
Resistance Training Regulates Cardiac Function through Modulation of miRNA-214

Abstract
:1. Introduction
2. Results
2.1. Body Mass and Left Ventricular Mass

| Parameters | CO | TR |
|---|---|---|
| BW (g) | 348.9 ± 6.7 | 341.7 ± 8.5 |
| LVW (mg) | 632.1 ± 30.9 | 753.5 ± 35.4 |
| LV/BW (mg/g) | 1.8 ± 0.06 | 2.2 ± 0.04 * |
| 1RM Initial (g) | 639.2 ± 35.2 | 643.7 ± 41.4 |
| 1RM Final (g) | 1070.4 ± 41.7 | 2168.2 ± 37.8 * |
2.2. Maximal Strength
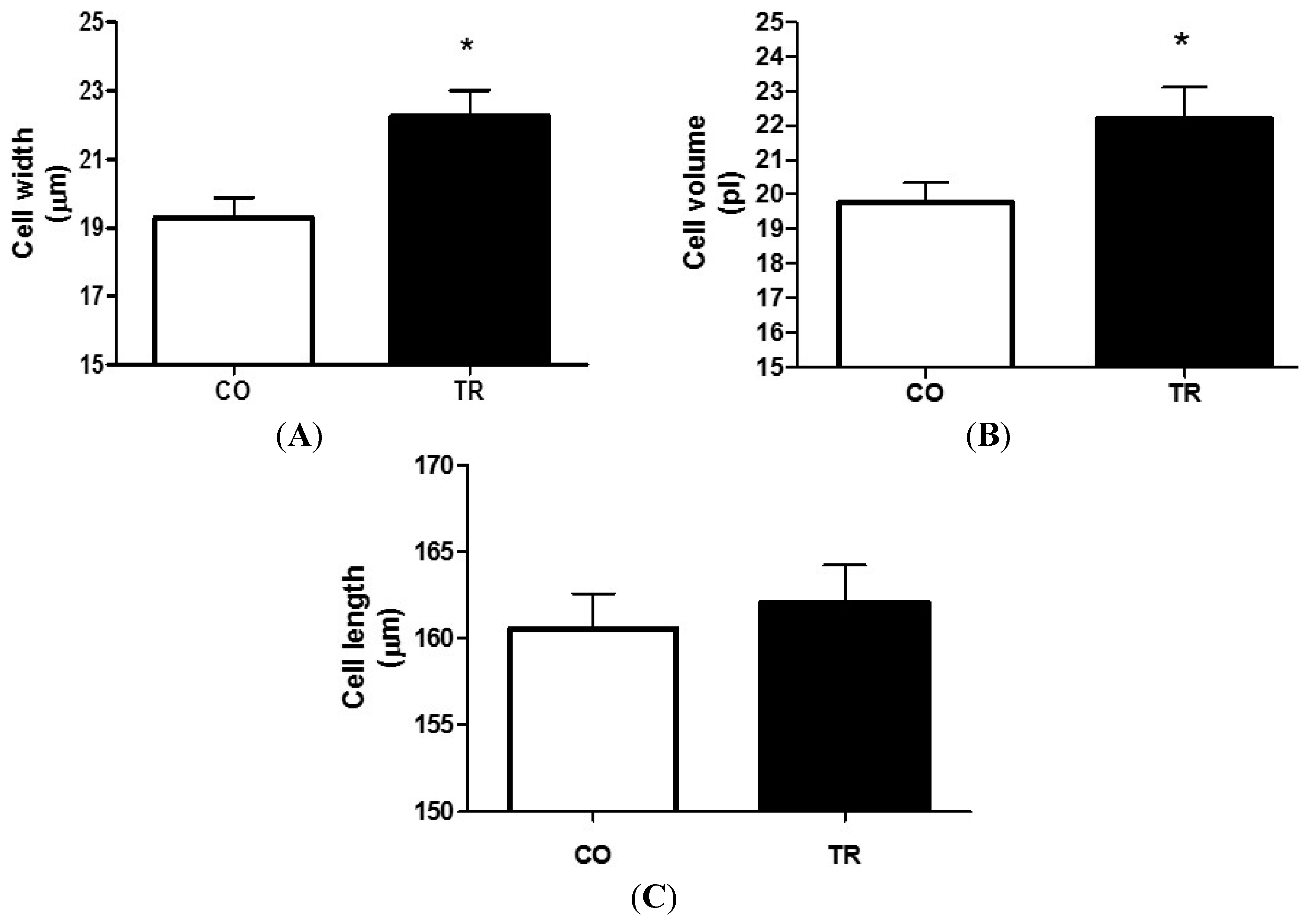
2.3. Effects of Training on Cardiomyocyte Dimensions
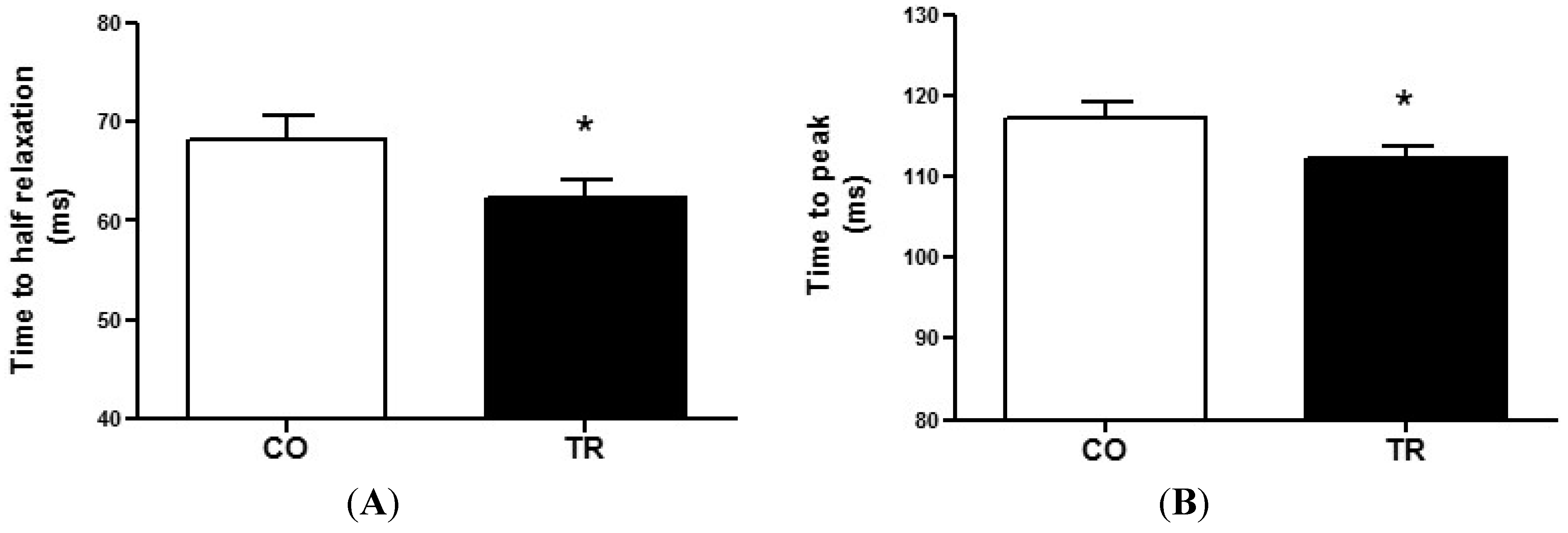
2.4. Cell Contractility
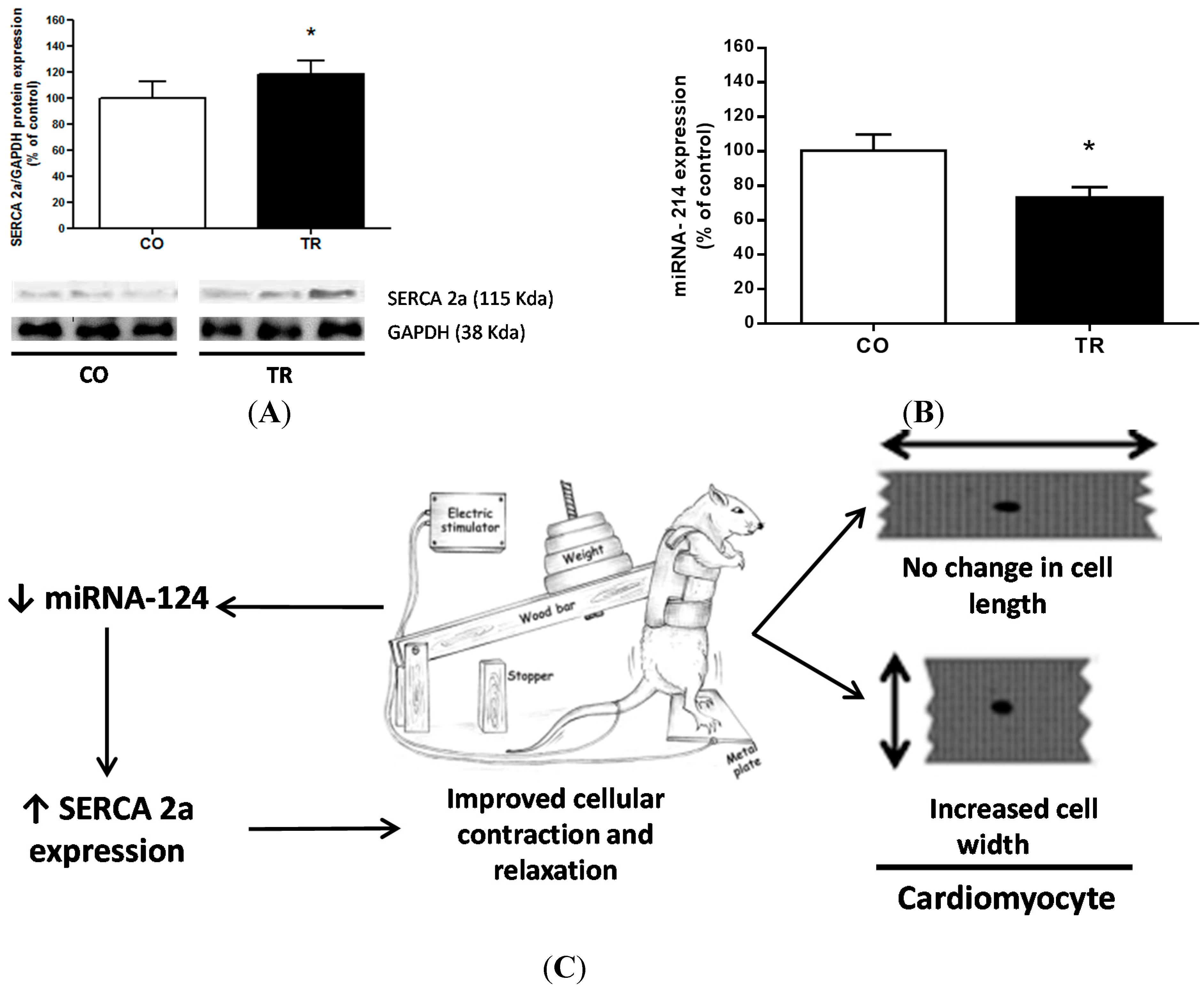
2.5. Protein and miRNA Expression



3. Discussion
4. Methods
4.1. Animals
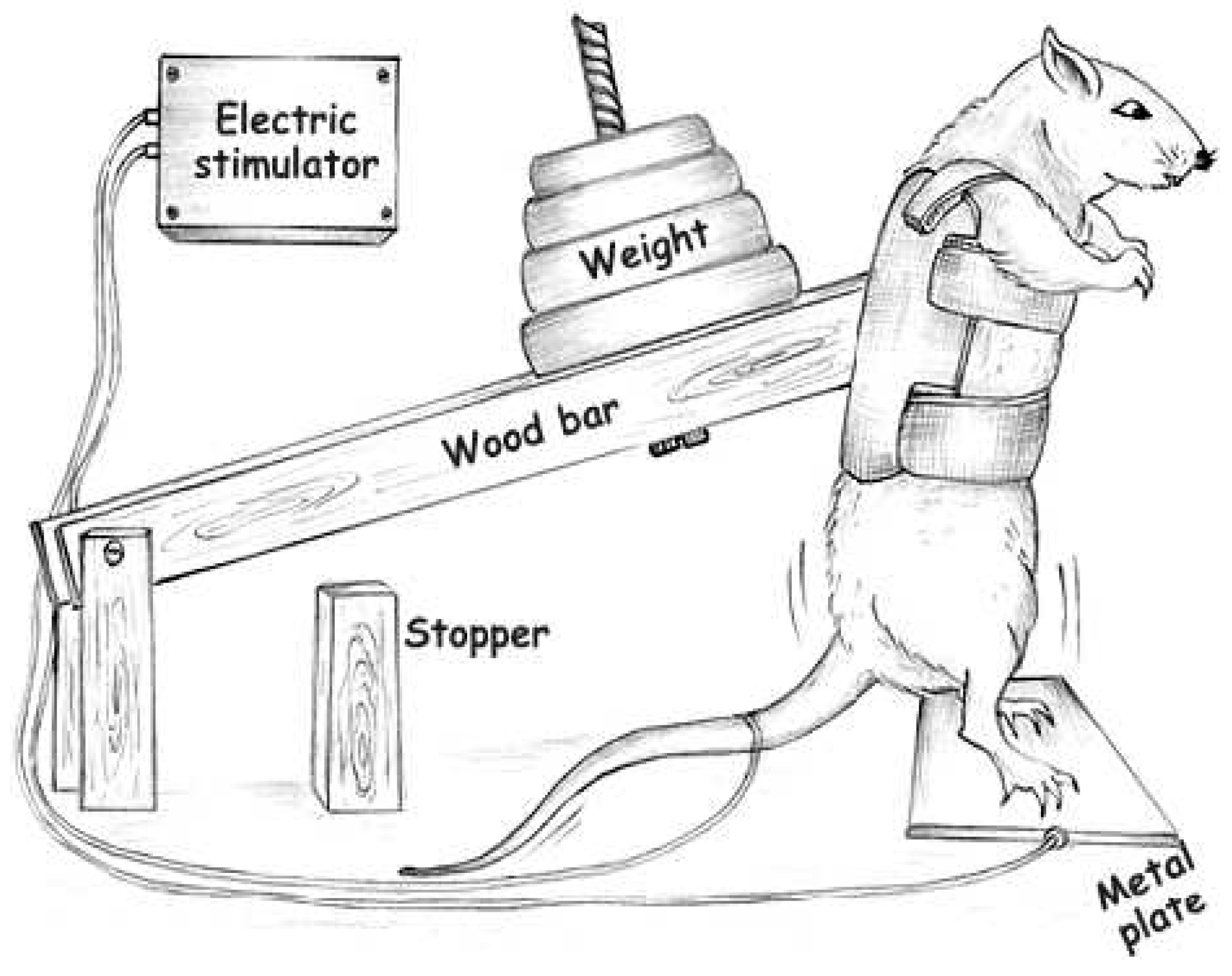
4.2. Exercise Training Protocol
4.3. Cardiomyocytes Isolation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
4.4. Measurements of Cell Contractility and Morphology
4.5. Western Blot Analysis
4.6. miRNA Quantification Using Real-Time PCR
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Alves, J.P.; Nunes, R.B.; Stefani, G.P.; dal Lago, P. Resistance training improves hemodynamic function, collagen deposition and inflammatory profiles: Experimental model of heart failure. PLoS ONE 2014, 9, e110317. [Google Scholar] [CrossRef] [PubMed]
- Braith, R.W.; Beck, D.T. Resistance exercise: Training adaptations and developing a safe exercise prescription. Heart Fail. Rev. 2008, 13, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Drummond, L.R.; Carlo, R.J.; Melo, S.F.S.; Carneiro-Junior, M.A.; Silva, K.A.; Rodrigues, A.C.; Soares, P.N.P.; Gomes, T.N.P.; Lousada, M.J.Q.; Oliveira, E.M.; et al. Enhanced femoral neck strength in response to weightlifting exercise training in maturing male rats. Int. Sport Med. J. 2013, 14, 155–167. [Google Scholar]
- Barauna, V.G.; Magalhaes, F.C.; Krieger, J.E.; Oliveira, E.M. AT1 receptor participates in the cardiac hypertrophy induced by resistance training in rats. Am. J. Physiol. 2008, 295, R381–R387. [Google Scholar]
- Pinter, R.D.E.; Padilha, A.S.; Oliveira, E.M.; Vassallo, D.V.; Fucio, L.J.H. Cardiovascular adaptive responses in rats submitted to moderate resistance training. Eur. J. Appl. Physiol. 2008, 103, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Naylor, L.H.; George, K.; O’Driscoll, G.; Green, D.J. The athlete’s heart: A contemporary appraisal of the “Morganroth hypothesis”. Sports Med. 2008, 38, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, B.C.; Weeks, K.L.; Pretorius, L.; McMullen, J.R. Molecular distinction between physiological and pathological cardiac hypertrophy: Experimental findings and therapeutic strategies. Pharmacol. Ther. 2010, 128, 191–227. [Google Scholar] [CrossRef] [PubMed]
- Melo, S.F.S.; Lunz, W.; Fontes, E.P.B.; Dias, C.M.G.C.; Júnior, M.A.C.; de Moura, A.G.; del Carlo, R.J.; Natali, A.J. Different levels of Hsp72 in female rat myocardium in response to voluntary exercise and forced exercise. Arq. Bras. Cardiol. 2009, 93, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Colan, S.D.; Sanders, S.P.; Borow, K.M. Physiologic hypertrophy: Effects on left ventricular systolic mechanics in athletes. J. Am. Coll. Cardiol. 1987, 9, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Hossack, K.F. Cardiovascular responses to dynamic exercise. Cardiol. Clin. 1987, 5, 147–156. [Google Scholar] [PubMed]
- Fagard, R.H. Athlete’s heart: A meta-analysis of the echocardiographic experience. Int. J. Sports Med. 1996, 17, 140–144. [Google Scholar] [CrossRef] [PubMed]
- White, F.C.; Witzel, G.; Breisch, E.A.; Bloor, C.M.; Nimmo, L.E. Regional capillary and myocyte distribution in normal and exercise trained male and female rat hearts. Am. J. Cardiovasc. Pathol. 1988, 2, 247–253. [Google Scholar] [PubMed]
- Laughlin, M.H.; Schaefer, M.E.; Sturek, M. Effect of exercise training on intracellular free Ca2+ transients in ventricular myocytes of rats. J. Appl. Physiol. 1992, 73, 1441–1448. [Google Scholar] [PubMed]
- Natali, A.J.; Turner, D.L.; Harrison, S.M.; White, E. Regional effects of voluntary exercise on cell size and contraction-frequency responses in rat cardiac myocytes. J. Exp. Biol. 2001, 204, 1191–1199. [Google Scholar] [PubMed]
- Wisløff, U.; Loennechen, J.P.; Falck, G.; Beisvag, V.; Currie, S.; Smith, G.; Ellingsen, O. Increased contractility and calcium sensitivity in cardiac myocytes isolated from endurance trained rats. Cardiovasc. Res. 2001, 50, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Primola-Gomes, T.N.; Campos, L.A.; Lauton-Santos, S.; Balthazar, C.H.; Guatimosim, S.; Capettini, L.S.; Lemos, V.S.; Coimbra, C.C.; Soares, D.D.; Carneiro-Júnior, M.A.; et al. Exercise capacity is related to calcium transients in ventricular cardiomyocytes. J. Appl. Physiol. 2009, 107, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Grossman, W.; Jones, D.; McLaurin, L.P. Wall stress and patterns of hypertrophy in the human left ventricle. J. Clin. Investig. 1975, 56, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Longhurst, J.C.; Kelly, A.R.; Gonyea, W.J.; Mitchell, J.H. Echocardiographic left ventricular masses in distance runners and weight lifters. J. Appl. Physiol. 1980, 48, 154–162. [Google Scholar] [PubMed]
- Barauna, V.G.; Batista, M.L., Jr.; Costa Rosa, L.F.; Casarini, D.E.; Krieger, J.E.; Oliveira, E.M. Cardiovascular adaptations in rats submitted to a resistance-training model. Clin. Exp. Pharmacol. Physiol. 2005, 32, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.L.; Korzick, D.H. Cellular adaptations of the myocardium to chronic exercise. Prog. Cardiovasc. Dis. 1995, 37, 371–396. [Google Scholar] [CrossRef] [PubMed]
- Natali, A.J.; Wilson, L.A.; Peckham, M.; Turner, D.L.; Harrison, S.M.; White, E. Different regional effects of voluntary exercise on the mechanical and electrical properties of rat ventricular myocytes. J. Physiol. 2002, 541, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Kemi, O.J.; MacQuaide, N.; Hoydal, M.A.; Ellingsen, O.; Smith, G.L.; Wisloff, U. Exercise training corrects control of spontaneous calcium waves in hearts from myocardial infarction heart failure rats. J. Cell. Physiol. 2012, 227, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Soci, U.P.; Fernandes, T.; Hashimoto, N.Y.; Mota, G.F.; Amadeu, M.A.; Rosa, K.T.; Irigoyen, M.C.; Phillips, M.I.; Oliveira, E.M. MicroRNAs 29 are involved in the improvement of ventricular compliance promoted by aerobic exercise training in rats. Physiol. Genomics 2011, 43, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Das Neves, V.J.; Fernandes, T.; Redondo, F.R.R.; Soci, U.P.R.; Melo, S.F.S.; de Oliveira, E.M. Exercise training in hypertension: Role of microRNAs. World J. Cardiol. 2014, 6, 713–727. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, T.; Magalhães, F.C.; Roque, F.R.; Phillips, M.I.; Oliveira, E.M. Exercise training prevents the microvascular rarefaction in hypertension balancing angiogenic and apoptotic factors: Role of microRNAs-16, -21, and -126. Hypertension 2012, 59, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Melo, S.T.F.; Fernandes, T.; Baraúna, V.G.; Matos, K.C.; Santos, A.A.; Tucci, P.J.F.; Oliveira, E.M. Expression of microRNA-29 and collagen in cardiac muscle after swimming training in myocardial-infarcted rats. Cell. Physiol. Biochem. 2014, 33, 657–669. [Google Scholar] [CrossRef] [PubMed]
- Dickerman, R.D.; Schaller, F.; McConathy, W.J. Left ventricular wall thickening does occur in elite power athletes with or without anabolic steroid use. Cardiology 1998, 90, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Barauna, V.G.; Rosa, K.T.; Irigoyen, M.C.; Oliveira, E.M. Effects of resistance training on ventricular function and hypertrophy in a rat model. Clin. Med. Res. 2007, 5, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Mokelke, E.A.; Palmer, B.M.; Cheung, J.Y.; Moore, R.L. Endurance training does not affect intrinsic calcium current characteristics in rat myocardium. Am. J. Physiol. 1997, 273, H1193–H1197. [Google Scholar] [PubMed]
- Palmer, B.M.; Thayer, A.M.; Snyder, S.M.; Moore, R.L. Shortening and [Ca2+] dynamics of left ventricular myocytes isolated from exercise-trained rats. J. Appl. Physiol. 1998, 85, 2159–2168. [Google Scholar] [PubMed]
- Diffee, G.M.; Nagle, D.F. Exercise training alters length dependence of contractile properties in rat myocardium. J. Appl. Physiol. 2003, 94, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Diffee, G.M.; Seversen, E.A.; Titus, M.M. Exercise training increases the Ca2+ sensitivity of tension in rat cardiac myocytes. J. Appl. Physiol. 2001, 91, 309–315. [Google Scholar] [PubMed]
- Pluim, B.M.; Zwinderman, A.H.; van der Laarse, A.; van der Wall, E.E. The athlete’s heart. A meta-analysis of cardiac structure and function. Circulation 2000, 101, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Harada, M.; Luo, X.; Murohara, T.; Yang, B.; Dobrev, D.; Nattel, S. MicroRNA regulation and cardiac calcium signaling: Role in cardiac disease and therapeutic potential. Circ. Res. 2014, 114, 689–705. [Google Scholar] [CrossRef] [PubMed]
- MacDougall, J.D.; Tuxen, D.; Sale, D.G.; Moroz, J.R.; Sutton, J.R. Arterial blood pressure response to heavy resistance exercise. J. Appl. Physiol. 1985, 58, 785–790. [Google Scholar] [PubMed]
- Tibbits, G.F.; Barnard, R.J.; Baldwin, K.M.; Cugalj, N.; Roberts, N.K. Influence of exercise on excitation-contraction coupling in rat myocardium. Am. J. Physiol. 1981, 240, 472–480. [Google Scholar]
- Bers, D.M. Cardiac excitation-contraction coupling. Nature 2002, 415, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Sah, R.; Ramirez, R.J.; Oudit, G.Y.; Gidrewicz, D.; Trivieri, M.G.; Zobel, C.; Backx, P.H. Regulation of cardiac excitation-contraction coupling by action potential repolarization: Role of the transient outward potassium current (Ito). J. Physiol. 2003, 546, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.L.; Hashimoto, K.; Grupp, I.L.; Ji, Y.; Reed, T.; Loukianov, E.; Grupp, G.; Bhagwhat, A.; Hoit, B.; Walsh, R.; et al. Targeted overexpression of the sarcoplasmic reticulum Ca2+-ATPase increases cardiac contractility in transgenic mouse hearts. Circ. Res. 1998, 83, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, M.; Bhupathy, P.; Babu, G.J. Regulation of sarcoplasmic reticulum Ca2+ ATPase pump expression and its relevance to cardiac muscle physiology and pathology. Cardiovasc. Res. 2008, 77, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, M.; Huke, S. SERCA pump level is a critical determinant of Ca2+ homeostasis and cardiac contractility. J. Mol. Cell. Cardiol. 2001, 33, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Vetter, R.; Rehfeld, U.; Reissfelder, C.; Weiss, W.; Wagner, K.D.; Günther, J.; Hammes, A.; Tschöpe, C.; Dillmann, W.; Paul, M. Transgenic overexpression of the sarcoplasmic reticulum Ca2+ATPase improves reticular Ca2+ handling in normal and diabetic rat hearts. FASEB J. 2002, 16, 1657–1659. [Google Scholar] [PubMed]
- Kemi, O.J.; Ellingsen, Ø.; Ceci, M.; Grimaldi, S.; Smith, G.L.; Condorelli, G.; Wisløff, U. Aerobic interval training enhances cardiomyocyte contractility and Ca2+ cycling by phosphorylation of CaMKII and Thr-17 of phospholamban. J. Mol. Cell. Cardiol. 2007, 43, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Van Rooij, E.; Olson, E.N. MicroRNA therapeutics for cardiovascular disease: Opportunities and obstacles. Nat. Rev. Drug. Discov. 2012, 11, 860–872. [Google Scholar] [CrossRef] [PubMed]
- Gurha, P.; Abreu-Goodger, C.; Wang, T.; Ramirez, M.O.; Drumond, A.L.; van Dongen, S.; Chen, Y.; Bartonicek, N.; Enright, A.J.; Lee, B.; et al. Targeted deletion of microRNA-22 promotes stress-induced cardiac dilation and contractile dysfunction. Circulation 2012, 125, 2751–2761. [Google Scholar] [CrossRef] [PubMed]
- Wahlquist, C.; Jeong, D.; Rojas-Muñoz, A.; Kho, C.; Lee, A.; Mitsuyama, S.; van Mil, A.; Park, W.J.; Sluijter, J.P.; Doevendans, P.A.; et al. Inhibition of miR-25 improves cardiac contractility in the failing heart. Nature 2014, 508, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Aurora, A.B; Mahmoud, A.I.; Luo, X.; Johnson, B.A.; van Rooij, E.; Matsuzaki, S.; Humphries, K.M.; Hill, J.A.; Bassel-Duby, R.; Sadek, H.A.; et al. MicroRNA-214 protects the mouse heart from ischemic injury by controlling Ca2+ overload and cell death. J. Clin. Investig. 2012, 122, 1222–1232. [Google Scholar] [CrossRef] [PubMed]
- Morissette, M.P.; Susser, S.E.; Stammers, A.N.; O’Hara, K.A.; Gardiner, P.F.; Sheppard, P.; Moffatt, T.L.; Duhamel, T.A. Differential regulation of the fibert ype-specific gene expression of the sarcoplasmic reticulum calcium-ATPase isoforms induced by exercise training. J. Appl. Physiol. 2014, 117, 544–555. [Google Scholar] [CrossRef] [PubMed]
- Safwat, Y.; Yassin, N.; Gamal El Din, M.; Kassem, L. Modulation of skeletal muscle performance and SERCA by exercise and adiponectin gene therapy in insulin-resistant rat. DNA Cell Biol. 2013, 32, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, T.; Uchiyama, S.; Nakano, S. A weight-lifting exercise model for inducing hypertrophy in the hindlimb muscles of rats. Med. Sci. Sports Exerc. 1992, 24, 881–886. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melo, S.F.S.; Barauna, V.G.; Júnior, M.A.C.; Bozi, L.H.M.; Drummond, L.R.; Natali, A.J.; De Oliveira, E.M. Resistance Training Regulates Cardiac Function through Modulation of miRNA-214. Int. J. Mol. Sci. 2015, 16, 6855-6867. https://doi.org/10.3390/ijms16046855
Melo SFS, Barauna VG, Júnior MAC, Bozi LHM, Drummond LR, Natali AJ, De Oliveira EM. Resistance Training Regulates Cardiac Function through Modulation of miRNA-214. International Journal of Molecular Sciences. 2015; 16(4):6855-6867. https://doi.org/10.3390/ijms16046855
Chicago/Turabian StyleMelo, Stéphano Freitas Soares, Valério Garrone Barauna, Miguel Araújo Carneiro Júnior, Luiz Henrique Marchesi Bozi, Lucas Rios Drummond, Antônio José Natali, and Edilamar Menezes De Oliveira. 2015. "Resistance Training Regulates Cardiac Function through Modulation of miRNA-214" International Journal of Molecular Sciences 16, no. 4: 6855-6867. https://doi.org/10.3390/ijms16046855