
Plant MYB Transcription Factors: Their Role in Drought Response Mechanisms

Abstract
:
1. Introduction

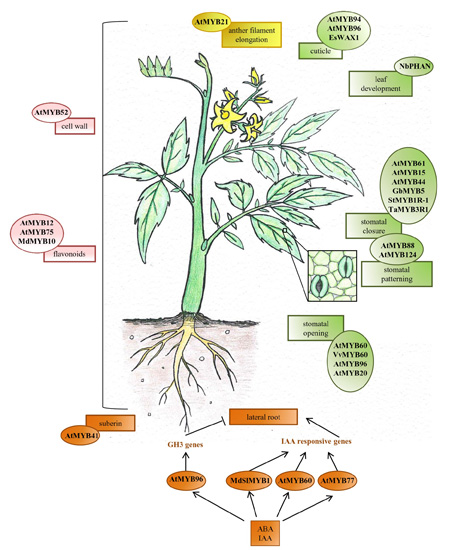{kind=link}
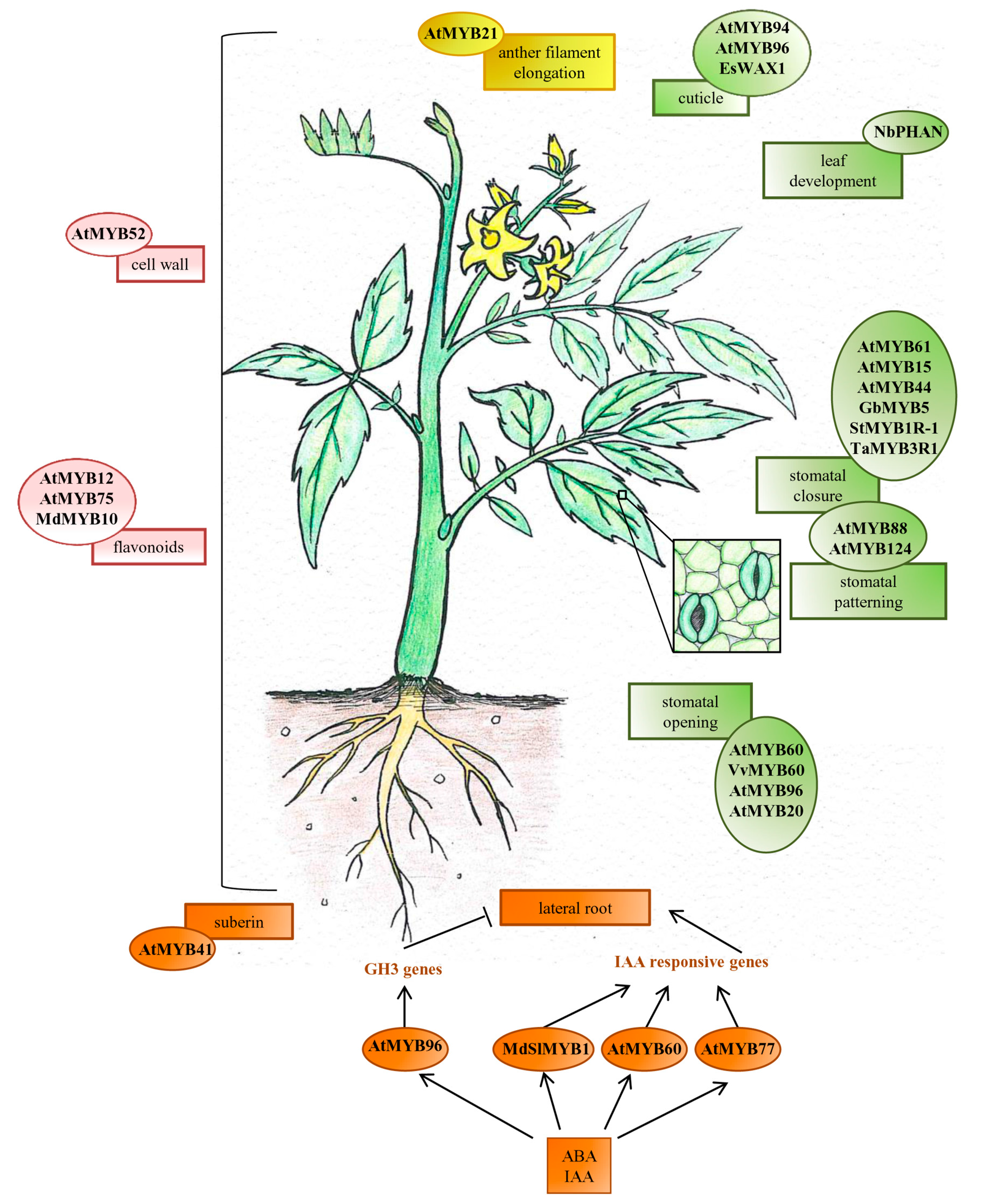{kind=link}
| Gene Name | Locus/Accession Nr | Species | Transgenic Plants/Mutants | References |
|---|---|---|---|---|
| Root | ||||
| AtMYB77 | AT3G50060 | Arabidopsis thaliana | Arabidopsis KO | [12,13] |
| AtMYB60 | AT1G08810 | Arabidopsis thaliana | Arabidopsis OE | [14] |
| AtMYB96 | AT5G62470 | Arabidopsis thaliana | Arabidopsis activation tagged OE | [15] |
| MdSIMYB1 | KC691248 | Malus x domestica | apple OE, tobacco OE | [16] |
| Leaf | ||||
| NbPHAN | FR878011 | Nicotiana benthamiana | N. benthamiana VIGS | [17] |
| Stomata | ||||
| AtMYB88 | AT2G02820 | Arabidopsis thaliana | Arabidopsis KO | [18,19,20,21,22] |
| FLP/AtMYB124 | AT1G14350 | Arabidopsis thaliana | Arabidopsis KO | [18,19,20,21,22] |
| AtMYB60 | AT1G08810 | Arabidopsis thaliana | Arabidopsis OE, KO | [14,23] |
| VvMYB60 | ACF21938 | Vitis vinifera | complementation of Arabidopsis mutant | [24] |
| AtMYB96 | AT5G62470 | Arabidopsis thaliana | Arabidopsis activation tagged OE, KO | [15,25] |
| AtMYB20 | AT1G66230 | Arabidopsis thaliana | Arabidopsis OE, KO | [26] |
| AtMYB61 | AT1G09540 | Arabidopsis thaliana | Arabidopsis OE, KO | [27,28,29,30] |
| AtMYB15 | AT3G23250 | Arabidopsis thaliana | Arabidopsis OE | [31] |
| AtMYB44 | AT5G67300 | Arabidopsis thaliana | Arabidopsis OE | [32] |
| GbMYB5 | JF820389 | Gossypium barbadense | cotton VIGS, tobacco OE | [33] |
| StMYB1R-1 | AU279205 | Solanum tuberosum | Potato OE | [11] |
| TaMYB3R1 | HQ236494 | Triticum aestivum | Arabidopsis OE | [10] |
| Flower | ||||
| AtMYB21 | AT3G27810 | Arabidopsis thaliana | Arabidopsis KO | [34,35,36] |
| CmMYB2 | JF795918 | Chrysanthemum morifolium | Arabidopsis OE | [37] |
| Cell Wall | ||||
| AtMYB52 | AT1G17950 | Arabidopsis thaliana | Arabidopsis OE, activation tagged OE | [38] |
| Cuticle | ||||
| AtMYB94 | AT3G47600 | Arabidopsis thaliana | Arabidopsis OE | [39] |
| AtMYB96 | AT5G62470 | Arabidopsis thaliana | Arabidopsis OE; Camelina OE | [15,40,41] |
| EsWAX1 | BAJ34253 | Eutrema salsugineum | Arabidopsis OE | [42] |
| Suberin | ||||
| AtMYB41 | AT4G28110 | Arabidopsis thaliana | Arabidopsis OE | [43,44,45,46] |
| Flavonoids | ||||
| AtMYB12/PFG1 | AT2G47460 | Arabidopsis thaliana | Arabidopsis OE | [47] |
| AtMYB75/PAP1 | AT1G56650 | Arabidopsis thaliana | Arabidopsis OE | [47] |
| MdMYB10 | ABB84753 | Malus x domestica | Arabidopsis OE | [48,49] |
| Abiotic Stresses | ||||
| AtMYB2 | AT2G47190 | Arabidopsis thaliana | Arabidopsis OE, KO | [50,51,52,53] |
| AtMYB20 | AT1G66230 | Arabidopsis thaliana | Arabidopsis OE, KO | [26,54] |
| OsMYB3R-2 | LOC_Os01g62410/ Os01g0841500 | Oryza sativa | Arabidopsis OE | [9,55] |
| OsMYB2 | LOC_Os03g20090/ Os03g0315400 | Oryza sativa | Rice OE, RNAi | [56] |
| OsMYB48-1 | LOC_Os01g74410/ Os01g0975300 | Oryza sativa | Rice OE | [57,58] |
| TaMYB2A | AY615199 | Triticum aestivum | Arabidopsis OE | [59] |
| TaMYB19 | JF951903 | Triticum aestivum | Arabidopsis OE | [60] |
| TaMYB30-B | JF951913 | Triticum aestivum | Arabidopsis OE | [61] |
| TaMYB33 | JN584645 | Triticum aestivum | Arabidopsis OE | [62] |
| GmMYBJ1 | KC751453 | Glycine max | Arabidopsis OE | [63] |
| CpMYB10 | AF510112 | Craterostigma plantagineum | Arabidopsis OE | [64] |
| MdoMYB121 | KC834015 | Malus x domestica | apple OE; tomato OE | [65] |
| MdSIMYB1 | KC691248 | Malus x domestica | apple OE, tobacco OE | [16] |
| PtsrMYB | – | Poncirus trifoliata | tobacco OE | [66] |
| Abiotic and Biotic Stresses | ||||
| AtMYB12/PFG1 | AT2G47460 | Arabidopsis thaliana | tobacco OE | [67,68,69] |
| AtMYB75/PAP1 | AT1G56650 | Arabidopsis thaliana | tobacco OE | [68] |
| AtMYB44 | AT5G67300 | Arabidopsis thaliana | Arabidopsis OE, KO; soybean OE | [32,70,71,72,73] |
| AtMYB96 | AT5G62470 | Arabidopsis thaliana | Arabidopsis activation tagged OE | [74,75] |
| AtBOS1/MYB108 | AT3G06490 | Arabidopsis thaliana | Arabidopsis KO | [76] |
| AtMYB15 | AT3G23250 | Arabidopsis thaliana | Arabidopsis OE, KO | [77,78,79,80,81] |
| OsMYB4 | LOC_Os04g43680/ Os04g0517100 | Oryza sativa | OE in rice, Arabidopsis, apple, tomato, potato, sage, tobacco, Osteospermum, barley | [82,83,84,85,86,87,88,89,90,91,92] |
| TaPIMP1 | CN011324 | Triticum aestivum | tobacco OE; wheat OE, RNAi | [93,94] |
| SpMYB | – | Solanum pimpinellifolium | tobacco OE | [95] |
| Other | ||||
| QsMYB1 | JF970262 and JF970262 | Quercus suber | – | [96,97] |
| AtMYB33 | AT5G06100 | Arabidopsis thaliana | – | [98,99,100,101] |
| AtMYB101 | AT2G32460 | Arabidopsis thaliana | – | [98,99,100,101] |
| StGAMyb-like1 | – | Solanum tuberosum | – | [102] |
| StGAMyb-like2.1 | – | Solanum tuberosum | – | [102] |
| StGAMyb-like2.2 | – | Solanum tuberosum | – | [102] |
| TaMYB2 | AB252145 | Triticum aestivum | – | [103] |
| QsMYB1 | JF970262 and JF970262 | Quercus suber | – | [96,97] |
2. Development, Growth and Function of Organs and Specific Cell-Types
2.1. Root

2.2. Leaf
2.3. Stomata
2.4. Flower
3. Metabolite Biosynthesis
3.1. Cell Wall Components
3.2. Cuticle and Suberin
3.3. Flavonoids
4. Crosstalk among Different Stress Responses
| Gene Name | Abiotic Stresses | Biotic Stresses | Other Stresses | |||
|---|---|---|---|---|---|---|
| Salt | Cold/Freezing | Bacteria | Fungi | Insects | ||
| AtMYB2 | – | – | – | – | – | low oxygen, Pi deficiency |
| AtMYB20 | × | – | – | – | – | – |
| AtMYB12 | – | – | – | – | S. litura | UV-B |
| PFG1 | H. armigera | flg 22 | ||||
| AtMYB75 | – | – | – | – | S. litura | – |
| PAP1 | ||||||
| AtMYB44 | × | – | P. syringae | – | M. persicae | wounding |
| P. xylostella | ||||||
| AtMYB96 | – | × | P. syringae | – | – | flg 22 |
| AtBOS1 | × | × | – | B. cinerea | – | oxidative |
| MYB108 | ||||||
| AtMYB15 | × | × | – | – | M. persicae | wounding |
| HrpNEa | ||||||
| OsMYB3R-2 | × | × | – | – | – | – |
| OsMYB2 | × | × | – | – | – | – |
| OsMYB48-1 | × | – | – | – | – | – |
| OsMYB4 | × | × | P. syringae | B. cinerea | – | oxidative |
| TNV | ||||||
| TaMYB2A | × | × | – | – | – | – |
| TaMYB19 | × | × | – | – | – | – |
| TaMYB33 | × | – | – | – | – | – |
| TaPIMP1 | × | – | R. solanacearum | B. sorokiniana | – | oxidative |
| GmMYBJ1 | – | × | – | – | – | – |
| CpMYB10 | × | – | – | – | – | – |
| MdSIMYB1 | × | × | – | – | – | – |
| MdoMYB121 | × | × | – | – | – | – |
| SpMYB | × | – | – | A. alternata | – | – |
4.1. Crosstalk among Different Abiotic Stress Responses
4.2. Crosstalk among Abiotic and Biotic Stress Responses
5. Post-Transcriptional Control of Some MYB Genes Involved in Drought Stress Response
6. Conclusions and Future Perspectives
Acknowledgments
Conflicts of Interest
References
- Pennisi, E. Plant genetics. Getting to the root of drought responses. Science 2008, 320, 173. [Google Scholar] [CrossRef] [PubMed]
- National Centers for Environmental Information. Available online: http://www.ncdc.noaa.gov/billions/events (accessed on 30 June 2015).
- Li, J.; Lin, X.; Chen, A.; Peterson, T.; Ma, K.; Bertzky, M.; Ciais, P.; Kapos, V.; Peng, C.; Poulter, B. Global priority conservation areas in the face of 21st century climate change. PLoS ONE 2013, 8, e54839. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Lawlor, D.W. Genetic engineering to improve plant performance under drought: Physiological evaluation of achievements, limitations, and possibilities. J. Exp. Bot. 2013, 64, 83–108. [Google Scholar] [CrossRef] [PubMed]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 2014, 5, 151. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ng, C.K.-Y.; Fan, L.-M. MYB transcription factors, active players in abiotic stress signaling. Environ. Exp. Bot. 2015, 114, 80–91. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Xu, Y.; Ma, Q.; Xu, W.; Wang, T.; Xue, Y.; Chong, K. Overexpression of an R1R2R3 MYB gene, OsMYB3R-2, increases tolerance to freezing, drought, and salt stress in transgenic Arabidopsis. Plant Physiol. 2007, 143, 1739–1751. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Tian, S.; Dong, H.; Guo, C. Pleiotropic effects of TaMYB3R1 on plant development and response to osmotic stress in transgenic Arabidopsis. Gene 2015, 558, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.; Moon, S.J.; Han, S.; Kim, B.G.; Park, S.R.; Lee, S.K.; Yoon, H.J.; Lee, H.E.; Kwon, H.B.; Baek, D.; et al. Expression of StMYB1R-1, a novel potato single MYB-like domain transcription factor, increases drought tolerance. Plant Physiol. 2011, 155, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xing, L.; Wang, X.; Hou, Y.J.; Gao, J.; Wang, P.; Duan, C.G.; Zhu, X.; Zhu, J.K. The ABA receptor PYL8 promotes lateral root growth by enhancing MYB77-dependent transcription of auxin-responsive genes. Sci. Signal. 2014, 7, ra53. [Google Scholar] [CrossRef] [PubMed]
- Shin, R.; Burch, A.Y.; Huppert, K.A.; Tiwari, S.B.; Murphy, A.S.; Guilfoyle, T.J.; Schachtman, D.P. The Arabidopsis transcription factor MYB77 modulates auxin signal transduction. Plant Cell 2007, 19, 2440–2453. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.E.; Kwon, Y.; Kim, J.H.; Noh, H.; Hong, S.W.; Lee, H. A dual role for MYB60 in stomatal regulation and root growth of Arabidopsis thaliana under drought stress. Plant Mol. Biol. 2011, 77, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Seo, P.J.; Xiang, F.; Qiao, M.; Park, J.Y.; Lee, Y.N.; Kim, S.G.; Lee, Y.H.; Park, W.J.; Park, C.M. The MYB96 transcription factor mediates abscisic acid signaling during drought stress response in Arabidopsis. Plant Physiol. 2009, 151, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.K.; Cao, Z.H.; Hao, Y.J. Overexpression of a R2R3 MYB gene MdSIMYB1 increases tolerance to multiple stresses in transgenic tobacco and apples. Physiol. Plant 2014, 150, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Hu, G.; Li, F.; Li, Y.; Wu, J.; Zhou, X. NbPHAN, a MYB transcriptional factor, regulates leaf development and affects drought tolerance in Nicotiana benthamiana. Physiol. Plant 2013, 149, 297–309. [Google Scholar] [PubMed]
- Lai, L.B.; Nadeau, J.A.; Lucas, J.; Lee, E.K.; Nakagawa, T.; Zhao, L.; Geisler, M.; Sack, F.D. The Arabidopsis R2R3 MYB proteins FOUR LIPS and MYB88 restrict divisions late in the stomatal cell lineage. Plant Cell 2005, 17, 2754–2767. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Lee, E.; Lucas, J.R.; Morohashi, K.; Li, D.; Murray, J.A.; Sack, F.D.; Grotewold, E. Regulation of cell proliferation in the stomatal lineage by the Arabidopsis MYB FOUR LIPS via direct targeting of core cell cycle genes. Plant Cell 2010, 22, 2306–2321. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, S.; Coppens, F.; Lee, E.; Donner, T.J.; Xie, Z.; van Isterdael, G.; Dhondt, S.; de Winter, F.; de Rybel, B.; Vuylsteke, M.; et al. Developmental regulation of CYCA2s contributes to tissue-specific proliferation in Arabidopsis. EMBO J. 2011, 30, 3430–3441. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Li, D.; Wang, L.; Sack, F.D.; Grotewold, E. Role of the stomatal development regulators FLP/MYB88 in abiotic stress responses. Plant J. 2010, 64, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Bhave, N.S.; Veley, K.M.; Nadeau, J.A.; Lucas, J.R.; Bhave, S.L.; Sack, F.D. TOO MANY MOUTHS promotes cell fate progression in stomatal development of Arabidopsis stems. Planta 2009, 229, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Cominelli, E.; Galbiati, M.; Vavasseur, A.; Conti, L.; Sala, T.; Vuylsteke, M.; Leonhardt, N.; Dellaporta, S.L.; Tonelli, C. A guard-cell-specific MYB transcription factor regulates stomatal movements and plant drought tolerance. Curr. Biol. 2005, 15, 1196–1200. [Google Scholar] [CrossRef] [PubMed]
- Galbiati, M.; Matus, J.T.; Francia, P.; Rusconi, F.; Cañón, P.; Medina, C.; Conti, L.; Cominelli, E.; Tonelli, C.; Arce-Johnson, P. The grapevine guard cell-related VvMYB60 transcription factor is involved in the regulation of stomatal activity and is differentially expressed in response to ABA and osmotic stress. BMC Plant Biol. 2011, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. The plant hormone abscisic acid mediates the drought-induced expression but not the seed-specific expression of rd22, a gene responsive to dehydration stress in Arabidopsis thaliana. Mol. Gen. Genet. 1993, 238, 17–25. [Google Scholar] [PubMed]
- Gao, S.; Zhang, Y.; Yang, L.; Song, J.; Yang, Z. AtMYB20 is negatively involved in plant adaptive response to drought stress. Plant Soil 2014, 376, 433–443. [Google Scholar] [CrossRef]
- Newman, L.J.; Perazza, D.E.; Juda, L.; Campbell, M.M. Involvement of the R2R3-MYB, AtMYB61, in the ectopic lignification and dark-photomorphogenic components of the det3 mutant phenotype. Plant J. 2004, 37, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Penfield, S.; Meissner, R.C.; Shoue, D.A.; Carpita, N.C.; Bevan, M.W. MYB61 is required for mucilage deposition and extrusion in the Arabidopsis seed coat. Plant Cell 2001, 13, 2777–2791. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.K.; Dubos, C.; Dodd, I.C.; Holroyd, G.H.; Hetherington, A.M.; Campbell, M.M. AtMYB61, an R2R3-MYB transcription factor controlling stomatal aperture in Arabidopsis thaliana. Curr. Biol. 2005, 15, 1201–1206. [Google Scholar] [CrossRef] [PubMed]
- Romano, J.M.; Dubos, C.; Prouse, M.B.; Wilkins, O.; Hong, H.; Poole, M.; Kang, K.Y.; Li, E.; Douglas, C.J.; Western, T.L.; et al. AtMYB61, an R2R3-MYB transcription factor, functions as a pleiotropic regulator via a small gene network. New Phytol. 2012, 195, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Li, S.; An, X.; Liu, X.; Qin, H.; Wang, D. Transgenic expression of MYB15 confers enhanced sensitivity to abscisic acid and improved drought tolerance in Arabidopsis thaliana. J. Genet. Genomics 2009, 36, 17–29. [Google Scholar] [CrossRef]
- Jung, C.; Seo, J.S.; Han, S.W.; Koo, Y.J.; Kim, C.H.; Song, S.I.; Nahm, B.H.; Choi, Y.D.; Cheong, J.J. Overexpression of AtMYB44 enhances stomatal closure to confer abiotic stress tolerance in transgenic Arabidopsis. Plant Physiol. 2008, 146, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Li, W.; Hu, X.; Guo, J.; Liu, A.; Zhang, B. A cotton MYB transcription factor, GbMYB5, is positively involved in plant adaptive response to drought stress. Plant Cell Physiol. 2015, 56, 917–929. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Ma, X.; Guo, H.; Sukiran, N.; Guo, B.; Assmann, S.; Ma, H. Flower development under drought stress: Morphological and transcriptomic analyses reveal acute responses and long-term acclimation in Arabidopsis. Plant Cell 2013, 25, 3785–3807. [Google Scholar] [CrossRef] [PubMed]
- Mandaokar, A.; Thines, B.; Shin, B.; Lange, B.; Choi, G.; Koo, Y.; Yoo, Y.; Choi, Y.; Browse, J. Transcriptional regulators of stamen development in Arabidopsis identified by transcriptional profiling. Plant J. 2006, 46, 984–1008. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Song, S.; Xiao, L.; Soo, H.; Cheng, Z.; Xie, D.; Peng, J. Gibberellin acts through jasmonate to control the expression of MYB21, MYB24, and MYB57 to promote stamen filament growth in Arabidopsis. PLoS Genet. 2009, 5, e1000440. [Google Scholar] [CrossRef] [PubMed]
- Shan, H.; Chen, S.; Jiang, J.; Chen, F.; Chen, Y.; Gu, C.; Li, P.; Song, A.; Zhu, X.; Gao, H.; et al. Heterologous expression of the chrysanthemum R2R3-MYB transcription factor CmMYB2 enhances drought and salinity tolerance, increases hypersensitivity to ABA and delays flowering in Arabidopsis thaliana. Mol. Biotechnol. 2012, 51, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Kang, J.; Kim, S. Overexpression of AtMYB52 confers ABA hypersensitivity and drought tolerance. Mol. Cells 2011, 31, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Suh, M.C. Cuticular wax biosynthesis is up-regulated by the MYB94 transcription factor in Arabidopsis. Plant Cell Physiol. 2015, 56, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Seo, P.J.; Lee, S.B.; Suh, M.C.; Park, M.J.; Go, Y.S.; Park, C.M. The MYB96 transcription factor regulates cuticular wax biosynthesis under drought conditions in Arabidopsis. Plant Cell 2011, 23, 1138–1152. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Kim, H.; Kim, R.J.; Suh, M.C. Overexpression of Arabidopsis MYB96 confers drought resistance in Camelina sativa via cuticular wax accumulation. Plant Cell Rep. 2014, 33, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Guo, J.; Zhu, J.; Zhou, C. Enhanced expression of EsWAX1 improves drought tolerance with increased accumulation of cuticular wax and ascorbic acid in transgenic Arabidopsis. Plant Physiol. Biochem. 2014, 75, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Kosma, D.K.; Murmu, J.; Razeq, F.M.; Santos, P.; Bourgault, R.; Molina, I.; Rowland, O. AtMYB41 activates ectopic suberin synthesis and assembly in multiple plant species and cell types. Plant J. 2014, 80, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Cominelli, E.; Sala, T.; Calvi, D.; Gusmaroli, G.; Tonelli, C. Over-expression of the Arabidopsis AtMYB41 gene alters cell expansion and leaf surface permeability. Plant J. 2008, 53, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Lippold, F.; Sanchez, D.H.; Musialak, M.; Schlereth, A.; Scheible, W.R.; Hincha, D.K.; Udvardi, M.K. AtMYB41 regulates transcriptional and metabolic responses to osmotic stress in Arabidopsis. Plant Physiol. 2009, 149, 1761–1772. [Google Scholar] [CrossRef] [PubMed]
- Hoang, M.H.; Nguyen, X.C.; Lee, K.; Kwon, Y.S.; Pham, H.T.; Park, H.C.; Yun, D.J.; Lim, C.O.; Chung, W.S. Phosphorylation by AtMPK6 is required for the biological function of AtMYB41 in Arabidopsis. Biochem. Biophys. Res. Commun. 2012, 422, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, R.; Yonekura-Sakakibara, K.; Urano, K.; Suzuki, M.; Yamada, Y.; Nishizawa, T.; Matsuda, F.; Kojima, M.; Sakakibara, H.; Shinozaki, K.; et al. Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J. 2014, 77, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Espley, R.V.; Hellens, R.P.; Putterill, J.; Stevenson, D.E.; Kutty-Amma, S.; Allan, A.C. Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J. 2007, 49, 414–427. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.J.; Zhang, Z.; Peng, R.H.; Xiong, A.S.; Xu, J.; Zhu, B.; Yao, Q.H. Forced expression of MdMYB10, a MYB transcription factor gene from apple, enhances tolerance to osmotic stress in transgenic Arabidopsis. Mol. Biol. Rep. 2011, 38, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Abe, H.; Yamaguchi-Shinozaki, K.; Urao, T.; Iwasaki, T.; Hosokawa, D.; Shinozaki, K. Role of Arabidopsis MYC and MYB homologs in drought- and abscisic acid-regulated gene expression. Plant Cell 1997, 9, 1859–1868. [Google Scholar] [PubMed]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Hoeren, F.U.; Dolferus, R.; Wu, Y.; Peacock, W.J.; Dennis, E.S. Evidence for a role for AtMYB2 in the induction of the Arabidopsis alcohol dehydrogenase gene (Adh1) by low oxygen. Genetics 1998, 149, 479–490. [Google Scholar] [PubMed]
- Baek, D.; Kim, M.C.; Chun, H.J.; Kang, S.; Park, H.C.; Shin, G.; Park, J.; Shen, M.; Hong, H.; Kim, W.Y.; et al. Regulation of mir399f transcription by AtMYB2 affects phosphate starvation responses in Arabidopsis. Plant Physiol. 2013, 161, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Yoo, K.; Hyoung, S.; Nguyen, H.; Kim, Y.; Kim, H.; Ok, S.; Yoo, S.; Shin, J. An Arabidopsis R2R3-MYB transcription factor, AtMYB20, negatively regulates type 2C serine/threonine protein phosphatases to enhance salt tolerance. FEBS Lett. 2013, 587, 1773–1778. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Dai, X.; Xu, Y.; Guo, J.; Liu, Y.; Chen, N.; Xiao, J.; Zhang, D.; Xu, Z.; Zhang, X.; et al. Enhanced tolerance to chilling stress in OsMYB3R-2 transgenic rice is mediated by alteration in cell cycle and ectopic expression of stress genes. Plant Physiol. 2009, 150, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Dai, X.; Zhang, W. A R2R3-type MYB gene, OsMYB2, is involved in salt, cold, and dehydration tolerance in rice. J. Exp. Bot. 2012, 63, 2541–2556. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Li, J.; Liu, P.; Duan, J.; Zhao, Y.; Guo, X.; Li, Y.; Zhang, H.; Ali, J.; Li, Z. Overexpression of OsMYB48-1, a novel MYB-related transcription factor, enhances drought and salinity tolerance in rice. PLoS ONE 2014, 9, e92913. [Google Scholar] [CrossRef] [PubMed]
- Quan, R.; Hu, S.; Zhang, Z.; Zhang, H.; Zhang, Z.; Huang, R. Overexpression of an ERF transcription factor TSRF1 improves rice drought tolerance. Plant Biotechnol. J. 2010, 8, 476–488. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Jia, D.; Li, A.; Zhang, H.; Tian, S.; Zhang, X.; Jia, J.; Jing, R. Transgenic expression of TaMYB2A confers enhanced tolerance to multiple abiotic stresses in Arabidopsis. Funct. Integr. Genomics 2011, 11, 445–465. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, G.; Zhao, G.; Xia, C.; Jia, J.; Liu, X.; Kong, X. Characterization of a wheat R2R3-MYB transcription factor gene, TaMYB19, involved in enhanced abiotic stresses in Arabidopsis. Plant Cell Physiol. 2014, 55, 1802–1812. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, G.; Xia, C.; Jia, J.; Liu, X.; Kong, X. A wheat R2R3-MYB gene, TaMYB30-B, improves drought stress tolerance in transgenic Arabidopsis. J. Exp. Bot. 2012, 63, 5873–5885. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Wang, M.; Tian, Y.; He, W.; Han, L.; Xia, G. Over-expression of TaMYB33 encoding a novel wheat MYB transcription factor increases salt and drought tolerance in Arabidopsis. Mol. Biol. Rep. 2012, 39, 7183–7192. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Li, J.; Liu, D.; Zhai, Y.; Zhang, H.; Li, X.; Zhang, Q.; Wang, Y.; Wang, Q. A novel MYB transcription factor, GmMYBj1, from soybean confers drought and cold tolerance in Arabidopsis thaliana. Gene 2014, 538, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Villalobos, M.; Bartels, D.; Iturriaga, G. Stress tolerance and glucose insensitive phenotypes in Arabidopsis overexpressing the CpMYB10 transcription factor gene. Plant Physiol. 2004, 135, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Zhang, S.; Wang, R.; Zhang, R.; Hao, Y. Genome wide analysis of the apple MYB transcription factor family allows the identification of MdoMYB121 gene confering abiotic stress tolerance in plants. PLoS ONE 2013, 8, e69955. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Zhu, X.; Huang, X.; Liu, J. Overexpression of a stress-responsive MYB transcription factor of Poncirus trifoliata confers enhanced dehydration tolerance and increases polyamine biosynthesis. Plant Physiol. Biochem. 2014, 78, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Misra, P.; Pandey, A.; Tiwari, M.; Chandrashekar, K.; Sidhu, O.P.; Asif, M.H.; Chakrabarty, D.; Singh, P.K.; Trivedi, P.K.; Nath, P.; et al. Modulation of transcriptome and metabolome of tobacco by Arabidopsis transcription factor, AtMYB12, leads to insect resistance. Plant Physiol. 2010, 152, 2258–2268. [Google Scholar] [CrossRef] [PubMed]
- Mitsunami, T.; Nishihara, M.; Galis, I.; Alamgir, K.M.; Hojo, Y.; Fujita, K.; Sasaki, N.; Nemoto, K.; Sawasaki, T.; Arimura, G. Overexpression of the PAP1 transcription factor reveals a complex regulation of flavonoid and phenylpropanoid metabolism in Nicotiana tabacum plants attacked by Spodoptera litura. PLoS ONE 2014, 9, e108849. [Google Scholar] [CrossRef] [PubMed]
- Schenke, D.; Böttcher, C.; Scheel, D. Crosstalk between abiotic ultraviolet-B stress and biotic (flg22) stress signalling in Arabidopsis prevents flavonol accumulation in favor of pathogen defence compound production. Plant Cell Environ. 2011, 34, 1849–1864. [Google Scholar] [CrossRef] [PubMed]
- Persak, H.; Pitzschke, A. Tight interconnection and multi-level control of Arabidopsis MYB44 in MAPK cascade signalling. PLoS ONE 2013, 8, e57547. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Sohn, H.; Noh, K.; Jung, C.; An, J.; Donovan, C.; Somers, D.; Kim, D.; Jeong, S.; Kim, C.; et al. Expression of the Arabidopsis AtMYB44 gene confers drought/salt-stress tolerance in transgenic soybean. Mol. Breed. 2012, 29, 601–608. [Google Scholar] [CrossRef]
- Jaradat, M.; Feurtado, J.; Huang, D.; Lu, Y.; Cutler, A. Multiple roles of the transcription factor AtMYBr1/AtMYB44 in ABA signaling, stress responses, and leaf senescence. BMC Plant Biol. 2013, 13, 192. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, Y.; Zhang, L.; Wang, X.; Zhao, Z.; Tao, Z.; Wang, J.; Wang, J.; Lin, M.; Li, X.; et al. Arabidopsis ABA receptor RCAR1/PYL9 interacts with an R2R3-type MYB transcription factor, AtMYB44. Int. J. Mol. Sci. 2014, 15, 8473–8490. [Google Scholar] [CrossRef] [PubMed]
- Seo, P.J.; Park, C.M. MYB96-mediated abscisic acid signals induce pathogen resistance response by promoting salicylic acid biosynthesis in Arabidopsis. New Phytol. 2010, 186, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yang, H.; Zhang, X.; Yang, S. Lipid transfer protein 3 as a target of MYB96 mediates freezing and drought stress in Arabidopsis. J. Exp. Bot. 2013, 64, 1755–1767. [Google Scholar] [CrossRef] [PubMed]
- Mengiste, T.; Chen, X.; Salmeron, J.; Dietrich, R. The Botrytis susceptible1 gene encodes an R2R3MYB transcription factor protein that is required for biotic and abiotic stress responses in Arabidopsis. Plant Cell 2003, 15, 2551–2565. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Jin, J.; Lee, J.; Yoo, C.; Stirm, V.; Miura, T.; Ashworth, E.; Bressan, R.; Yun, D.; Hasegawa, P. SIZ1-mediated sumoylation of ICE1 controls CBF3/DREB1A expression and freezing tolerance in Arabidopsis. Plant Cell 2007, 19, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, M.; Hao, Y.; Kapoor, A.; Dong, C.H.; Fujii, H.; Zheng, X.; Zhu, J.K. A R2R3 type MYB transcription factor is involved in the cold regulation of CBF genes and in acquired freezing tolerance. J. Biol. Chem. 2006, 281, 37636–37645. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liang, C.; Kao, A.; Yang, C. HHP1, a novel signalling component in the cross-talk between the cold and osmotic signalling pathways in Arabidopsis. J. Exp. Bot. 2010, 61, 3305–3320. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, X.; Wu, W.; Chen, Z.; Gu, H.; Qu, L. Overexpression of the wounding-responsive gene AtMYB15 activates the shikimate pathway in Arabidopsis. J. Integr. Plant Biol. 2006, 48, 1084–1095. [Google Scholar] [CrossRef]
- Liu, R.; Lu, B.; Wang, X.; Zhang, C.; Zhang, S.; Qian, J.; Chen, L.; Shi, H.; Dong, H. Thirty-seven transcription factor genes differentially respond to a harpin protein and affect resistance to the green peach aphid in Arabidopsis. J. Biosci. 2010, 35, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Baldoni, E.; Genga, A.; Medici, A.; Coraggio, I.; Locatelli, F. The OsMYB4 gene family: Stress response and transcriptional auto-regulation mechanisms. Biol. Plant. 2013, 57, 691–700. [Google Scholar] [CrossRef]
- Vannini, C.; Locatelli, F.; Bracale, M.; Magnani, E.; Marsoni, M.; Osnato, M.; Mattana, M.; Baldoni, E.; Coraggio, I. Overexpression of the rice OsMYB4 gene increases chilling and freezing tolerance of Arabidopsis thaliana plants. Plant J. 2004, 37, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Mattana, M.; Biazzi, E.; Consonni, R.; Locatelli, F.; Vannini, C.; Provera, S.; Coraggio, I. Overexpression of OsMYB4 enhances compatible solute accumulation and increases stress tolerance of Arabidopsis thaliana. Physiol. Plant. 2005, 125, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Vannini, C.; Iriti, M.; Bracale, M.; Locatelli, F.; Faoro, F.; Croce, P.; Pirona, R.; di Maro, A.; Coraggio, I.; Genga, A. The ectopic expression of the rice OsMYB4 gene in Arabidopsis increases tolerance to abiotic, environmental and biotic stresses. Physiol. Mol. Plant Pathol. 2006, 69, 26–42. [Google Scholar] [CrossRef]
- Park, M.; Yun, K.; Mohanty, B.; Herath, V.; Xu, F.; Wijaya, E.; Bajic, V.; Yun, S.; de Los Reyes, B. Supra-optimal expression of the cold-regulated OsMYB4 transcription factor in transgenic rice changes the complexity of transcriptional network with major effects on stress tolerance and panicle development. Plant Cell Environ. 2010, 33, 2209–2230. [Google Scholar] [CrossRef] [PubMed]
- Vannini, C.; Campa, M.; Iriti, M.; Genga, A.; Faoro, F.; Carravieri, S.; Rotino, G.; Rossoni, M.; Spinardi, A.; Bracale, M. Evaluation of transgenic tomato plants ectopically expressing the rice OsMYB4 gene. Plant Sci. 2007, 173, 231–239. [Google Scholar] [CrossRef]
- Pasquali, G.; Biricolti, S.; Locatelli, F.; Baldoni, E.; Mattana, M. OsMYB4 expression improves adaptive responses to drought and cold stress in transgenic apples. Plant Cell Rep. 2008, 27, 1677–1686. [Google Scholar] [CrossRef] [PubMed]
- Laura, M.; Consonni, R.; Locatelli, F.; Fumagalli, E.; Allavena, A.; Coraggio, I.; Mattana, M. Metabolic response to cold and freezing of Osteospermum ecklonis overexpressing OsMYB4. Plant Physiol. Biochem. 2010, 48, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Docimo, T.; Mattana, M.; Fasano, R.; Consonni, R.; de Tommasi, N.; Coraggio, I.; Leone, A. Ectopic expression of the Osmyb4 rice gene enhances synthesis of hydroxycinnamic acid derivatives in tobacco and clary sage. Biol. Plant. 2013, 57, 179–183. [Google Scholar] [CrossRef]
- Soltesz, A.; Vagujfalvi, A.; Rizza, F.; Kerepesi, I.; Galiba, G.; Cattivelli, L.; Coraggio, I.; Crosatti, C. The rice OsMYB4 gene enhances tolerance to frost and improves germination under unfavourable conditions in transgenic barley plants. J. Appl. Genet. 2012, 53, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Aydin, G.; Yucel, M.; Chan, M.; Oktem, H. Evaluation of abiotic stress tolerance and physiological characteristics of potato (Solanum tuberosum L. Cv. Kennebec) that heterologously expresses the rice OsMYB4 gene. Plant Biotechnol. Rep. 2014, 8, 295–304. [Google Scholar] [CrossRef]
- Liu, H.; Zhou, X.; Dong, N.; Liu, X.; Zhang, H.; Zhang, Z. Expression of a wheat MYB gene in transgenic tobacco enhances resistance to Ralstonia solanacearum, and to drought and salt stresses. Funct. Integr. Genomic 2011, 11, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, X.; Wang, X.; Zhou, M.; Zhou, X.; Ye, X.; Wei, X. An R2R3 MYB transcription factor in wheat, TaPIMP1, mediates host resistance to Bipolaris sorokiniana and drought stresses through regulation of defense- and stress-related genes. New Phytol. 2012, 196, 1155–1170. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Luan, Y.; Yin, Y. SpMYB overexpression in tobacco plants leads to altered abiotic and biotic stress responses. Gene 2014, 547, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Almeida, T.; Menendez, E.; Capote, T.; Ribeiro, T.; Santos, C.; Goncalves, S. Molecular characterization of Quercus suber MYB1, a transcription factor up-regulated in cork tissues. J. Plant Physiol. 2013, 170, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Almeida, T.; Pinto, G.; Correia, B.; Santos, C.; Goncalves, S. QsMYB1 expression is modulated in response to heat and drought stresses and during plant recovery in quercus suber. Plant Physiol. Biochem. 2013, 73, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Gocal, G.; Sheldon, C.; Gubler, F.; Moritz, T.; Bagnall, D.; MacMillan, C.; Li, S.; Parish, R.; Dennis, E.; Weigel, D.; et al. GaMyb-like genes, flowering, and gibberellin signaling in Arabidopsis. Plant Physiol. 2001, 127, 1682–1693. [Google Scholar] [CrossRef] [PubMed]
- Palatnik, J.; Allen, E.; Wu, X.; Schommer, C.; Schwab, R.; Carrington, J.; Weigel, D. Control of leaf morphogenesis by microRNAs. Nature 2003, 425, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Herr, A.; Baulcombe, D.; Harberd, N. Modulation of floral development by a gibberellin-regulated microRNA. Development 2004, 131, 3357–3365. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.; Chua, N. ABA induction of mir159 controls transcript levels of two Myb factors during Arabidopsis seed germination. Plant J. 2007, 49, 592–606. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, N.; Mi, X.; Wu, L.; Ma, R.; Zhu, X.; Yao, L.; Jin, X.; Si, H.; Wang, D. Identification of mir159s and their target genes and expression analysis under drought stress in potato. Comput. Biol. Chem. 2014, 53, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Garg, B.; Lata, C.; Prasad, M. A study of the role of gene TaMYB2 and an associated SNP in dehydration tolerance in common wheat. Mol. Biol. Rep. 2012, 39, 10865–10871. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, A.; Smita, S.; Lenka, S.K.; Rajwanshi, R.; Chinnusamy, V.; Bansal, K.C. Genome-wide classification and expression analysis of MYB transcription factor families in rice and Arabidopsis. BMC Genomics 2012, 13, 544. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Hirsch-Hoffmann, M.; Hennig, L.; Gruissem, W. Genevestigator Arabidopsis microarray database and analysis toolbox. Plant Physiol. 2004, 136, 2621–2632. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Gewin, V. Food: An underground revolution. Nature 2010, 466, 552–553. [Google Scholar] [CrossRef] [PubMed]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci. 2013, 4, 442. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, C.; Zhao, M.; Gonzalez, A.; Lloyd, A.; Schiefelbein, J. The bHLH genes GL3 and EGL3 participate in an intercellular regulatory circuit that controls cell patterning in the Arabidopsis root epidermis. Development 2005, 132, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Skirycz, A.; Inzé, D. More from less: Plant growth under limited water. Curr. Opin. Biotechnol. 2010, 21, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Sirichandra, C.; Wasilewska, A.; Vlad, F.; Valon, C.; Leung, J. The guard cell as a single-cell model towards understanding drought tolerance and abscisic acid action. J. Exp. Bot. 2009, 60, 1439–1463. [Google Scholar] [CrossRef] [PubMed]
- Bates, G.W.; Rosenthal, D.M.; Sun, J.; Chattopadhyay, M.; Peffer, E.; Yang, J.; Ort, D.R.; Jones, A.M. A comparative study of the Arabidopsis thaliana guard-cell transcriptome and its modulation by sucrose. PLoS ONE 2012, 7, e49641. [Google Scholar] [CrossRef] [PubMed]
- Cominelli, E.; Galbiati, M.; Tonelli, C. Transcription factors controlling stomatal movements and drought tolerance. Transcription 2010, 1, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.S.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef] [PubMed]
- Cominelli, E.; Galbiati, M.; Albertini, A.; Fornara, F.; Conti, L.; Coupland, G.; Tonelli, C. DOF-binding sites additively contribute to guard cell-specificity of AtMYB60 promoter. BMC Plant Biol. 2011, 11, 162. [Google Scholar] [CrossRef] [PubMed]
- Rusconi, F.; Simeoni, F.; Francia, P.; Cominelli, E.; Conti, L.; Riboni, M.; Simoni, L.; Martin, C.R.; Tonelli, C.; Galbiati, M. The Arabidopsis thaliana MYB60 promoter provides a tool for the spatio-temporal control of gene expression in stomatal guard cells. J. Exp. Bot. 2013, 64, 3361–3371. [Google Scholar] [CrossRef] [PubMed]
- Ito, M. Conservation and diversification of three-repeat MYB transcription factors in plants. J. Plant Res. 2005, 118, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Sheoran, I.; Saini, H. Drought-induced male sterility in rice: Changes in carbohydrate levels and enzyme activities associated with the inhibition of starch accumulation in pollen. Sex. Plant Reprod. 1996, 9, 161–169. [Google Scholar] [CrossRef]
- Lalonde, S.; Beebe, D.; Saini, H. Early signs of disruption of wheat anther development associated with the induction of male sterility by meiotic-stage water deficit. Sex. Plant Reprod. 1997, 10, 40–48. [Google Scholar] [CrossRef]
- Saini, H. Effects of water stress on male gametophyte development in plants. Sex. Plant Reprod. 1997, 10, 67–73. [Google Scholar] [CrossRef]
- Park, S.; Hwang, S.; Hauser, B. The phenotype of Arabidopsis ovule mutants mimics the morphology of primitive seed plants. Proc. R. Soc. B 2004, 271, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Hunt, K.; Hauser, B. Ovule abortion in Arabidopsis triggered by stress. Plant Physiol. 2004, 135, 2358–2367. [Google Scholar] [CrossRef] [PubMed]
- Mandaokar, A.; Browse, J. MYB108 acts together with MYB24 to regulate jasmonate-mediated stamen maturation in Arabidopsis. Plant Physiol. 2009, 149, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Putterill, J.; Laurie, R.; Macknight, R. It’s time to flower: The genetic control of flowering time. Bioessays 2004, 26, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2014, 5, 771. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Ye, Z.H. Secondary cell walls: Biosynthesis, patterned deposition and transcriptional regulation. Plant Cell Physiol. 2015, 56, 195–214. [Google Scholar] [CrossRef] [PubMed]
- Pollard, M.; Beisson, F.; Li, Y.; Ohlrogge, J.B. Building lipid barriers: Biosynthesis of cutin and suberin. Trends Plant Sci. 2008, 13, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Suh, M.C. Advances in the understanding of cuticular waxes in Arabidopsis thaliana and crop species. Plant Cell Rep. 2015, 34, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Jarvis, D.E.; Chen, H.; Beilstein, M.A.; Grimwood, J.; Jenkins, J.; Shu, S.; Prochnik, S.; Xin, M.; Ma, C.; et al. The reference genome of the halophytic plant Eutrema salsugineum. Front. Plant Sci. 2013, 4, 46. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Kasuga, M.; Liu, Q.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor. Nat. Biotechnol. 1999, 17, 287–291. [Google Scholar] [PubMed]
- Nakabayashi, R.; Saito, K. Integrated metabolomics for abiotic stress responses in plants. Curr. Opin. Plant Biol. 2015, 24C, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Zoratti, L.; Karppinen, K.; Luengo Escobar, A.; Häggman, H.; Jaakola, L. Light-controlled flavonoid biosynthesis in fruits. Front. Plant Sci. 2014, 5, 534. [Google Scholar] [CrossRef] [PubMed]
- Castellarin, S.D.; Pfeiffer, A.; Sivilotti, P.; Degan, M.; Peterlunger, E.; di Gaspero, G. Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit. Plant Cell Environ. 2007, 30, 1381–1399. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Achievements and challenges in understanding plant abiotic stress responses and tolerance. Plant Cell Physiol. 2011, 52, 1569–1582. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The transcriptional regulatory network in the drought response and its crosstalk in abiotic stress responses including drought, cold, and heat. Front. Plant Sci. 2014, 5, 170. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Schumaker, K.; Zhu, J. Molecular genetic perspectives on cross-talk and specificity in abiotic stress signalling in plants. J. Exp. Bot. 2004, 55, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, N.; Urwin, P. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Benkova, E. Cytokinin cross-talking during biotic and abiotic stress responses. Front. Plant Sci. 2013, 4, 451. [Google Scholar] [CrossRef] [PubMed]
- Urao, T.; Yamaguchi-Shinozaki, K.; Urao, S.; Shinozaki, K. An Arabidopsis MYB homolog is induced by dehydration stress and its gene product binds to the conserved MYB recognition sequence. Plant Cell 1993, 5, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- Bari, R.; Datt Pant, B.; Stitt, M.; Scheible, W.R. Pho2, microrna399, and phr1 define a phosphate-signaling pathway in plants. Plant Physiol. 2006, 141, 988–999. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Gan, S. AtMYB2 regulates whole plant senescence by inhibiting cytokinin-mediated branching at late stages of development in Arabidopsis. Plant Physiol. 2011, 156, 1612–1619. [Google Scholar] [CrossRef] [PubMed]
- Baek, D.; Park, H.C.; Kim, M.C.; Yun, D.J. The role of Arabidopsis MYB2 in mir399f-mediated phosphate-starvation response. Plant Signal. Behav. 2013, 8, e23488. [Google Scholar] [CrossRef] [PubMed]
- Gaff, D.F. Desiccation-tolerant flowering plants in Southern Africa. Science 1971, 174, 1033–1034. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Dey, A.; Gupta, B. Plant polyamines in abiotic stress responses. Acta Physiol. Plant 2013, 35, 2015–2036. [Google Scholar] [CrossRef]
- Jin, H.; Cominelli, E.; Bailey, P.; Parr, A.; Mehrtens, F.; Jones, J.; Tonelli, C.; Weisshaar, B.; Martin, C. Transcriptional repression by AtMYB4 controls production of UV-protecting sunscreens in Arabidopsis. EMBO J. 2000, 19, 6150–6161. [Google Scholar] [CrossRef] [PubMed]
- Persak, H.; Pitzschke, A. Dominant repression by Arabidopsis transcription factor MYB44 causes oxidative damage and hypersensitivity to abiotic stress. Int. J. Mol. Sci. 2014, 15, 2517–2537. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.; Jung, C.; Lee, S.; Min, K.; Lee, Y.; Choi, Y.; Lee, J.; Song, J.; Kim, J.; Choi, Y. AtMYB44 regulates WRKY70 expression and modulates antagonistic interaction between salicylic acid and jasmonic acid signaling. Plant J. 2013, 73, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Li, X.; Sun, W.; Li, L.; Gao, R.; Zhu, Q.; Tian, S.; Fu, M.; Yu, H.; Tang, X.; et al. AtMYB44 regulates resistance to the green peach aphid and diamondback moth by activating EIN2-affected defences in Arabidopsis. Plant Biol. 2013, 15, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, K.; Yamaguchi-Shinozaki, K.; Seki, M. Regulatory network of gene expression in the drought and cold stress responses. Curr. Opin. Plant Biol. 2003, 6, 410–417. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Kim, S.G.; Hong, S.; Park, C.M. Small interfering peptides as a novel way of transcriptional control. Plant Signal. Behav. 2008, 3, 615–617. [Google Scholar] [CrossRef] [PubMed]
- Matlin, A.; Clark, F.; Smith, C. Understanding alternative splicing: Towards a cellular code. Nat. Rev. Mol. Cell Biol. 2005, 6, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Syed, N.H.; Kalyna, M.; Marquez, Y.; Barta, A.; Brown, J.W. Alternative splicing in plants—Coming of age. Trends Plant Sci. 2012, 17, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, F.; Bracale, M.; Magaraggia, F.; Faoro, F.; Manzocchi, L.; Coraggio, I. The product of the rice MYB7 unspliced mRNA dimerizes with the maize leucine zipper Opaque2 and stimulates its activity in a transient expression assay. J. Biol. Chem. 2000, 275, 17619–17625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Li, X.; Guo, L.; Lu, F.; Feng, X.; He, K.; Wei, L.; Chen, Z.; Qu, L.; Gu, H. A subgroup of MYB transcription factor genes undergoes highly conserved alternative splicing in Arabidopsis and rice. J. Exp. Bot. 2006, 57, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Beers, E. Alternative splicing of Myb-related genes MYR1 and MYR2 may modulate activities through changes in dimerization, localization, or protein folding. Plant Signal. Behav. 2013, 8, e27325. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.; Dalmay, T.; Bartels, D. The role of small RNAs in abiotic stress. FEBS Lett. 2007, 581, 3592–3597. [Google Scholar] [CrossRef] [PubMed]
- Sunkar, R.; Li, Y.; Jagadeeswaran, G. Functions of microRNAs in plant stress responses. Trends Plant Sci. 2012, 17, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Tao, Y.; Zhu, C. Emerging roles of microRNAs in the mediation of drought stress response in plants. J. Exp. Bot. 2013, 64, 3077–3086. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, Q. MicroRNA-based biotechnology for plant improvement. J. Cell. Physiol. 2015, 230, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, H.; Aya, K.; Ueguchi-Tanaka, M.; Shimada, Y.; Nakazono, M.; Watanabe, R.; Nishizawa, N.; Gomi, K.; Shimada, A.; Kitano, H.; et al. GaMyb controls different sets of genes and is differentially regulated by microRNA in aleurone cells and anthers. Plant J. 2006, 47, 427–444. [Google Scholar] [CrossRef] [PubMed]
- Fahlgren, N.; Howell, M.; Kasschau, K.; Chapman, E.; Sullivan, C.; Cumbie, J.; Givan, S.; Law, T.; Grant, S.; Dangl, J.; et al. High-throughput sequencing of Arabidopsis microRNAs: Evidence for frequent birth and death of miRNA genes. PLoS ONE 2007, 2, e219. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.; Li, J.; Stahle, M.; Dubroue, A.; Gubler, F.; Millar, A. Genetic analysis reveals functional redundancy and the major target genes of the Arabidopsis mir159 family. Proc. Natl. Acad. Sci. USA 2007, 104, 16371–16376. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.; Gubler, F. The Arabidopsis GaMyb-like genes, MYB33 and MYB65, are microRNA-regulated genes that redundantly facilitate anther development. Plant Cell 2005, 17, 705–721. [Google Scholar] [CrossRef] [PubMed]
- Gubler, F.; Kalla, R.; Roberts, J.; Jacobsen, J. Gibberellin-regulated expression of a myb gene in barley aleurone cells—Evidence for Myb transactivation of a high-pl alpha-amylase gene promoter. Plant Cell 1995, 7, 1879–1891. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldoni, E.; Genga, A.; Cominelli, E. Plant MYB Transcription Factors: Their Role in Drought Response Mechanisms. Int. J. Mol. Sci. 2015, 16, 15811-15851. https://doi.org/10.3390/ijms160715811
Baldoni E, Genga A, Cominelli E. Plant MYB Transcription Factors: Their Role in Drought Response Mechanisms. International Journal of Molecular Sciences. 2015; 16(7):15811-15851. https://doi.org/10.3390/ijms160715811
Chicago/Turabian StyleBaldoni, Elena, Annamaria Genga, and Eleonora Cominelli. 2015. "Plant MYB Transcription Factors: Their Role in Drought Response Mechanisms" International Journal of Molecular Sciences 16, no. 7: 15811-15851. https://doi.org/10.3390/ijms160715811