
Cellular Anti-Melanogenic Effects of a Euryale ferox Seed Extract Ethyl Acetate Fraction via the Lysosomal Degradation Machinery



Abstract
:
1. Introduction
2. Results
2.1. In Vitro Antioxidant Activities of E. ferox Seed Ethyl Acetate Fraction (Efse-EA)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Concentration (µg/mL) | DPPH (1) | FRAP (2) | CUPRAC (3) | ABTS (4) |
|---|---|---|---|---|---|
| Activity (% of Control) | |||||
| Ascorbic acid | 1 | 4.5 ± 0.011 (5) | 12.7 ± 0.002 ** | 12.2 ± 0.001 | −1.7 ± 0.002 |
| 3 | 8.5 ± 0.016 **,(6) | 52.5 ± 0.001 ** | 42.7 ± 0.001 ** | 2.7 ± 0.001 ** | |
| 10 | 18.5 ± 0.054 * | 170.8 ± 0.006 ** | 147.8 ± 0.001 ** | 18.1 ± 0.003 ** | |
| 30 | 57.3 ± 0.069 ** | 523.3 ± 0.026 ** | 119.2 ± 0.002 ** | 57.9 ± 0.008 ** | |
| Efse-EA | 1 | 0.6 ± 0.002 | 19.9 ± 0.003 ** | 12.8 ± 0.003 * | 5.2 ± 0.001 ** |
| 3 | 7.9 ± 0.012 | 82.0 ± 0.009 ** | 54.3 ± 0.009 ** | 18.6 ± 0.013 * | |
| 10 | 28.1 ± 0.006 ** | 234.4 ± 0.027 ** | 251.9 ± 0.029 ** | 55.1 ± 0.024 * | |
| 30 | 78.0 ± 0.021 ** | 820.4 ± 0.065 ** | 669.6 ± 0.043 ** | 89.7 ± 0.001 * | |

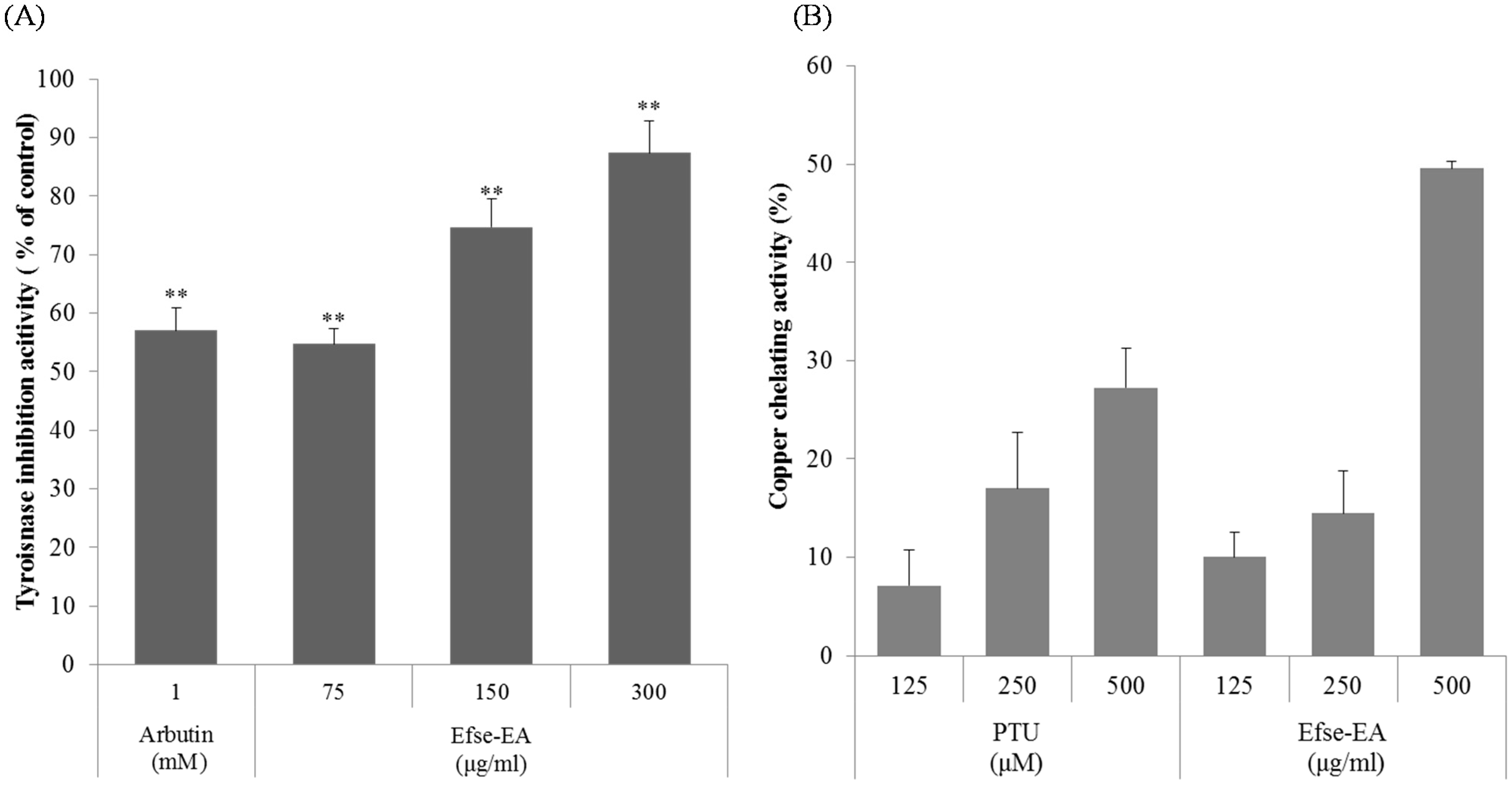
2.2. Effect of Efse-EA on Mushroom Tyrosinase and Copper Chelation

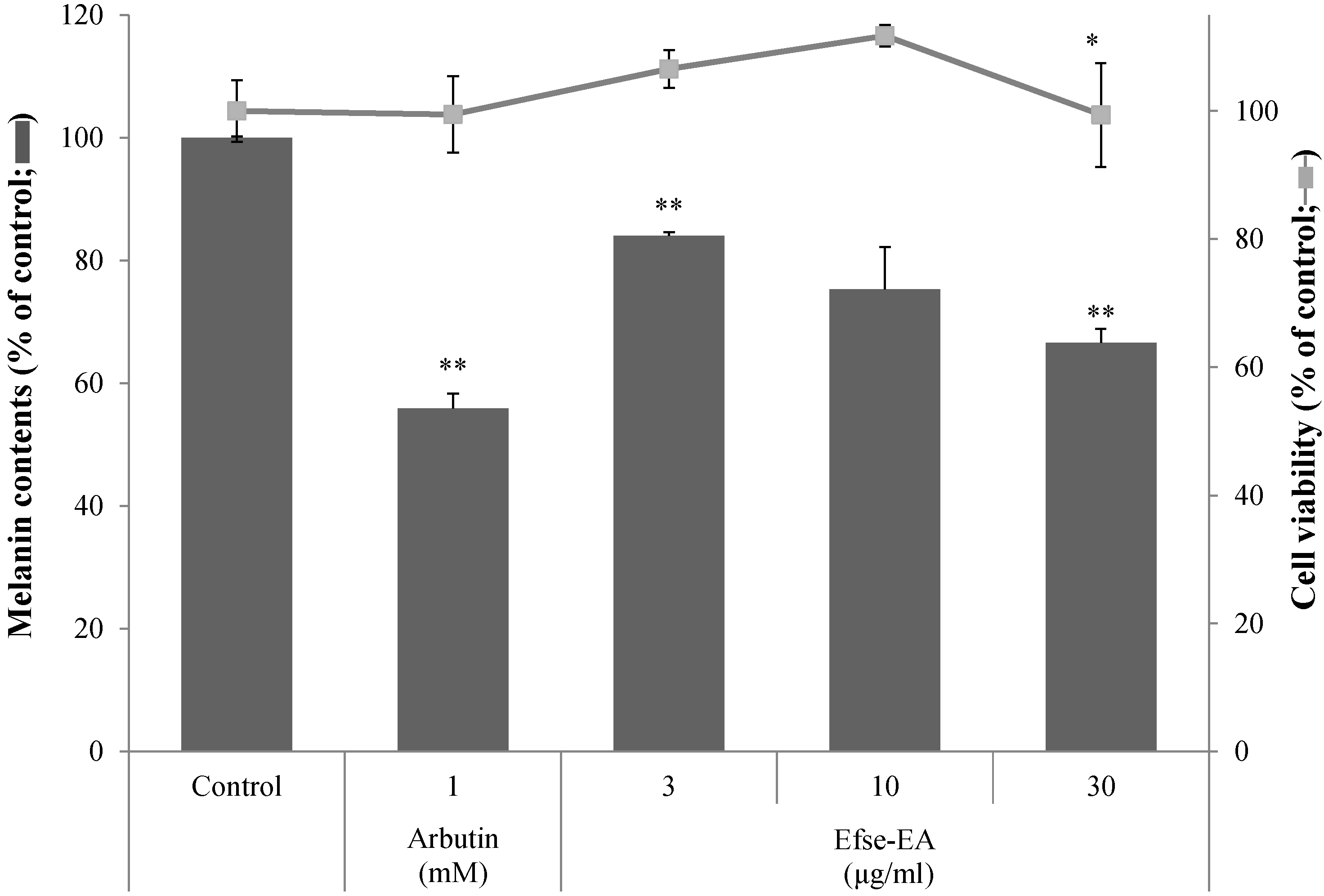
2.3. Effect of Efse-EA on Cell Viability and Melanin Levels

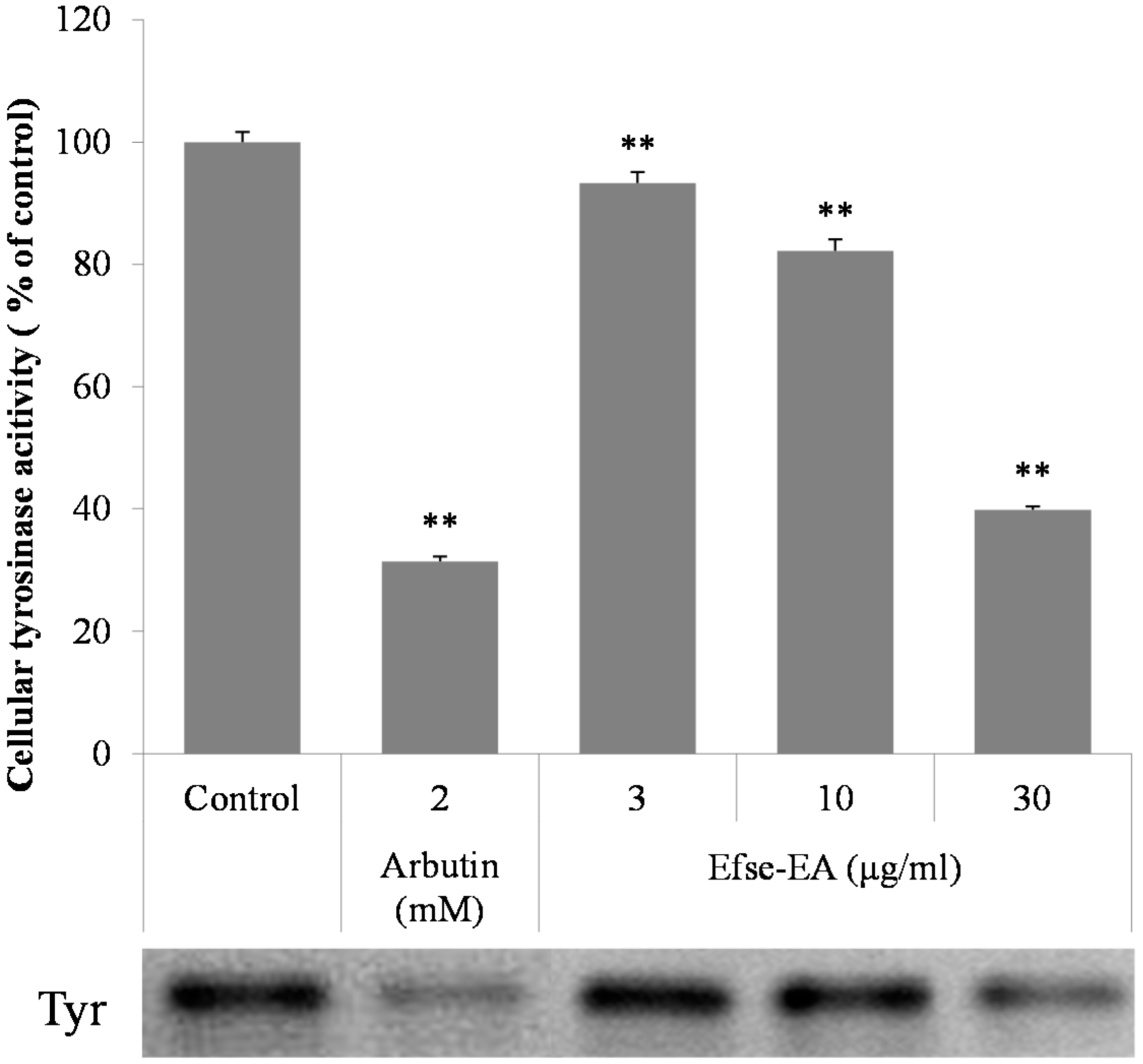
2.4. Effects of Efse-EA on l-DOPA (l-3,4-Dihydroxyphenylalanine) Zymography and on Cellular Tyrosinase Activity

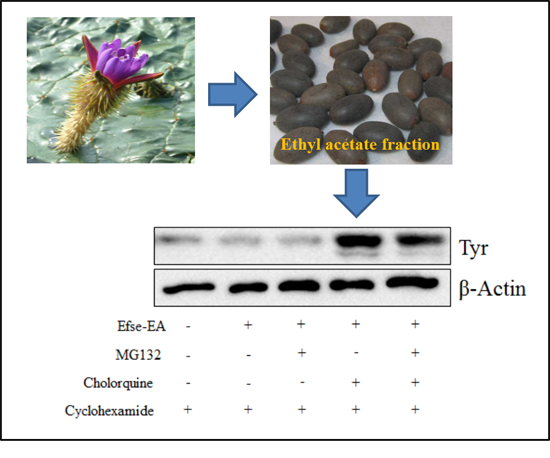
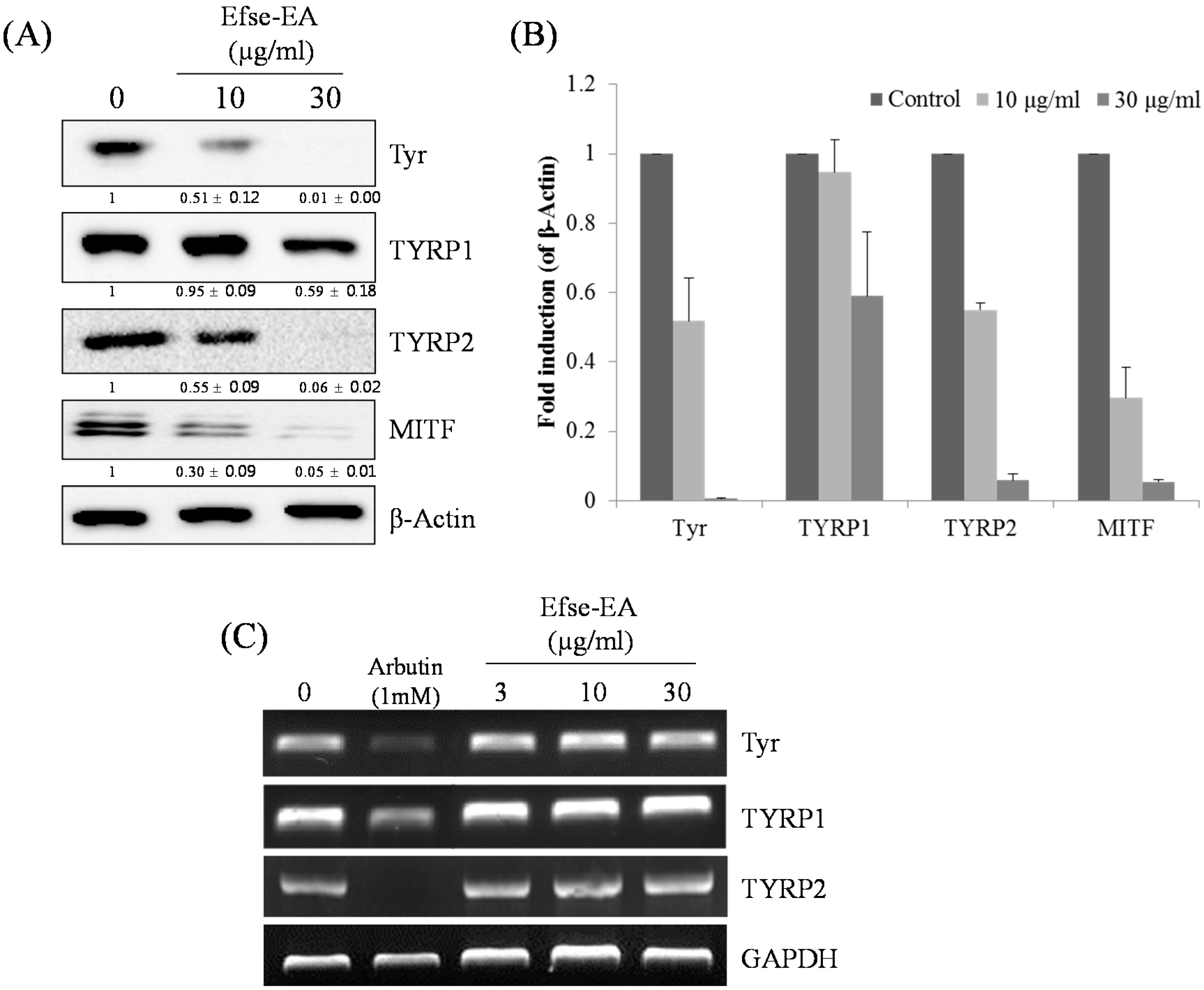
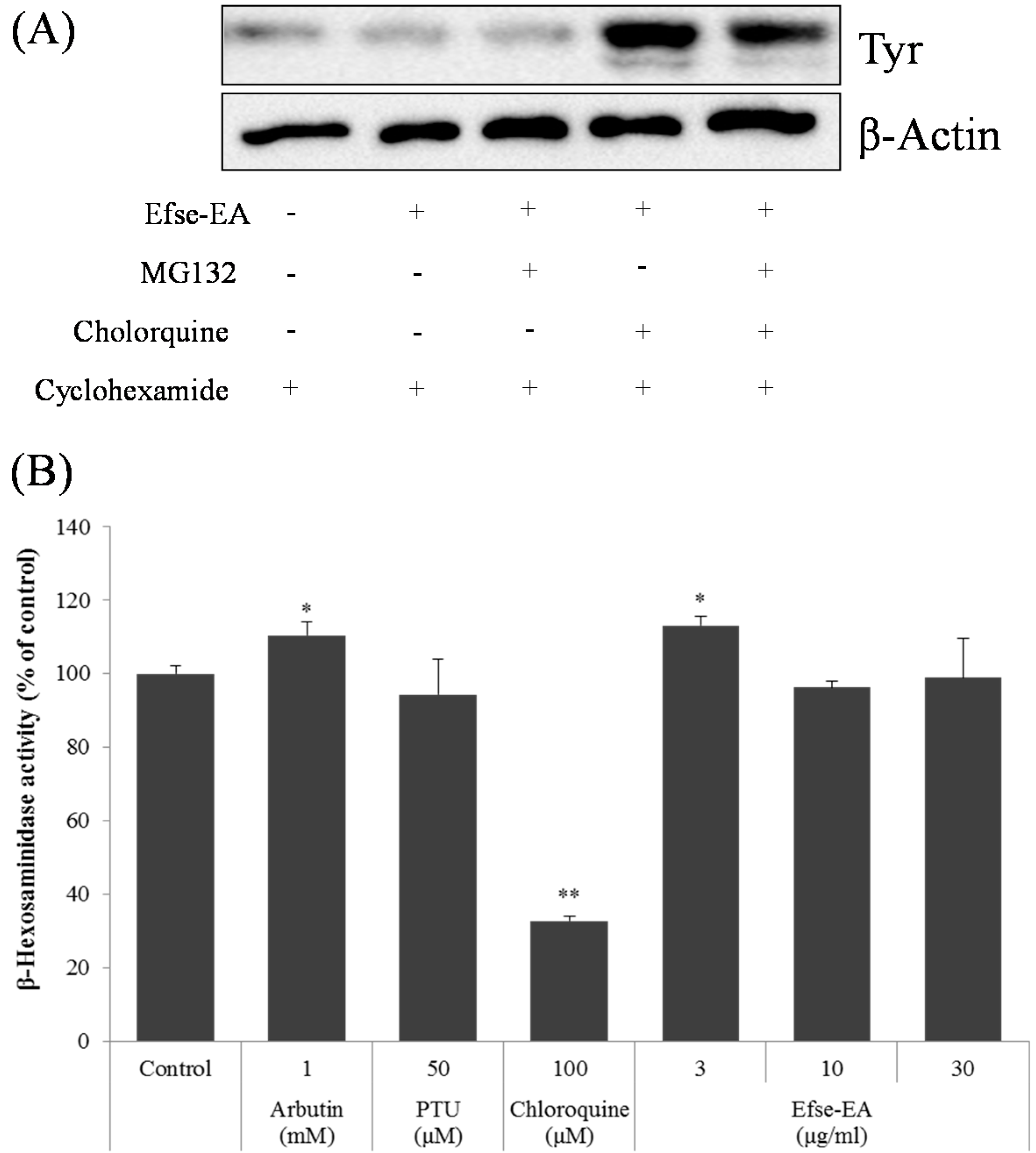
2.5. Effect of Efse-EA on Proteins Involved in Melanogenesis and Lysosomal Degradation


3. Discussion
4. Experimental Section
4.1. Materials
4.2. Cell Cultures
4.3. Preparation of Plant Extracts
4.4. Cell Viability Assay
4.5. Evaluation of in Vitro Antioxidant Activities
4.6. Measurement of Tyrosinase and Copper Chelating Activity
4.7. Melanogenesis Inhibition Assay in Melan-a Cells
4.8. Analysis of Intracellular Tyrosinase by Zymography
4.9. Reverse-Transcription Polymerase Chain Reaction (RT-PCR) Analysis
4.10. Western Blot Analysis
4.11. β-Hexosaminidase Assay
4.12. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Song, C.W.; Wang, S.M.; Zhou, L.L.; Hou, F.F.; Wang, K.J.; Han, Q.B.; Li, N.; Cheng, Y.X. Isolation and identification of compounds responsible for antioxidant capacity of Euryale ferox seeds. J. Agric. Food Chem. 2011, 59, 1199–1204. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Lee, M.J.; Lee, H.S.; Kim, J.G.; Park, W.H. Screening of antioxidative effect and suppressive effect of LDL oxidation of Euryale ferox Salisbury. Korean J. Orient. Physiol. Pathol. 2011, 25, 92–99. [Google Scholar]
- Das, S.; Der, P.; Raychaudhuri, U.; Maulik, N.; Das, D.K. The effect of Euryale ferox (Makhana), an herb of aquatic origin, on myocardial ischemic reperfusion injury. Mol. Cell. Biochem. 2006, 289, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Luo, J.; Kong, L.Y. Two new tocopherol polymers from the seeds of Euryale ferox. J. Asian Nat. Prod. Res. 2012, 14, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhao, S.; Guillaume, D.; Sun, C. New cerebrosides from Euryale ferox. J. Nat. Prod. 1994, 57, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhao, S.; Sun, C.; Guillaume, D. Glucosylsterols in extracts of Euryale ferox identified by high resolution NMR and mass spectrometry. J. Lipid Res. 1989, 30, 1633–1637. [Google Scholar] [PubMed]
- Row, L.C.; Ho, J.C.; Chen, C.M. Cerebrosides and tocopherol trimers from the seeds of Euryale ferox. J. Nat. Prod. 2007, 70, 1214–1217. [Google Scholar] [CrossRef] [PubMed]
- Choo, S.J.; Kim, Y.H.; Ryoo, I.J.; Xu, G.H.; Yoo, I.D. Application as a cosmeceutical ingredient of Euryale ferox seed extract. J. Soc. Cosmet. Sci. Korea 2009, 35, 309–315. [Google Scholar]
- Puri, A.; Sahai, R.; Singh, K.L.; Saxena, R.P.; Tandon, J.S.; Saxena, K.C. Immunostimulant activity of dry fruits and plant materials used in indian traditional medical system for mothers after child birth and invalids. J. Ethnopharmacol. 2000, 71, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Kim, J.H.; Son, E.S.; Park, H.R. Protective effect of extracts from Euryale ferox against glutamate-induced cytotoxicity in neuronal cells. Nat. Prod. Sci. 2009, 15, 162–166. [Google Scholar]
- Wu, C.; Chen, R.; Wang, X.S.; Shen, B.; Yue, W.; Wu, Q. Antioxidant and anti-fatigue activities of phenolic extract from the seed coat of Euryale ferox Salisb. and identification of three phenolic compounds by LC–ESI-MS/MS. Molecules 2013, 18, 11003–10021. [Google Scholar] [CrossRef] [PubMed]
- Hideya, A.; Hirofumi, K.; Masamitsu, I.; Vincent, J.H. Approaches to identify inhibitors of melanin biosynthesis via the quality control of tyrosinase. J. Investig. Dermatol. 2007, 127, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Hearing, V.J. Determination of melanin synthetic pathways. J. Investig. Dermatol. 2011, 17, E8–E11. [Google Scholar]
- Sato, K.; Takahashi, H.; Iraha, R.; Toriyama, M. Down-regulation of tyrosinase expression by acetylsalicylic acid in murine B16 melanoma. Biol. Pharm. Bull. 2008, 31, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Chao, H.C.; Najjaa, H.; Villareal, M.O.; Ksouri, R.; Han, J.; Neffati, M.; Isoda, H. Arthrophytum scoparium inhibitions melanogenesis through the down-regulation of tyrosinase and melanogenic gene expressions in B16 melanoma cells. Exp. Dermatol. 2013, 22, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Rao, F.; Yuting, Z.; Yiran, G.; Fang, C. Antioxidant and tyrosinase inhibition activities of the ethanol-insoluble fraction of water extract of Sapium sebiferum (L.) Roxb. leaves. South. Afr. J. Bot. 2014, 93, 98–104. [Google Scholar] [CrossRef]
- Chang, T.S. An updated review of tyrosinase inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.N.; Yang, H.M.; Kang, S.M.; Kim, D.; Ahn, G.; Jeon, Y.J. Octaphlorethol A isolated from Ishige foliacea inhibits α-MSH-stimulated induced melanogenesis via ERK pathway in B16F10 melanoma cells. Food Chem. Toxicol. 2013, 59, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Su, T.R.; Lin, J.J.; Tsai, C.C.; Huang, T.K.; Yang, Z.Y.; Wu, M.O.; Zheng, Y.Q.; Su, C.C.; Wu, Y.J. Inhibition of melanogenesis by gallic acid: Possible involvement of the PI3K/Akt, MEK/ERK and Wnt/β-catenin signaling pathways in B16F10 cells. Int. J. Mol. Sci. 2013, 14, 20443–20458. [Google Scholar] [CrossRef] [PubMed]
- Zi, S.X.; Ma, H.J.; Li, Y.; Liu, W.; Yang, Q.Q.; Zhao, G.; Lian, S. Oligomeric proanthocyanidins from grape seeds effectively inhibit ultraviolet-induced melanogenesis of human melanocytes in vitro. Int. J. Mol. Med. 2009, 23, 197–204. [Google Scholar] [PubMed]
- Wrześniok, D.; Beberok, A.; Otręba, M.; Buszman, E. Effect of streptomycin on melanogenesis and antioxidant status in melanocytes. Mol. Cell. Biochem. 2013, 383, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Halvorsen, B.L.; Holte, K.; Myhrstad, M.C.; Barikmo, I.; Hvattum, E.; Remberg, S.F.; Wold, A.B.; Haffner, K.; Baugerød, H.; Andersen, L.F.; et al. A systematic screening of total antioxidants in dietary plants. J. Nutr. 2002, 132, 461–471. [Google Scholar] [PubMed]
- Payne, A.C.; Mazzer, A.; Clarkson, G.J.; Taylor, G. Antioxidant assays—Consistent findings from FRAP and ORAC reveal a negative impact of organic cultivation on antioxidant potential in spinach but not watercress or rocket leaves. Food Sci. Nutr. 2013, 1, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, İ. Antioxidant activity of food constituents: An overview. Arch. Toxicol. 2012, 86, 345–391. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, D.S.; Lee, H.K.; Kwon, S.B.; Lee, S.; Ryoo, I.J.; Kim, W.G.; Yoo, I.D.; Park, K.C. Long-term suppression of tyrosinase by terrein via tyrosinase degradation and its decreased expression. Exp. Dermatol. 2009, 18, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Goh, M.J.; Lee, H.K.; Cheng, L.; Kong, D.Y.; Yeon, J.H.; He, Q.Q.; Cho, J.C.; Na, Y.J. Depigmentation effect of kadsuralignan F on melan-a murine melanocytes and human skin equivalents. Int. J. Mol. Sci. 2013, 14, 1655–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellei, B.; Maresca, V.; Flori, E.; Pitisci, A.; Larue, L.; Picardo, M. p38 regulates pigmentation via proteasomal degradation of tyrosinase. J. Biol. Chem. 2010, 285, 7288–7299. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.B.; Shin, G.H.; Lee, J.S.; Lee, O.H.; Park, I.J.; Cho, J.H. Antioxidant and nitrite scavenging activities of Acanthopanax senticosus extract fermented with different mushroom mycelia. Korean J. Food Sci. Technol. 2014, 46, 205–212. [Google Scholar] [CrossRef]
- Hida, T.; Wakamatsu, K.; Sviderskaya, E.V.; Donkin, A.J.; Montoliu, L.; Lynn, L.M.; Yu, B.; Millhauser, G.L.; Ito, S.; Barsh, G.S.; et al. Agouti protein, mahogunin, and attractin in pheomelanogenesis and melanoblast-like alteration of melanocytes: A cAMP-independent pathway. Pigment Cell Melanoma Res. 2009, 22, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.D.; Lee, W.H.; Roh, E.; Seo, C.S.; Son, J.K.; Lee, S.H.; Hwang, B.Y.; Jung, S.H.; Han, S.B.; Kim, Y. Manassantin A inhibits cAMP-induced melanin production by down-regulating the gene expressions of MITF and tyrosinase in melanocytes. Exp. Dermatol. 2011, 20, 761–763. [Google Scholar] [CrossRef] [PubMed]
- Borovanský, J.; Mommaas, A.M.; Smit, N.P.; Eygendaal, D.; Winder, A.J.; Vermeer, B.J.; Pavel, S. Melanogenesis in transfected fibroblasts induces lysosomal activation. Arch. Dermatol. Res. 1997, 289, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.B.; Nielsen, S.E.; Berg, K. Re-examination and further development of a precise and rapid dye method for measuring cell growth/cell kill. J. Immunol. Methods 1989, 119, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Mori, K.; Hatano, T.; Okumura, T.; Uehara, I.; Komagoe, K.; Fujita, Y.; Okuda, T. Studies on inhibition mechanism of autoxidation by tannins and flavonoids. V. Radical-scavenging effects of tannins and related polyphenols on 1,1-diphenyl-2-picrylhydrzyl radical. Chem. Pharm. Bull. 1989, 37, 1919–1921. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of antioxidant power: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Güçlü, K.; Ozyürek, M.; Karademir, S.E. Novel total antioxidant capacity index for dietary polyphenols and vitamins C and E, using their cupric ion reducing capability in the presence of neocuproine: CUPRAC method. J. Agric. Food Chem. 2004, 52, 7970–7981. [Google Scholar] [CrossRef] [PubMed]
- Adewusi, E.A.; Steenkamp, V. In vitro screening for acetylcholinesterase inhibition and antioxidant activity of medicinal plants from southern Africa. Asian Pac. J. Trop. Med. 2011, 4, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, Y.; Oka, M.; Tsunakawa, M.; Tomita, K.; Hatori, M.; Yamamoto, H.; Kamei, H.; Miyaki, T.; Konishi, M.; Oki, T. Melanostatin, a new melanin synthesis inhibitor. Production, isolation, chemical properties, structure and biological activity. J. Antibiot. 1991, 44, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Bennett, D.C.; Cooper, P.J.; Hart, I.R. A line of non-tumorigenic mouse melanocytes, syngeneic with the B16 melanoma and requiring a tumour promoter for growth. Int. J. Cancer 1987, 39, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Elias, P.M.; Menon, G.; Wetzel, B.K.; Williams, J.W. Evidence that stress to the epidermal barrier influenced the development of pigmentation in humans. Pigment Cell Melanoma Res. 2009, 22, 420–434. [Google Scholar] [CrossRef] [PubMed]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [PubMed]
- Gahl, W.A.; Potterf, B.; Durham-Pierre, D.; Brilliant, M.H.; Hearing, V.J. Melanosomal tyrosine transport in normal and pink-eyed dilution murine melanocytes. Pigment Cell Res. 1995, 8, 229–233. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baek, S.-H.; Nam, I.-J.; Kwak, H.S.; Kim, K.-C.; Lee, S.-H. Cellular Anti-Melanogenic Effects of a Euryale ferox Seed Extract Ethyl Acetate Fraction via the Lysosomal Degradation Machinery. Int. J. Mol. Sci. 2015, 16, 9217-9235. https://doi.org/10.3390/ijms16059217
Baek S-H, Nam I-J, Kwak HS, Kim K-C, Lee S-H. Cellular Anti-Melanogenic Effects of a Euryale ferox Seed Extract Ethyl Acetate Fraction via the Lysosomal Degradation Machinery. International Journal of Molecular Sciences. 2015; 16(5):9217-9235. https://doi.org/10.3390/ijms16059217
Chicago/Turabian StyleBaek, Seung-Hwa, In-Jeong Nam, Hyeong Seob Kwak, Ki-Chan Kim, and Sang-Han Lee. 2015. "Cellular Anti-Melanogenic Effects of a Euryale ferox Seed Extract Ethyl Acetate Fraction via the Lysosomal Degradation Machinery" International Journal of Molecular Sciences 16, no. 5: 9217-9235. https://doi.org/10.3390/ijms16059217