkhowa, R.; Gil, E.S.; Kludge, J.A.; Numata, K.; Wang, L.; Wang, X.; Kaplan, D.L. Reinforcing silk scaffolds with silk particles. Macromol. Biosci. 2010, 10, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Soffer, L.; Wang, X.; Zhang, X.; Kluge, J.; Dorfmann, L.; Kaplan, D.L.; Leisk, G. Silk-based electrospun tubular scaffolds for tissue-engineered vascular grafts. J. Biomater. Sci. Polym. Ed. 2008, 19, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, X.; Ding, F.; Zhang, P.; Liu, J.; Gu, X. Biocompatibility evaluation of silk fibroin with peripheral nerve tissues and cells in vitro. Biomaterials 2007, 28, 1643–1652. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Vepari, C.; Jin, H.J.; Kim, H.J.; Kaplan, D.L. Electrospun silk-BMP-2 scaffolds for bone tissue engineering. Biomaterials 2006, 27, 3115–3124. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Blasioli, D.J.; Kim, H.J.; Kim, H.S.; Kaplan, D.L. Cartilage tissue engineering with silk scaffolds and human articular chondrocytes. Biomaterials 2006, 27, 4434–4442. [Google Scholar] [CrossRef] [PubMed]
- Omenetto, F.G.; Kaplan, D.L. New opportunities for an ancient material. Science 2010, 329, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Kasemo, B. Biological surface science. Surf. Sci. 2002, 500, 656–677. [Google Scholar] [CrossRef]
- Lutolf, M.P.; Hubbell, J.A. Synthetic biomaterials as instructive extracellular microenvironments for morphogenesis in tissue engineering. Nat. Biotechnol. 2005, 23, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Aseh, A.; Ríos, C.N.; Aggarwal, B.B.; Mathur, A.B. Fabrication and characterization of silk fibroin-derived curcumin nanoparticles for cancer therapy. Int. J. Nanomed. 2009, 4, 115–122. [Google Scholar] [CrossRef]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef] [PubMed]
- Zetter, B.R. Angiogenesis and tumor metastasis. Annu. Rev. Med. 1998, 49, 407–424. [Google Scholar] [CrossRef] [PubMed]
- Danhier, F.; Feron, O.; Préat, V. To exploit the tumor microenvironment: Passive and active tumor targeting of nanocarriers for anti-cancer drug delivery. J. Control. Release 2010, 148, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Van der Lubben, I.M.; Verhoef, J.C.; Borchard, G.; Junginger, H.E. Chitosan for mucosal vaccination. Adv. Drug Deliv. Rev. 2001, 52, 139–144. [Google Scholar] [CrossRef] [PubMed]
- L’opez-Le’on, T.; Carvalho, E.L.S.; Seijo, B.; Ortega-Vinuesa, J.L.; Bastos-Gonz’alez, D. Physicochemical characterization of chitosan nanoparticles: electrokinetic and stability behavior. J. Colloid Interface Sci. 2005, 283, 344–351. [Google Scholar]
- Wilkins, M.R.; Sanchez, J.C.; Williams, K.L.; Hochstrasser, D.F. Current challenges and future applications for protein maps and post-translational vector maps in proteome projects. Electrophoresis 1996, 17, 830–838. [Google Scholar] [CrossRef] [PubMed]
- Mani, A.; Gelmann, E.P. The ubiquitin-proteasome pathway and its role in cancer. J. Clin. Oncol. 2005, 23, 4776–4789. [Google Scholar] [CrossRef] [PubMed]
- Pickart, C.M.; Eddins, M.J. Ubiquitin: Structures, functions, mechanisms. Biochim. Biophys. Acta 2004, 1695, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, O.D.; Berkers, C.R.; Mason, S.M.; Zheng, L.; Blyth, K.; Gottlieb, E.; Vousden, K.H. Serine starvation induces stress and p53-dependent metabolic remodelling in cancer cells. Nature 2013, 493, 542–546. [Google Scholar] [CrossRef] [PubMed]
- Geske, F.J.; Nelson, A.C.; Lieberman, R.; Strange, R.; Sun, T.; Gerschenson, L.E. DNA repair is activated in early stages of p53-induced apoptosis. Cell Death Differ. 2000, 7, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Bensaad, K.; Vousden, K.H. p53: New roles in metabolism. Trends Cell. Biol. 2007, 17, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Cheema, S.K.; Gobin, A.S.; Rhea, R.; Lopez-Berestein, G.; Newman, R.A.; Mathur, A.B. Silk fibroin mediated delivery of liposomal emodin to breast cancer cells. Int. J. Pharm. 2007, 341, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Pinedo, C.; El Mjiyad, N.; Ricci, J.E. Cancer metabolism: Current perspectives and future directions. Cell Death Dis. 2012, 3, e248. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, E.; Ciraolo, E.; Franco, I.; Ghigo, A.; Martini, M. PI3K in cancer–stroma interactions: Bad in seed and ugly in soil. Oncogene 2014, 33, 3083–3090. [Google Scholar] [CrossRef] [PubMed]
- Finley, L.W.S.; Zhang, J.; Ye, J.; Ward, P.S.; Thompson, C.B. SnapShot: Cancer metabolism pathways. Cell Metab. 2013, 17, 466–466. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yuan, S.S.; Chung, T.W.; Jong, S.B.; Lu, C.Y.; Tsai, W.C.; Chen, W.C.; Lin, P.C.; Chiang, P.W.; Tyan, Y.C. Characterization of silk fibroin modified surface: A proteomic view of cellular response proteins induced by biomaterials. BioMed. Res. Int. 2014, 2014, 209469. [Google Scholar] [PubMed]
- Calvo, P.; Remuñan-López, C.; Vila-Jato, J.L.; Alonso, M.J. Chitosan and chitosan/ethylene oxide-propylene oxide block copolymer nanoparticles as novel carriers for proteins and vaccines. Pharm. Res. 1997, 14, 1431–1436. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.-H.; Chung, T.-W.; Lu, Y.-S.; Chen, Y.-L.; Tsai, W.-C.; Jong, S.-B.; Yuan, S.-S.; Liao, P.-C.; Lin, P.-C.; Tyan, Y.-C. Activation of the Ubiquitin Proteasome Pathway by Silk Fibroin Modified Chitosan Nanoparticles in Hepatic Cancer Cells. Int. J. Mol. Sci. 2015, 16, 1657-1676. https://doi.org/10.3390/ijms16011657
Yang M-H, Chung T-W, Lu Y-S, Chen Y-L, Tsai W-C, Jong S-B, Yuan S-S, Liao P-C, Lin P-C, Tyan Y-C. Activation of the Ubiquitin Proteasome Pathway by Silk Fibroin Modified Chitosan Nanoparticles in Hepatic Cancer Cells. International Journal of Molecular Sciences. 2015; 16(1):1657-1676. https://doi.org/10.3390/ijms16011657
Chicago/Turabian StyleYang, Ming-Hui, Tze-Wen Chung, Yi-Shan Lu, Yi-Ling Chen, Wan-Chi Tsai, Shiang-Bin Jong, Shyng-Shiou Yuan, Pao-Chi Liao, Po-Chiao Lin, and Yu-Chang Tyan. 2015. "Activation of the Ubiquitin Proteasome Pathway by Silk Fibroin Modified Chitosan Nanoparticles in Hepatic Cancer Cells" International Journal of Molecular Sciences 16, no. 1: 1657-1676. https://doi.org/10.3390/ijms16011657