
Proteome Analysis of Dormancy-Released Seeds of Fraxinus mandshurica Rupr. in Response to Re-Dehydration under Different Conditions

Abstract
:
1. Introduction
2. Results and Discussion




{kind=link}
{kind=link}
{kind=link}
{kind=link}
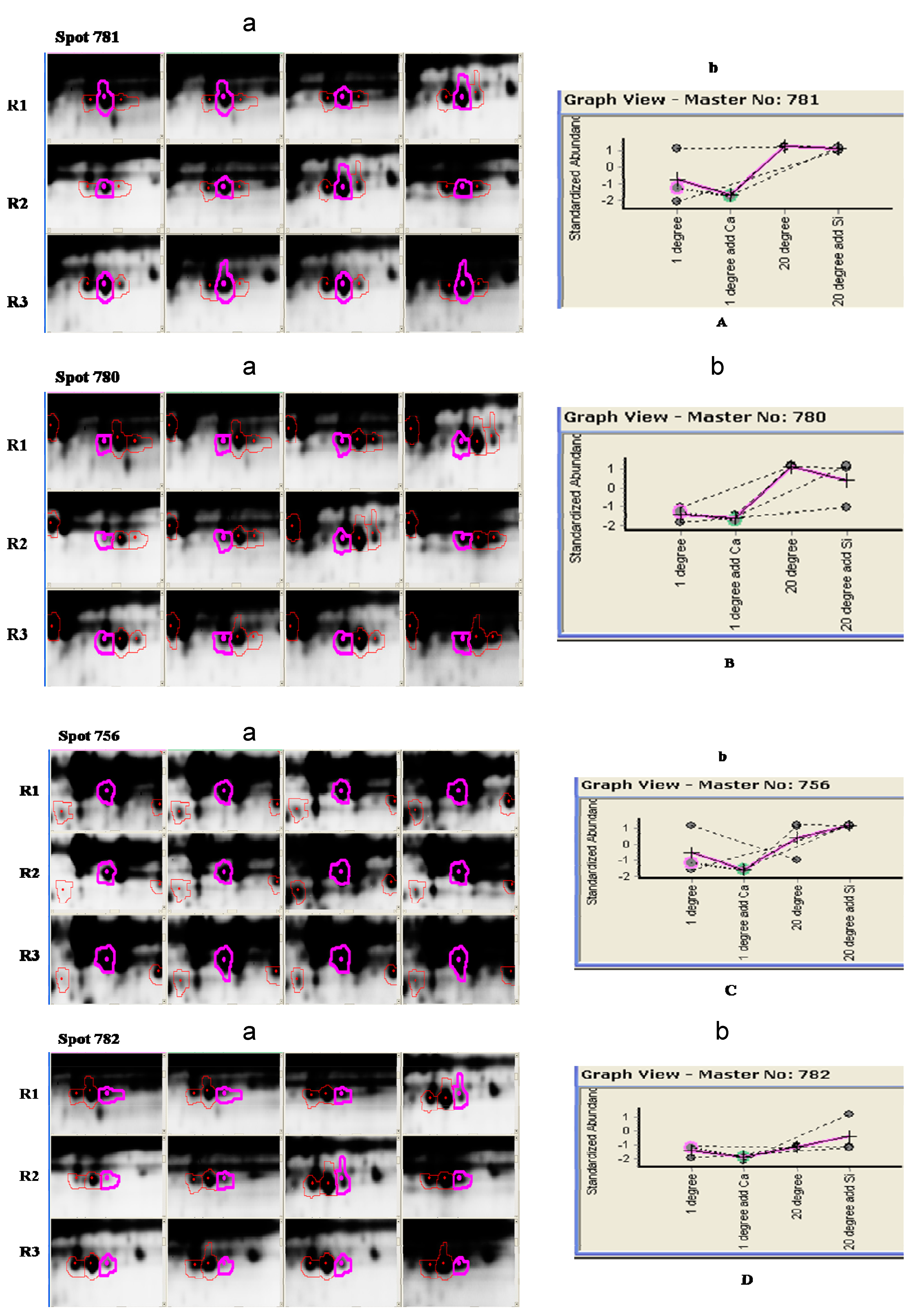{kind=link}
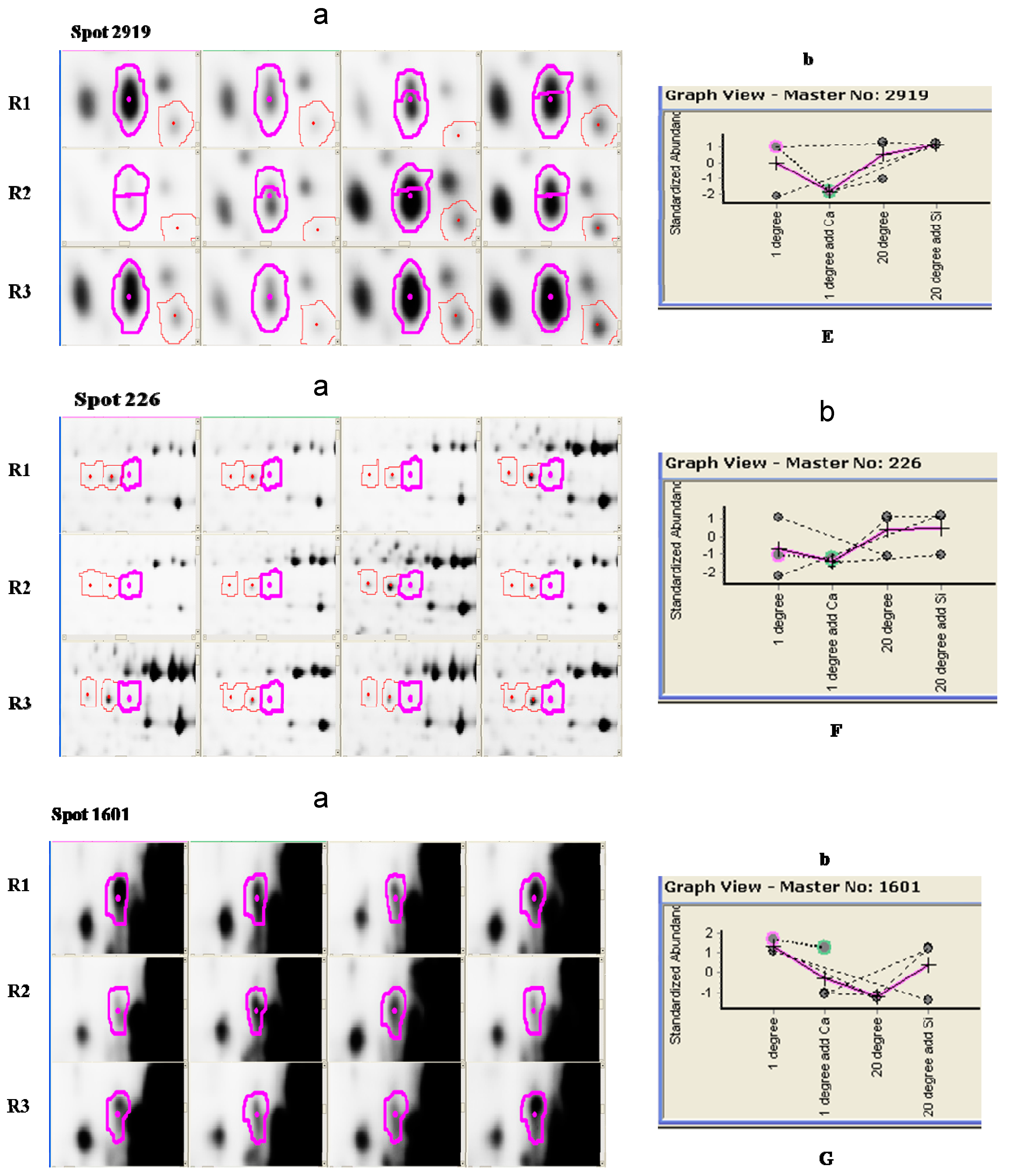{kind=link}
| Spot No. | Protein Name | Species | Accession No. | Protein Mw | Protein PI | Protein Score C.I. % |
|---|---|---|---|---|---|---|
| 226 | Putative acyl-coenzyme A oxidase 3.2, peroxisomal | Arabidopsis thaliana | sp|Q9LMI7|ACO32_ARATH | 76,479 | 7.93 | 86.829 |
| 756 | Catalase isozyme 3 | Nicotiana plumbaginifolia | sp|P49317|CATA3_NICPL | 57,493.5 | 6.78 | 100.000 |
| 780 | Catalase isozyme 1 | Ricinus communis | sp|Q01297|CATA1_RICCO | 56,941.5 | 7.08 | 100.000 |
| 781 | Catalase isozyme 3 | Nicotiana plumbaginifolia | sp|P49317|CATA3_NICPL | 57,493.5 | 6.78 | 100.000 |
| 782 | Catalase isozyme 3 | Nicotiana plumbaginifolia | sp|P49317|CATA3_NICPL | 57,493.5 | 6.78 | 100.000 |
| 1601 | Kinesin-1 | Arabidopsis thaliana | sp|Q07970|ATK1_ARATH | 89,676.6 | 6.80 | 89.294 |
| 2919 | Nucleoside diphosphate kinase | Capsicum annuum | sp|Q9M7P6|NDK_CAPA | 16,372.5 | 6.31 | 99.488 |
2.2. Proteome Responses of the Cellular Protective Program Associated with Different Treatments
2.2.1. Changes in Energy Metabolism (Transport-Associated Proteins)


2.2.2. Defense Mechanisms
3. Experimental Section
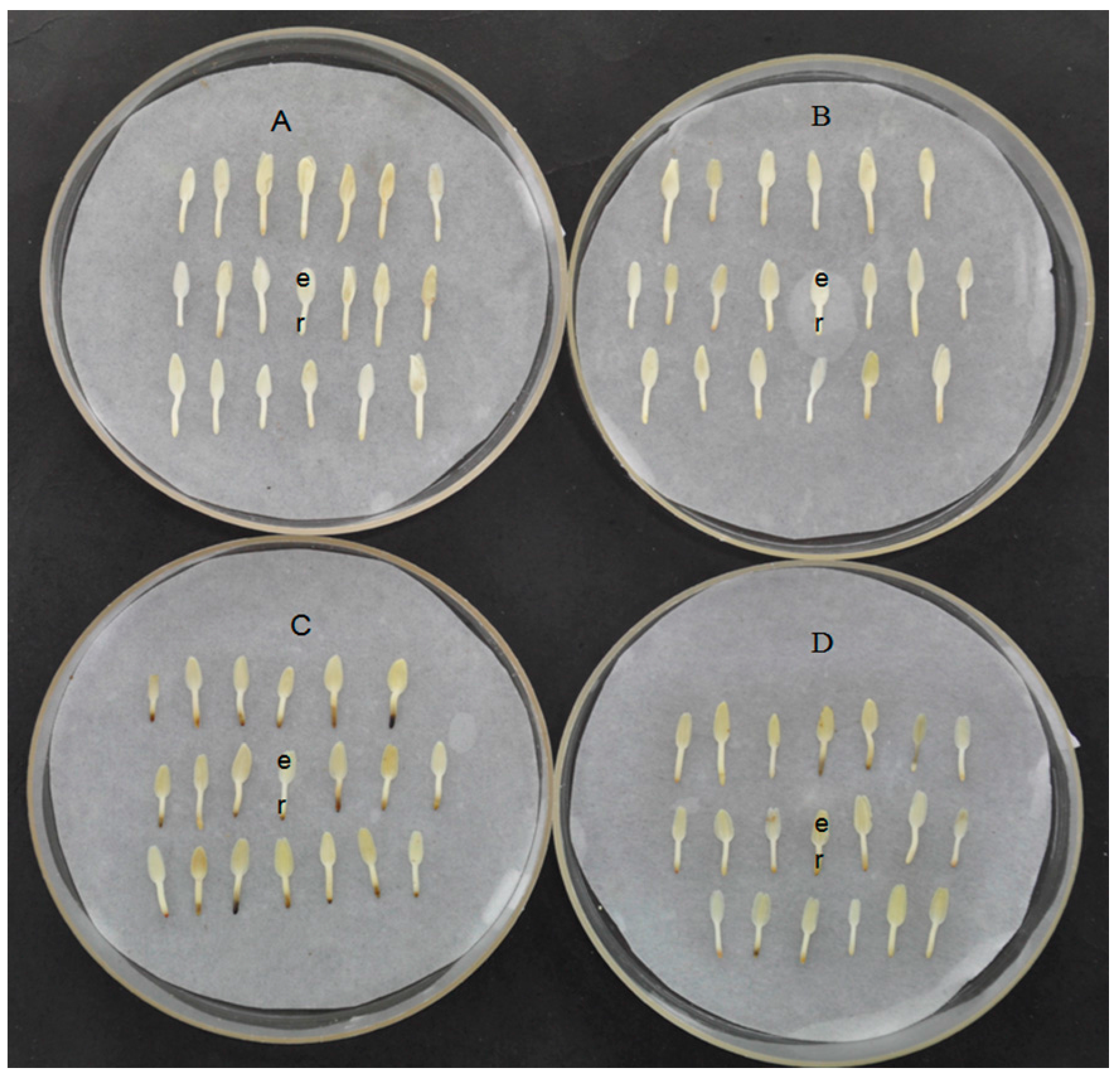
3.1. Plant Material and Treatments
3.2. Protein Extraction
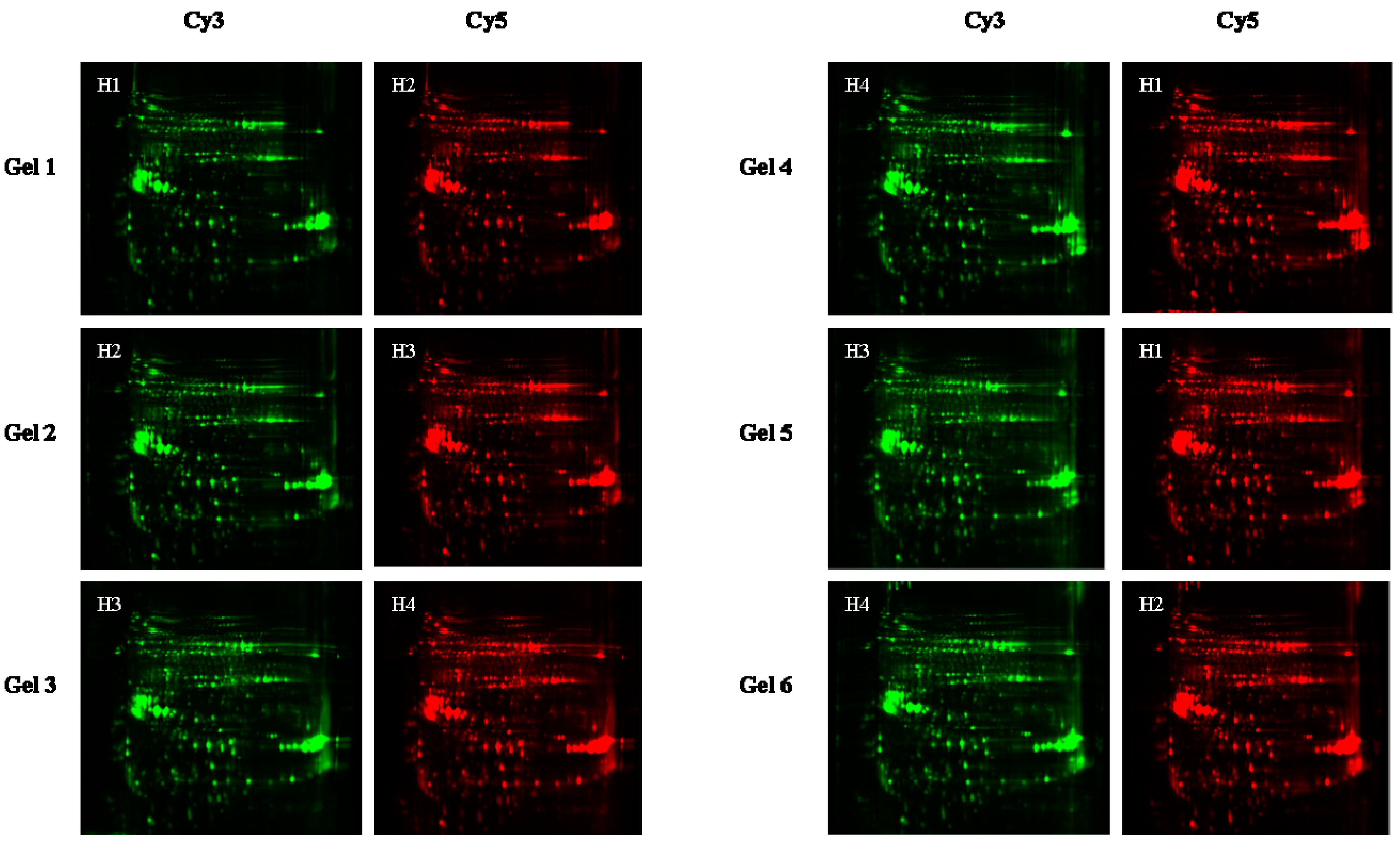
3.3. Protein Isolation and 2D-DIGE
3.4. Image Acquisition and 2D-DIGE Image Analysis
3.5. Identification of Proteins by MALDI TOF/TOF MS
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wu, L.D. Germination Physiology of Dormancy-Released Seeds of Manchurian ash (Fraxinus mandshurica L.). Dehydrated in Different Condition. Master’s Dissertation, Northeast Forestry University, Harbin, China, 2012. [Google Scholar]
- Alpert, P. Constraints of tolerance: Why are desiccation-tolerant organisms so small or rare? J. Exp. Biol. 2006, 209, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Proctor, M.C.; Oliver, M.J.; Wood, A.J.; Alpert, P.; Stark, L.R.; Cleavitt, N.L.; Mishler, B.D. Desiccation-tolerance in bryophytes: A review. Bryologist 2007, 110, 595–621. [Google Scholar] [CrossRef]
- Wood, A.J. Eco-hysiological adaptations to limited water enviornments. Plant Abiotic Stress 2005, 1–13. [Google Scholar] [CrossRef]
- Leprince, O.; Buitink, J. Desiccation tolerance: From genomics to the field. Plant Sci. 2010, 179, 554–564. [Google Scholar] [CrossRef] [Green Version]
- Fait, A.; Angelovici, R.; Less, H.; Ohad, I.; Urbanczyk-Wochniak, E.; Fernie, A.R.; Galili, G. Arabidopsis seed development and germination is associated with temporally distinct metabolic switches. Plant Physiol. 2006, 142, 839–854. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, K.; Firnhaber, C.; Zuber, H.; Héricher, D.; Belghazi, M.; Henry, C.; Küster, H.; Thompson, R. A combined proteome and transcriptome analysis of developing Medicago truncatula seeds. Mol. Cell. Proteomics 2007, 6, 2165–2179. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, L.; van Wuytswinkel, O.; Castelain, M.; Bellini, C. Combined networks regulating seed maturation. Trends Plant Sci. 2007, 12, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Hajduch, M.; Ganapathy, A.; Stein, J.W.; Thelen, J.J. A systematic proteomic study of seed filling in soybean. Establishment of high-resolution two-dimensional reference maps, expression profiles, and an interactive proteome database. Plant Physiol. 2005, 137, 1397–1419. [Google Scholar]
- Holdsworth, M.J.; Finch-Savage, W.E.; Grappin, P.; Job, D. Post-genomics dissection of seed dormancy and germination. Trends Plant Sci. 2008, 13, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Poole, R.; Huttly, A.; Toscano-Underwood, C.; Feeney, K.; Welham, S.; Gooding, M.; Mills, C.; Edwards, K.; Shewry, P. Transcriptome analysis of grain development in hexaploid wheat. BMC Genomics 2008, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoekstra, F.A.; Golovina, E.A.; Buitink, J. Mechanisms of plant desiccation tolerance. Trends Plant Sci. 2001, 6, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Soeda, Y.; Konings, M.C.J.M.; Vorst, O.; van Houwelingen, A.M.M.L.; Stoopen, G.M.; Maliepaard, C.A.; Kodde, J.; Bino, R.J.; Groot, S.P.C.; van der Geest, A.H.M. Gene expression programs during Brassica oleracea seed maturation, osmopriming, and germination are indicators of progression of the germination process and the stress tolerance level. Plant Physiol. 2005, 137, 354–368. [Google Scholar] [CrossRef] [PubMed]
- Boudet, J.; Buitink, J.; Hoekstra, F.A.; Rogniaux, H.; Larre, C.; Satour, P.; Leprince, O. Comparative analysis of the heat stable proteome of radicles of Medicago truncatula seeds during germination identifies late embryogenesis abundant proteins associated with desiccation tolerance. Plant Physiol. 2006, 140, 1418–1436. [Google Scholar] [CrossRef] [PubMed]
- Buitink, J.; Leger, J.J.; Guisle, I.; Vu, B.L.; Wuillème, S.; Lamirault, G.; Bars, A.L.; Meur, N.L.; Becker, A.; Kuster, H.; et al. Transcriptome profiling uncovers metabolic and regulatory processes occurring during the transition from desiccation-sensitive to desiccation-tolerant stages in Medicago truncatula seeds. Plant J. 2006, 47, 735–750. [Google Scholar]
- Buitink, J.; Ly Vu, B.; Satour, P.; Leprince, O. The re-establishment of desiccation tolerance in germinated radicles of Medicago truncatula Gaertn. seeds. Seed Sci. Res. 2003, 13, 273–286. [Google Scholar] [CrossRef]
- Mann, M.; Jensen, O.N. Proteomic analysis of post-translational modifications. Nat. Biotechnol. 2003, 21, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Schweppe, R.E.; Haydon, C.E.; Lewis, T.S.; Resing, K.A.; Ahn, N.G. The characterization of protein post-translational modifications by mass spectrometry. Acc. Chem. Res. 2003, 36, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Amme, S.; Matros, A.; Schlesier, B.; Mock, H.P. Proteome analysis of cold stress response in Arabidopsis thaliana using DIGE-technology. J. Exp. Bot. 2006, 57, 1537–1546. [Google Scholar] [CrossRef] [PubMed]
- Balbuena, T.S.; Jo, L.; Pieruzzi, F.P.; Dias, L.L.C.; Silveira, V.; Santa-Catarina, C.; Junqueira, M.; Thelen, J.J.; Shevchenko, A.; Floh, E.I.S. Differential proteome analysis of mature and germinated embryos of Araucaria angustifolia. Phytochemistry 2011, 72, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Chien, C.T.; Lin, C.H.; Chiu, Y.Y.; Yang, Y.S. Protein changes between dormant and dormancy-broken seeds of Prunus campanulata Maxim. Proteomics 2006, 6, 4147–4154. [Google Scholar] [CrossRef] [PubMed]
- Pawlowski, T.A. Proteomics of European beech (Fagus sylvatica L.) seed dormancy breaking: Influence of abscisic and gibberellic acids. Proteomics 2007, 7, 2246–2257. [Google Scholar]
- Sghaier-Hammami, B.; Valledor, L.; Drira, N.; Jorrin-Novo, J.V. Proteomic analysis of the development and germination of date palm (Phoenix dactylifera L.) zygotic embryos. Proteomics 2009, 9, 2543–2554. [Google Scholar] [CrossRef]
- Yang, M.F.; Liu, Y.J.; Liu, Y.; Chen, H.; Chen, F.; Shen, S.H. Proteomic analysis of oil mobilization in seed germination and postgermination development of Jatropha curcas. J. Proteome Res. 2009, 8, 1441–1451. [Google Scholar] [CrossRef] [PubMed]
- Marsoni, M.; Bracale, M.; Espen, L.; Prinsi, B.; Negri, A.S.; Vannini, C. Proteomic analysis of somatic embryogenesis in Vitis vinifera. Plant Cell Rep. 2008, 27, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Guan, R.; Zhu, S.; Deng, X. Proteomic analysis of somatic embryogenesis in Valencia sweet orange (Citrus sinensis Osbeck). Plant Cell Rep. 2009, 28, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Sghaier-Hammami, B.; Jorrin-Novo, J.V.J.; Gargouri-Bouzid, R.; Drira, N. Abscisic acid and sucrose increase the protein content in date palm somatic embryos, causing changes in 2-DE profile. Phytochemistry 2010, 71, 1223–1236. [Google Scholar] [CrossRef] [PubMed]
- Blackman, S.A.; Obendorf, R.L.; Leopold, A.C. Maturation proteins and sugars in desiccation tolerance of developing soybean seeds. Plant Physiol. 1992, 100, 225. [Google Scholar] [CrossRef] [PubMed]
- Etienne, H.; Montoro, P.; Michaux-Ferriere, N.; Carron, M. Effects of desiccation, medium osmolarity and abscisic acid on the maturation of Hevea brasiliensis somatic embryos. J. Exp. Bot. 1993, 44, 1613–1619. [Google Scholar] [CrossRef]
- Gilroy, S.; Bethke, P.C.; Jones, R.L. Calcium homeostasis in plants. J. Cell Sci. 1993, 106, 453–461. [Google Scholar] [PubMed]
- Bush, D.S. Regulation of cytosolic calcium in plants. Plant Physiol. 1993, 103, 7–13. [Google Scholar] [PubMed]
- Illing, N.; Denby, K.J.; Collett, H.; Shen, A.; Farrant, J.M. The signature of seeds in resurrection plants: a molecular and physiological comparison of desiccation tolerance in seeds and vegetative tissues. Integr. Comp. Biol. 2005, 45, 771–787. [Google Scholar] [CrossRef] [PubMed]
- Cuming, A. LEA Proteins. Seed Proteins; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1999; pp. 753–780. [Google Scholar]
- Bae, M.S.; Cho, E.J.; Choi, E.; Park, O.K. Analysis of the Arabidopsis nuclear proteome and its response to cold stress. Plant J. 2003, 36, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Adio, S.; Reth, J.; Bathe, F.; Woehlke, G. Review: Regulation mechanisms of Kinesin-1. J. Muscle Res. Cell Motil. 2006, 27, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Marcus, A.; Li, W.; Hu, Y.; Calzada, J.P.V.; Grossniklaus, U.; Cyr, R.J.; Ma, H. The Arabidopsis ATK1 gene is required for spindle morphogenesis in male meiosis. Development 2002, 129, 2401–2409. [Google Scholar] [CrossRef] [PubMed]
- Marcus, A.I.; Ambrose, J.C.; Blickley, L.; Hancock, W.O.; Cyr, R.J. Arabidopsis thaliana protein, ATK1, is a minus-end directed kinesin that exhibits non-processive movement. Cell Motil. Cytoskelet. 2002, 52, 144–150. [Google Scholar] [CrossRef]
- Lee, Y.R.J.; Liu, B. Cytoskeletal motors in Arabidopsis. Sixty-one kinesins and seventeen myosins. Plant Physiol. 2004, 136, 3877–3883. [Google Scholar]
- Navone, F.; Niclas, J.; Hom-Booher, N.; Sparks, L.; Bernstein, H.D.; McCaffrey, G.; Vale, R.D. Cloning and expression of a human kinesin heavy chain gene: Interaction of the COOH-terminal domain with cytoplasmic microtubules in transfected CV-1 cells. J. Cell Biol. 1992, 117, 1263–1275. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.T.; Saxton, W.M.; Stewart, R.J.; Raff, E.C.; Goldstein, L. Evidence that the head of kinesin is sufficient for force generation and motility in vitro. Science 1990, 249, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Sloboda, R.D.; Rosenbaum, J.L. Decoration and stabilization of intact, smooth-walled microtubules with microtubule-associated proteins. Biochemistry 1979, 18, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Yan, X.; Li, X.; Guo, G.; Hu, Y.; Ma, W.; Yan, Y. Proteome analysis of wheat leaf under salt stress by two-dimensional difference gel electrophoresis (2D-DIGE). Phytochemistry 2011, 72, 1180–1191. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.C.; Peng, Z.Y.; Li, C.L.; Li, F.; Liu, C.; Xia, G.M. Proteomic analysis on a high salt tolerance introgression strain of Triticum aestivum/Thinopyrum ponticum. Proteomics 2008, 8, 1470–1489. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.P.; Zhang, Q.Y.; Tang, Z.C.; Su, W.A.; Sun, W.N. Comparative proteomic analysis provides new insights into chilling stress responses in rice. Mol. Cell. Proteomics 2006, 5, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Canovas, F.M.; Dumas-Gaudot, E.; Recorbet, G.; Jorrin, J.; Mock, H.P.; Rossignol, M. Plant proteome analysis. Proteomics 2004, 4, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Chitteti, B.R.; Peng, Z. Proteome and phosphoproteome differential expression under salinity stress in rice (Oryza sativa) roots. J. Proteome Res. 2007, 6, 1718–1727. [Google Scholar] [CrossRef] [PubMed]
- Feierabend, J.; Schaan, C.; Hertwig, B. Photoinactivation of catalase occurs under both high-and low-temperature stress conditions and accompanies photoinhibition of photosystem II. Plant Physiol. 1992, 100, 1554. [Google Scholar] [CrossRef] [PubMed]
- Schöner, S.; Krause, G.H. Protective systems against active oxygen species in spinach: Response to cold acclimation in excess light. Planta 1990, 180, 383–389. [Google Scholar] [CrossRef] [PubMed]
- De Bellis, L.; Picciarelli, P.; Pistelli, L.; Alpi, A. Localization of glyoxylate-cycle marker enzymes in peroxisomes of senescent leaves and green cotyledons. Planta 1990, 180, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Dorion, S.; Matton, D.P.; Rivoal, J. Characterization of a cytosolic nucleoside diphosphate kinase associated with cell division and growth in potato. Planta 2006, 224, 108–124. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.; Yi, H.; Lee, J.; Kwon, Y.K.; Soh, M.S.; Shin, B.; Luka, Z.; Hahn, T.R.; Song, P.S. Phytochrome signalling is mediated through nucleoside diphosphate kinase 2. Nature 1999, 401, 610–613. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Yoshida, Y.; Hasunuma, K. Photomorphogenetic characteristics are severely affected in nucleoside diphosphate kinase-1 (NDK-1)-disrupted mutants in Neurospora crassa. Mol. Genet. Genomics 2006, 275, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.; Lee, B.; Choi, G.; Shin, D.; Prasad, D.T.; Lee, O.; Kwak, S.-S.; Kim, D.H.; Nam, J.; Bahk, J. NDPkinase 2 interacts with two oxidative stress-activated MAPKs to regulate cellular redox state and enhances multiple stress tolerance in transgenic plants. Proc. Natl. Acad. Sci. USA 2003, 100, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Seong, E.S.; Guo, J.; Kim, Y.H.; Cho, J.H.; Lim, C.K.; Hyun Hur, J.; Wang, M.H. Regulations of marker genes involved in biotic and abiotic stress by overexpression of the AtNDPK2 gene in rice. Biochem. Biophys. Res. Commun. 2007, 363, 126–132. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Liu, D.; Shen, H.; Li, Y.; Nie, Y. Proteome Analysis of Dormancy-Released Seeds of Fraxinus mandshurica Rupr. in Response to Re-Dehydration under Different Conditions. Int. J. Mol. Sci. 2015, 16, 4713-4730. https://doi.org/10.3390/ijms16034713
Zhang P, Liu D, Shen H, Li Y, Nie Y. Proteome Analysis of Dormancy-Released Seeds of Fraxinus mandshurica Rupr. in Response to Re-Dehydration under Different Conditions. International Journal of Molecular Sciences. 2015; 16(3):4713-4730. https://doi.org/10.3390/ijms16034713
Chicago/Turabian StyleZhang, Peng, Di Liu, Hailong Shen, Yuhua Li, and Yuzhe Nie. 2015. "Proteome Analysis of Dormancy-Released Seeds of Fraxinus mandshurica Rupr. in Response to Re-Dehydration under Different Conditions" International Journal of Molecular Sciences 16, no. 3: 4713-4730. https://doi.org/10.3390/ijms16034713