
Atrazine Triggers DNA Damage Response and Induces DNA Double-Strand Breaks in MCF-10A Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
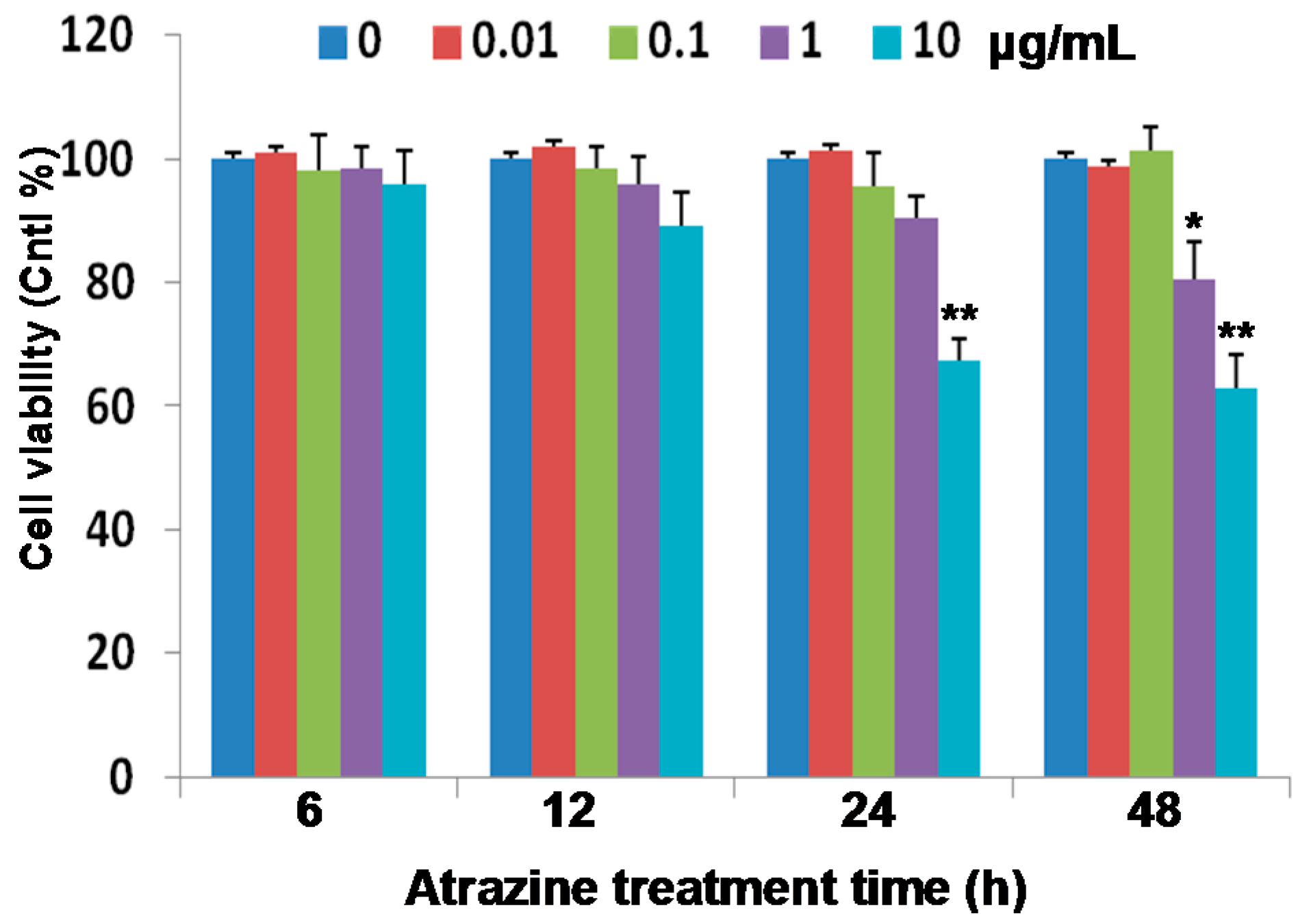
2.1. Reduction of Cell Viability

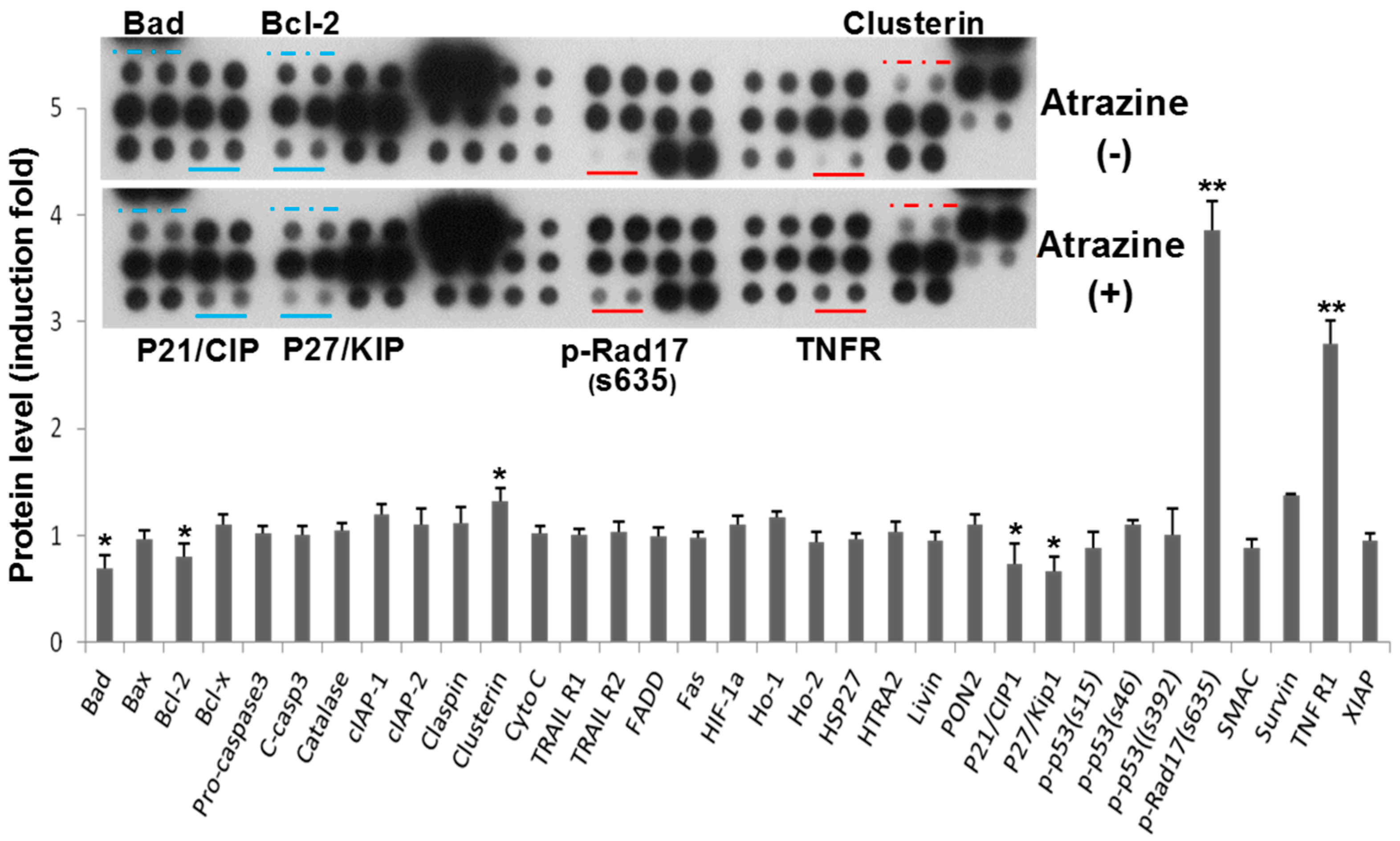
2.2. Regulation of Apoptosis-Related Protein Expression

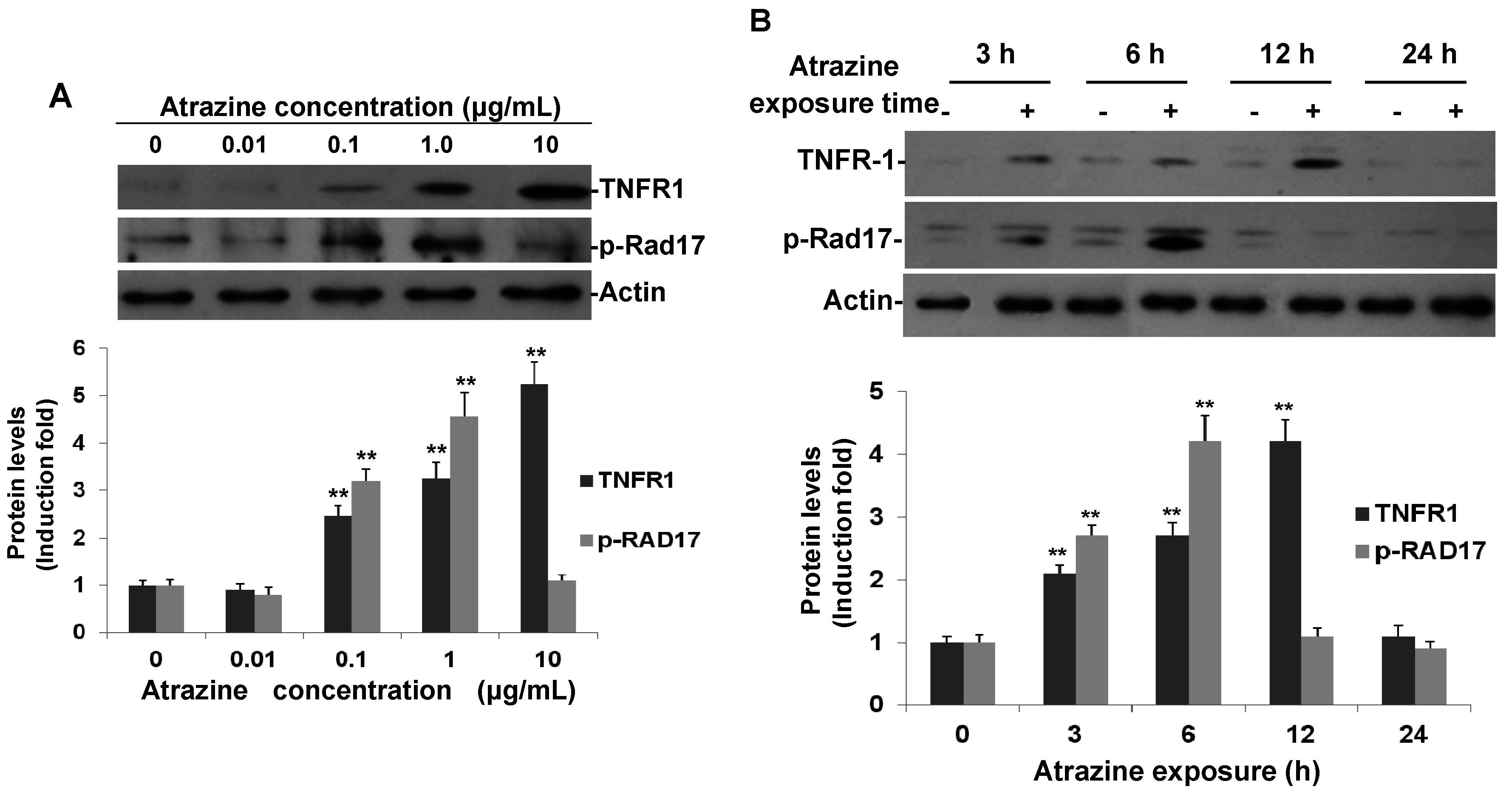
2.3. Confirmation of Phospho-Rad17 (Ser635) and TNFR1

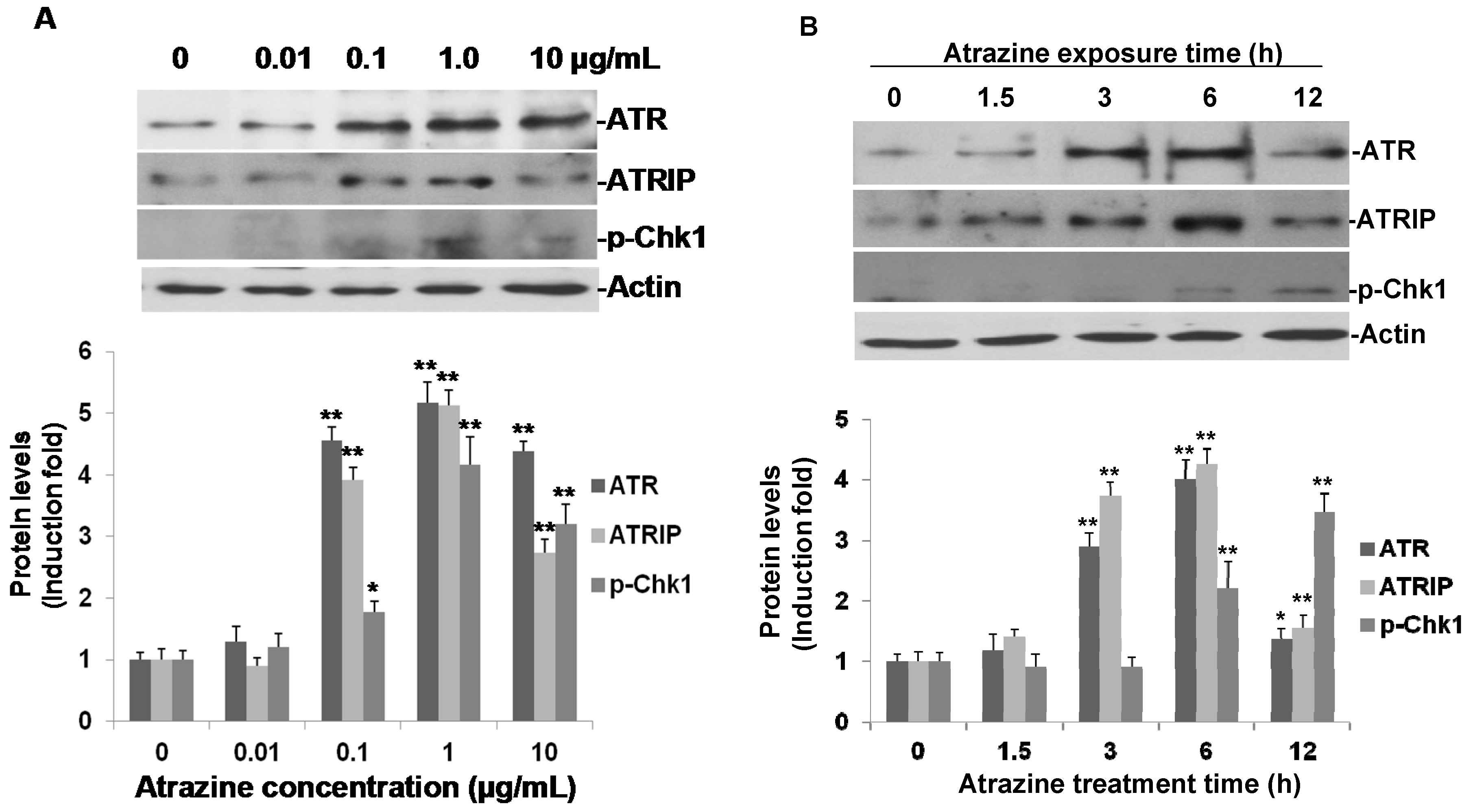
2.4. Activation of ATR-Chk1 Pathway

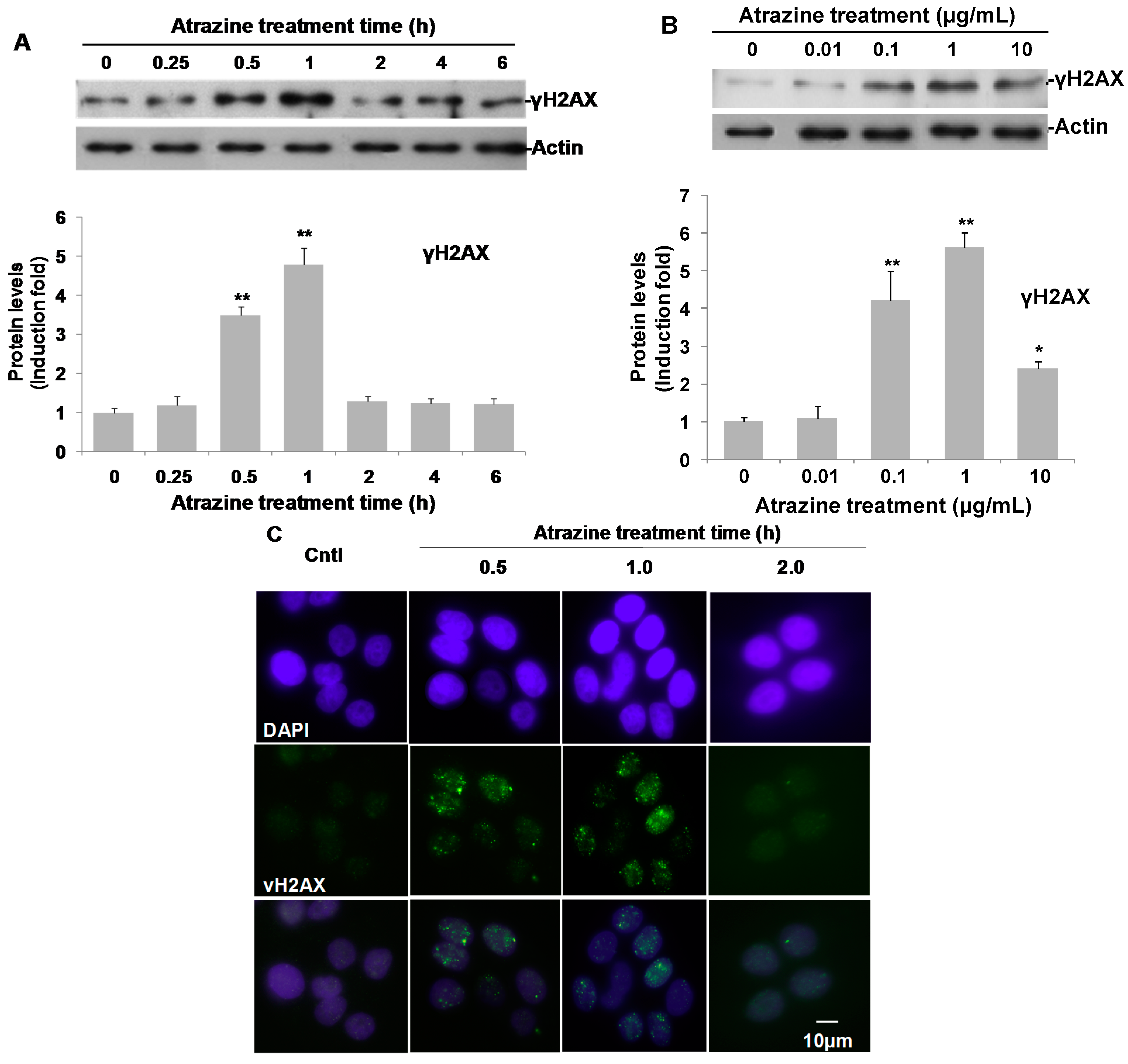
2.5. Induction of γH2AX and γH2AX Foci

3. Experimental Section
3.1. Chemicals and Antibodies
3.2. Cells and Cell Culture
3.3. Cell Proliferation and Toxicity Assay
3.4. Human Apoptosis Antibody Array
3.5. Western Blot Analysis
3.6. Immunofluorescence Imaging of γH2AX Foci
3.7. Statistics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Short, P.; Colborn, T. Pesticide use in the U.S. and policy implications: A focus on herbicides. Toxicol. Ind. Health 1999, 15, 240–275. [Google Scholar] [CrossRef] [PubMed]
- Selim, H.M. Retention and runoff losses of atrazine and metribuzin in soil. J. Environ. Qual. 2003, 32, 1058–1071. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Dong, X.; Xie, H.; Wang, J.; Wang, J.; Su, J.; Yu, C. DNA damage and effects on glutathione-S-transferase activity induced by atrazine exposure in zebrafish (Danio rerio). Environ. Toxicol. 2011, 26, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.L.; Stoker, T.E.; Tyrey, L.; Goldman, J.M.; McElroy, W.K. Atrazine disrupts the hypothalamic control of pituitary-ovarian function. Toxicol. Sci. 2000, 53, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Coban, A.; Filipov, N.M. Dopaminergic toxicity associated with oral exposure to the herbicide atrazine in juvenile male C57BL/6 mice. J. Neurochem. 2007, 100, 1177–1187. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, V.M.; Thiruchelvam, M.; Cory-Slechta, D.A. Sustained exposure to the widely used herbicide atrazine: Altered function and loss of neurons in brain monoamine systems. Environ. Health Perspect. 2005, 113, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Rayner, J.L.; Wood, C.; Fenton, S.E. Exposure parameters necessary for delayed puberty and mammary gland development in Long-Evans rats exposed in utero to atrazine. Toxicol. Appl. Pharmacol. 2004, 195, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Kniewald, J.; Jakominic, M.; Tomljenovic, A.; Simic, B.; Romac, P.; Vranesic, D.; Kniewald, Z. Disorders of male rat reproductive tract under the influence of atrazine. J. Appl. Toxicol. 2000, 20, 61–68. [Google Scholar] [CrossRef]
- Hayes, T.B.; Stuart, A.A.; Mendoza, M.; Collins, A.; Noriega, N.; Vonk, A.; Johnston, G.; Liu, R.; Kpodzo, D. Characterization of atrazine-induced gonadal malformations in African clawed frogs (Xenopus laevis) and comparisons with effects of an androgen antagonist (cyproterone acetate) and exogenous estrogen (17β-estradiol): Support for the demasculinization/feminization hypothesis. Environ. Health Perspect. 2006, 114, 134–141. [Google Scholar] [PubMed]
- Rowe, A.M.; Brundage, K.M.; Barnett, J.B. Developmental immunotoxicity of atrazine in rodents. Basic Clin. Pharmacol. Toxicol. 2008, 102, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Filipov, N.M.; Pinchuk, L.M.; Boyd, B.L.; Crittenden, P.L. Immunotoxic effects of short-term atrazine exposure in young male C57BL/6 mice. Toxicol. Sci. 2005, 86, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Solomon, K.R. Is atrazine a potent endocrine disruptor chemical? Environ. Sci. Technol. Columbia 2009, 43, 2993, author reply 2994. [Google Scholar] [CrossRef]
- Betancourt, M.; Resendiz, A.; Fierro, E.C. Effect of two insecticides and two herbicides on the porcine sperm motility patterns using computer-assisted semen analysis (CASA) in vitro. Reprod. Toxicol. 2006, 22, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Spano, L.; Tyler, C.R.; van Aerle, R.; Devos, P.; Mandiki, S.N.; Silvestre, F.; Thome, J.P.; Kestemont, P. Effects of atrazine on sex steroid dynamics, plasma vitellogenin concentration and gonad development in adult goldfish (Carassius auratus). Aquat. Toxicol. 2004, 66, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Tavera-Mendoza, L.; Ruby, S.; Brousseau, P.; Fournier, M.; Cyr, D.; Marcogliese, D. Response of the amphibian tadpole Xenopus laevis to atrazine during sexual differentiation of the ovary. Environ. Toxicol. Chem. 2002, 21, 1264–1267. [Google Scholar] [CrossRef]
- Tavera-Mendoza, L.; Ruby, S.; Brousseau, P.; Fournier, M.; Cyr, D.; Marcogliese, D. Response of the amphibian tadpole (Xenopus laevis) to atrazine during sexual differentiation of the testis. Environ. Toxicol. Chem. 2002, 21, 527–531. [Google Scholar] [CrossRef]
- Wetzel, L.T.; Luempert, L.G., 3rd.; Breckenridge, C.B.; Tisdel, M.O.; Stevens, J.T.; Thakur, A.K.; Extrom, P.J.; Eldridge, J.C. Chronic effects of atrazine on estrus and mammary tumor formation in female Sprague–Dawley and Fischer 344 rats. J. Toxicol. Environ. Health 1994, 43, 169–182. [Google Scholar] [CrossRef] [PubMed]
- De Roos, A.J.; Zahm, S.H.; Cantor, K.P.; Weisenburger, D.D.; Holmes, F.F.; Burmeister, L.F.; Blair, A. Integrative assessment of multiple pesticides as risk factors for non-Hodgkin’s lymphoma among men. Occup. Environ. Med. 2003, 60, E11. [Google Scholar] [CrossRef] [PubMed]
- Hoar Zahm, S.; Weisenburger, D.D.; Cantor, K.P.; Holmes, F.F.; Blair, A. Role of the herbicide atrazine in the development of non-Hodgkin’s lymphoma. Scand. J. Work Environ. Health 1993, 19, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Donna, A.; Crosignani, P.; Robutti, F.; Betta, P.G.; Bocca, R.; Mariani, N.; Ferrario, F.; Fissi, R.; Berrino, F. Triazine herbicides and ovarian epithelial neoplasms. Scand. J. Work Environ. Health 1989, 15, 47–53. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, P.A.; Delzell, E.; Sathiakumar, N.; Myers, S.L.; Cheng, H.; Grizzle, W.; Chen, V.W.; Wu, X.C. Cancer incidence among triazine herbicide manufacturing workers. J. Occup. Environ. Med. 2002, 44, 1048–1058. [Google Scholar] [CrossRef] [PubMed]
- Gammon, D.W.; Aldous, C.N.; Carr, W.C., Jr.; Sanborn, J.R.; Pfeifer, K.F. A risk assessment of atrazine use in California: Human health and ecological aspects. Pest Manag. Sci. 2005, 61, 331–355. [Google Scholar] [CrossRef] [PubMed]
- Meisner, L.F.; Roloff, B.D.; Belluck, D.A. In vitro effects of N-nitrosoatrazine on chromosome breakage. Arch. Environ. Contam. Toxicol. 1993, 24, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Ribas, G.; Frenzilli, G.; Barale, R.; Marcos, R. Herbicide-induced DNA damage in human lymphocytes evaluated by the single-cell gel electrophoresis (SCGE) assay. Mutat. Res. 1995, 344, 41–54. [Google Scholar] [CrossRef]
- Clements, C.; Ralph, S.; Petras, M. Genotoxicity of select herbicides in Rana catesbeiana tadpoles using the alkaline single-cell gel DNA electrophoresis (comet) assay. Environ. Mol. Mutagen. 1997, 29, 277–288. [Google Scholar] [CrossRef]
- Lioi, M.B.; Scarfi, M.R.; Santoro, A.; Barbieri, R.; Zeni, O.; Salvemini, F.; di Berardino, D.; Ursini, M.V. Cytogenetic damage and induction of pro-oxidant state in human lymphocytes exposed in vitro to gliphosate, vinclozolin, atrazine, and DPX-E9636. Environ. Mol. Mutagen. 1998, 32, 39–46. [Google Scholar] [CrossRef]
- Roloff, B.D.; Belluck, D.A.; Meisner, L.F. Cytogenetic studies of herbicide interactions in vitro and in vivo using atrazine and linuron. Arch. Environ. Contam. Toxicol. 1992, 22, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Dunkelberg, H.; Fuchs, J.; Hengstler, J.G.; Klein, E.; Oesch, F.; Struder, K. Genotoxic effects of the herbicides alachlor, atrazine, pendimethaline, and simazine in mammalian cells. Bull. Environ. Contam. Toxicol. 1994, 52, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Kligerman, A.D.; Doerr, C.L.; Tennant, A.H.; Peng, B. Cytogenetic studies of three triazine herbicides. II. In vivo micronucleus studies in mouse bone marrow. Mutat. Res. 2000, 471, 107–112. [Google Scholar]
- Tennant, A.H.; Peng, B.; Kligerman, A.D. Genotoxicity studies of three triazine herbicides: In vivo studies using the alkaline single cell gel (SCG) assay. Mutat. Res. 2001, 493, 1–10. [Google Scholar] [CrossRef]
- Rai, R.; Peng, G.; Li, K.; Lin, S.Y. DNA damage response: The players, the network and the role in tumor suppression. Cancer Genomics Proteomics 2007, 4, 99–106. [Google Scholar] [PubMed]
- Lin, S.Y.; Rai, R.; Li, K.; Xu, Z.X.; Elledge, S.J. BRIT1/MCPH1 is a DNA damage responsive protein that regulates the Brca1-Chk1 pathway, implicating checkpoint dysfunction in microcephaly. Proc. Natl. Acad. Sci. USA 2005, 102, 15105–15109. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.M.; Shao, J.Z.; Xiang, L.X.; Chen, X.Y. Cytotoxic effects and apoptosis induction of atrazine in a grass carp (Ctenopharyngodon idellus) cell line. Environ. Toxicol. 2006, 21, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.W.; Elledge, S.J. The DNA damage response: Ten years after. Mol. Cell 2007, 28, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Tibbetts, R.S.; Brumbaugh, K.M.; Fang, Y.; Richardson, D.A.; Ali, A.; Chen, S.M.; Abraham, R.T.; Wang, X.F. ATR/ATM-mediated phosphorylation of human Rad17 is required for genotoxic stress responses. Nature 2001, 411, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Guntuku, S.; Cui, X.S.; Matsuoka, S.; Cortez, D.; Tamai, K.; Luo, G.; Carattini-Rivera, S.; DeMayo, F.; Bradley, A.; et al. Chk1 is an essential kinase that is regulated by Atr and required for the G2/M DNA damage checkpoint. Genes Dev. 2000, 14, 1448–1459. [Google Scholar] [PubMed]
- Melo, J.A.; Cohen, J.; Toczyski, D.P. Two checkpoint complexes are independently recruited to sites of DNA damage in vivo. Genes Dev. 2001, 15, 2809–2821. [Google Scholar] [PubMed]
- Ghosh, S.; Narang, H.; Sarma, A.; Krishna, M. DNA damage response signaling in lung adenocarcinoma A549 cells following γ and carbon beam irradiation. Mutat. Res. 2011, 716, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Valentine, J.M.; Kumar, S.; Moumen, A. A p53-independent role for the MDM2 antagonist Nutlin-3 in DNA damage response initiation. BMC Cancer 2011, 11, 79. [Google Scholar] [CrossRef] [PubMed]
- Cavas, T. In vivo genotoxicity evaluation of atrazine and atrazine-based herbicide on fish Carassius auratus using the micronucleus test and the comet assay. Food Chem. Toxicol. 2011, 49, 1431–1435. [Google Scholar] [CrossRef] [PubMed]
- Zeljezic, D.; Garaj-Vrhovac, V.; Perkovic, P. Evaluation of DNA damage induced by atrazine and atrazine-based herbicide in human lymphocytes in vitro using a comet and DNA diffusion assay. Toxicol. In Vitro 2006, 20, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Laure Khoury, L.; Zalko, D.; Audebert, M. Validation of high-throughput genotoxicity assay screening using cH2AX in-cell Western assay on HepG2 cells. Environ. Mol. Mutagen. 2013, 54, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Mah, L.J.; El-Osta, A.; Karagiannis, T.C. γH2AX: A sensitive molecular marker of DNA damage and repair. Leukemia 2010, 24, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Hakem, R. DNA-damage repair; The good, the bad, and the ugly. EMBO J. 2008, 27, 589–605. [Google Scholar] [CrossRef] [PubMed]
- Abarikwu, S.O.; Farombi, E.O.; Kashyap, M.P.; Pant, A.B. Kolaviron protects apoptotic cell death in PC12 cells exposed to atrazine. Free Radic. Res. 2011, 45, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Abarikwu, S.O.; Pant, A.B.; Farombi, E.O. Dietary antioxidant, quercetin, protects sertoli-germ cell coculture from atrazine-induced oxidative damage. J. Biochem. Mol. Toxicol. 2012, 26, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Tchounwou, P.B.; Wilson, B.A.; Ishaque, A.B.; Schneider, J. Atrazine potentiation of arsenic trioxide-induced cytotoxicity and gene expression in human liver carcinoma cells (HepG2). Mol. Cell. Biochem. 2001, 222, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Yang, J.; Song, Q. Atrazine affects phosphoprotein and protein expression in MCF-10A human breast epithelial cells. Int. J. Mol. Sci. 2014, 15, 17806–17826. [Google Scholar] [CrossRef] [PubMed]
- Kansara, K.; Patel, P.; Shah, D.; Shukla, R.K.; Singh, S.; Kumar, A.; Dhawan, A. TiO2 nanoparticles induce DNA double strand breaks and cell cycle arrest in human alveolar cells. Environ. Mol. Mutagen. 2015, 56, 204–217. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, P.; Yang, J.; Ning, J.; Wang, M.; Song, Q. Atrazine Triggers DNA Damage Response and Induces DNA Double-Strand Breaks in MCF-10A Cells. Int. J. Mol. Sci. 2015, 16, 14353-14368. https://doi.org/10.3390/ijms160714353
Huang P, Yang J, Ning J, Wang M, Song Q. Atrazine Triggers DNA Damage Response and Induces DNA Double-Strand Breaks in MCF-10A Cells. International Journal of Molecular Sciences. 2015; 16(7):14353-14368. https://doi.org/10.3390/ijms160714353
Chicago/Turabian StyleHuang, Peixin, John Yang, Jie Ning, Michael Wang, and Qisheng Song. 2015. "Atrazine Triggers DNA Damage Response and Induces DNA Double-Strand Breaks in MCF-10A Cells" International Journal of Molecular Sciences 16, no. 7: 14353-14368. https://doi.org/10.3390/ijms160714353