
MicroRNA-944 Affects Cell Growth by Targeting EPHA7 in Non-Small Cell Lung Cancer

Abstract
:1. Introduction
2. Results
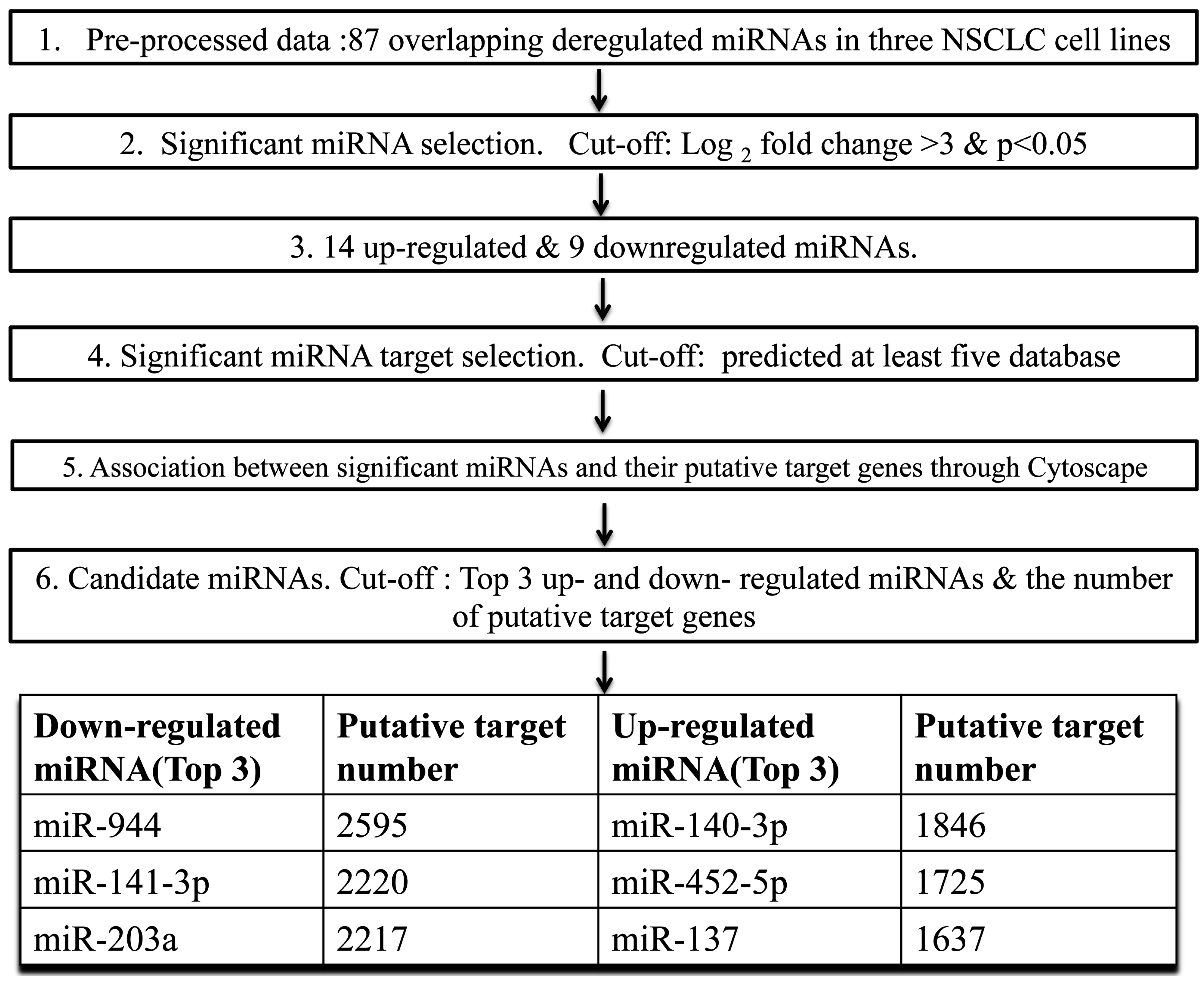
2.1. The Cancer-Related miRNAs Were Screened in Cultured NSCLC Cells
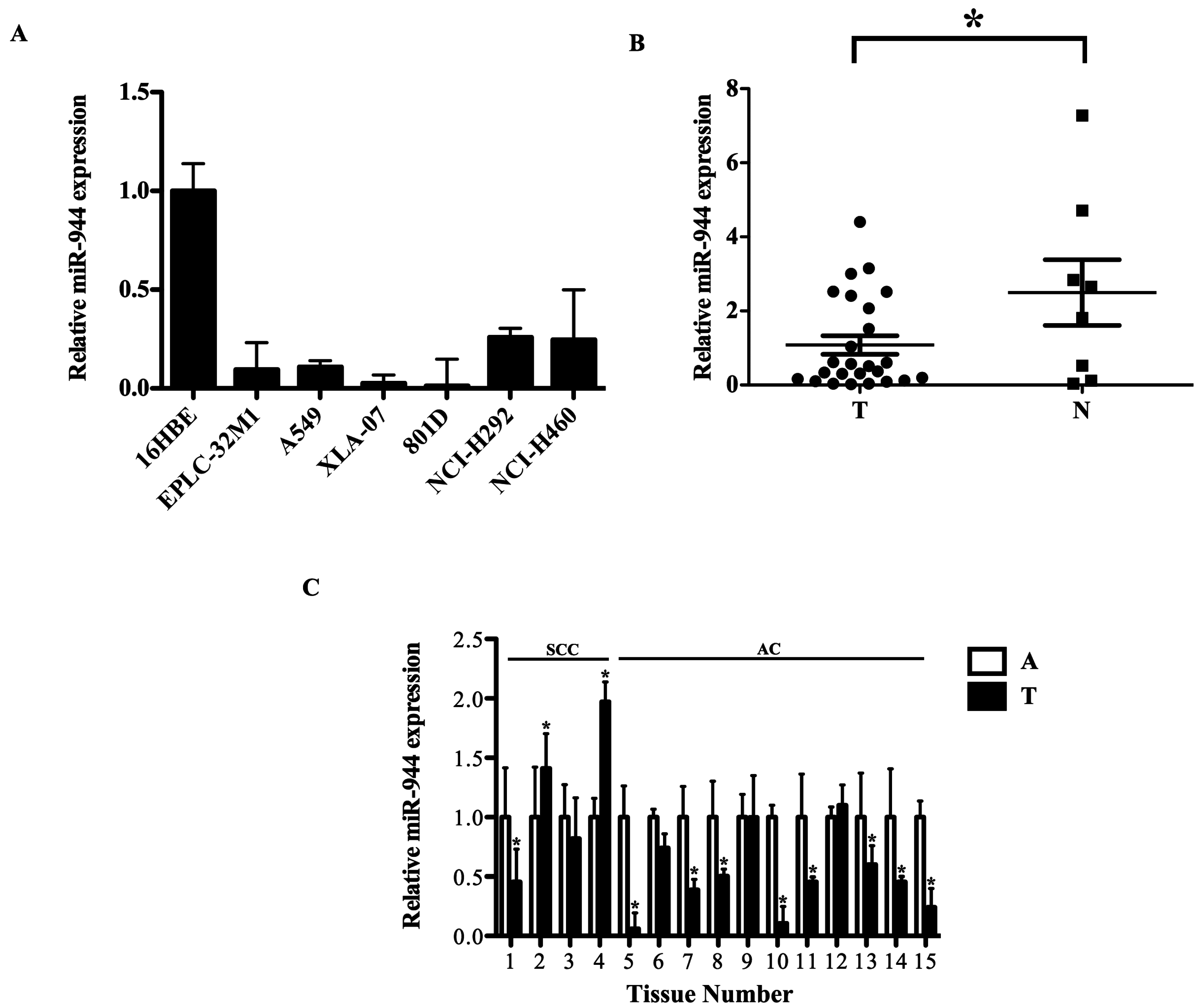
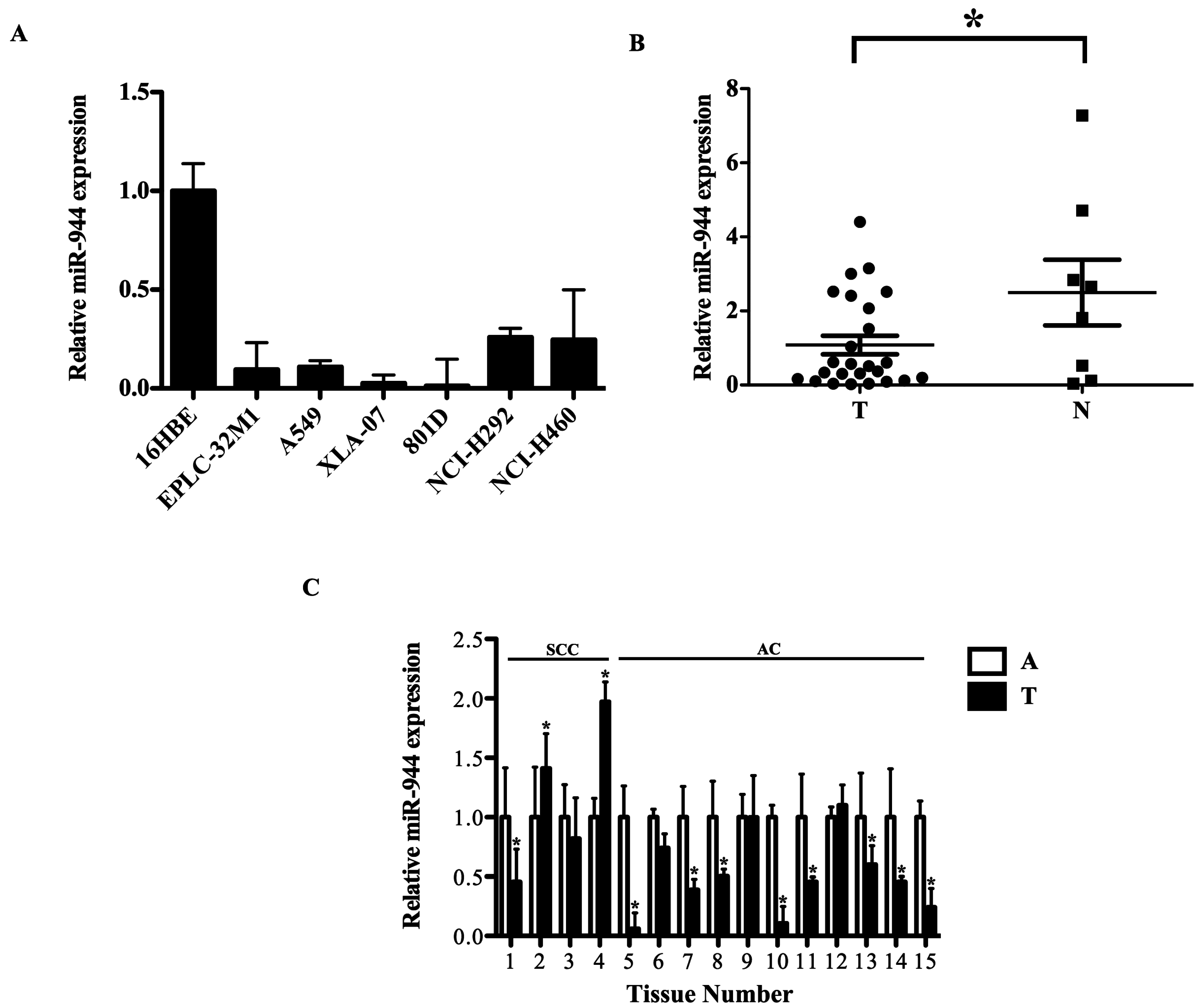
2.2. The miR-944 Expression Was Down-Regulated in NSCLC Tissues and Cell Lines
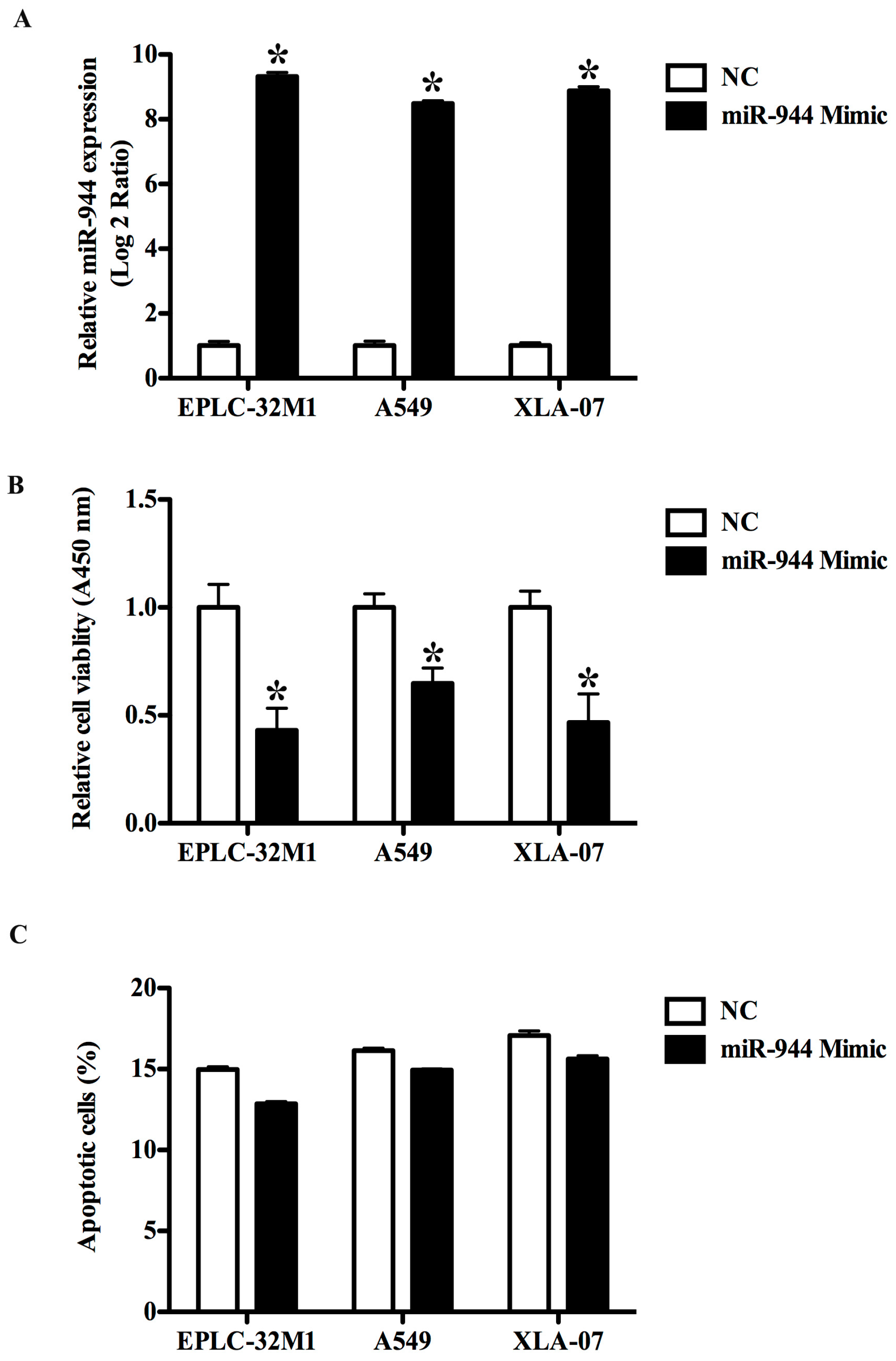
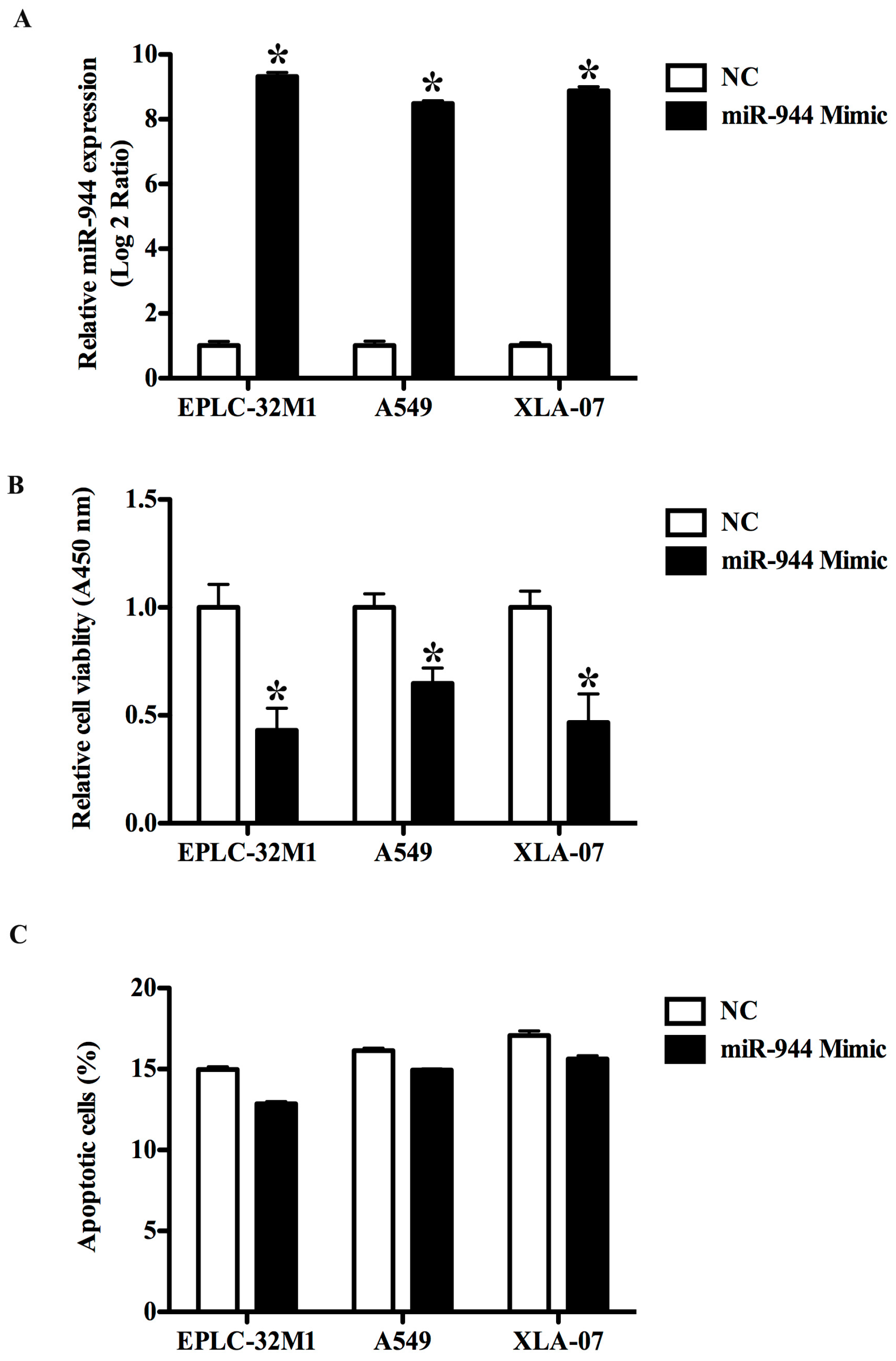
2.3. The Overexpression of miR-944 Could Suppress the Proliferation of NSCLC Cells
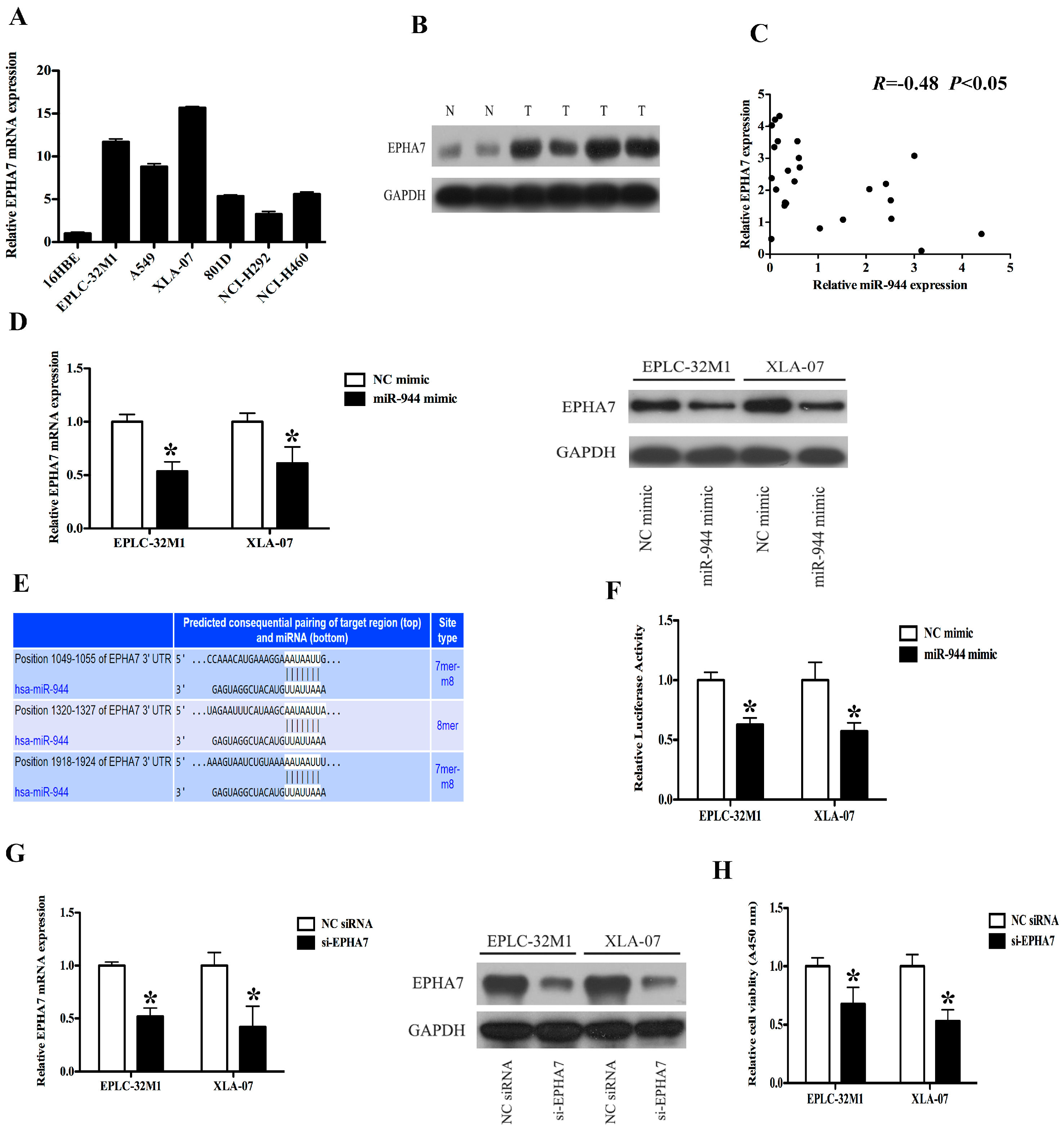
2.4. EPHA7 (a Target of miR-944) Was Screened in NSCLCs
2.5. The Expression Level of EPHA7 Is Inversely Correlated with the miR-944 Expression in NSCLC
2.6. The Up-Regulation of miR-944 Could Suppress the EPHA7 Expression
2.7. miR-944 Directly Targeted EPHA7 by Interacting with Its 3′-Untranslated Regions (UTRs)
2.8. The Down-Regulation of EPHA7 Could Suppress the Proliferation of NSCLC Cells
3. Discussion
4. Experimental Section
4.1. Human Tissue Specimens and Cell Lines
4.2. miRNA Expression Profiling
4.3. miRNA Target Prediction
4.4. Quantitative Real-Time PCR (qRT-PCR)
4.5. Oligonucleotide Transfection
4.6. MRNA Expression Profiling
4.7. Western Blot
4.8. Cell Growth and Apoptosis Assay
4.9. Luciferase Reporter Assays
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ettinger, D.S.; Akerley, W.; Borghaei, H.; Chang, A.C.; Cheney, R.T.; Chirieac, L.R.; D’Amico, T.A.; Demmy, T.L.; Govindan, R.; Grannis, F.W.; et al. Non-small cell lung cancer, version 2.2013. J. Natl. Compr. Canc. Netw. 2013, 11, 645–653. [Google Scholar] [PubMed]
- Molina, J.R.; Yang, P.; Cassivi, S.D.; Schild, S.E.; Adjei, A.A. Non-small cell lung cancer: Epidemiology, risk factors, treatment, and survivorship. Mayo Clin. Proc. 2008, 83, 584–594. [Google Scholar] [CrossRef]
- Sato, M.; Shames, D.S.; Gazdar, A.F.; Minna, J.D. A translational view of the molecular pathogenesis of lung cancer. J. Thorac. Oncol. 2007, 2, 327–343. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.-Y.; Yu, S.-L.; Yang, P.-C. MicroRNA in lung cancer. Br. J. Cancer 2010, 103, 1144–1148. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. Micrornas: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Ferracin, M.; Calin, G.A.; Negrini, M. MicroRNAs in cancer (an overview). In MicroRNAs in Cancer Translational Research; Springer: Berlin, Germany, 2011; pp. 1–71. [Google Scholar]
- Iorio, M.V.; Croce, C.M. MicroRNA dysregulation in cancer: Diagnostics, monitoring and therapeutics. EMBO Mol. Med. 2012, 4, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Creighton, C.J.; Reid, J.G.; Gunaratne, P.H. Expression profiling of microRNAs by deep sequencing. Brief. Bioinform. 2009, 10, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Markou, A.; Sourvinou, I.; Vorkas, P.A.; Yousef, G.M.; Lianidou, E. Clinical evaluation of microRNA expression profiling in non small cell lung cancer. Lung Cancer 2013, 81, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Cui, R.; Meng, W.; Sun, H.-L.; Kim, T.; Ye, Z.; Fassan, M.; Jeon, Y.-J.; Li, B.; Vicentini, C.; Peng, Y.; et al. MicroRNA-224 promotes tumor progression in nonsmall cell lung cancer. Proc. Natl. Acad. Sci. USA 2015, 112, E4288–E4297. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Sun, C.; Wang, S.; He, Q.; Li, D. MicroRNA miR-10b inhibition reduces cell proliferation and promotes apoptosis in non-small cell lung cancer (NSCLC) cells. Mol. Biosyst. 2015, 11, 2051–2059. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-X.; Zhou, K.-C.; Cao, Y. MCRS1 overexpression, which is specifically inhibited by miR-129*, promotes the epithelial-mesenchymal transition and metastasis in non-small cell lung cancer. Mol. Cancer 2014, 13, 245. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.-H.; Zhang, K.-D.; Duan, H.-X.; Liu, M.-X.; Wei, W.-L.; Cao, Y. ERGIC3, which is regulated by miR-203a, is a potential biomarker for non-small cell lung cancer. Cancer Sci. 2015, 106, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Nordentoft, I.; Birkenkamp-Demtroder, K.; Agerbæk, M.; Theodorescu, D.; Ostenfeld, M.S.; Hartmann, A.; Borre, M.; Ørntoft, T.F.; Dyrskjøt, L. miRNAs associated with chemo-sensitivity in cell lines and in advanced bladder cancer. BMC Med. Genom. 2012, 5, 40. [Google Scholar] [CrossRef] [PubMed]
- Schultz, N.A.; Andersen, K.K.; Roslind, A.; Willenbrock, H.; Wøjdemann, M.; Johansen, J.S. Prognostic MicroRNAs in Cancer Tissue from Patients Operated for Pancreatic Cancer—Five MicroRNAs in a Prognostic Index. World J. Surg. 2012, 36, 2699–2707. [Google Scholar] [CrossRef] [PubMed]
- Christensen, L.L.; Tobiasen, H.; Holm, A.; Schepeler, T.; Ostenfeld, M.S.; Thorsen, K.; Rasmussen, M.H.; Birkenkamp-Demtroeder, K.; Sieber, O.M.; Gibbs, P.; et al. miRNA-362–3p induces cell cycle arrest through targeting of E2F1, USF2 and PTPN1 and is associated with recurrence of colorectal cancer. Int. J. Cancer 2013, 133, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Mannoor, K.; Gao, L.; Tan, A.; Guarnera, M.A.; Zhan, M.; Shetty, A.; Stass, S.A.; Xing, L.; Jiang, F. Characterization of microRNA transcriptome in lung cancer by next-generation deep sequencing. Mol. Oncol. 2014, 8, 1208–1219. [Google Scholar] [CrossRef] [PubMed]
- Au, N.H.; Gown, A.M.; Cheang, M.; Huntsman, D.; Yorida, E.; Elliott, W.M.; Flint, J.; English, J.; Gilks, C.B.; Grimes, H.L. P63 expression in lung carcinoma: A tissue microarray study of 408 cases. Appl. Immunohistochem. Mol. Morphol. 2004, 12, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Bishop, J.A.; Teruya-Feldstein, J.; Westra, W.H.; Pelosi, G.; Travis, W.D.; Rekhtman, N. P40 (ΔNp63) is superior to p63 for the diagnosis of pulmonary squamous cell carcinoma. Mod. Pathol. 2012, 25, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.P.; Lau, N.C.; Garrett-Engele, P.; Grimson, A.; Schelter, J.M.; Castle, J.; Bartel, D.P.; Linsley, P.S.; Johnson, J.M. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 2005, 433, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Giaginis, C.; Tsoukalas, N.; Bournakis, E.; Alexandrou, P.; Kavantzas, N.; Patsouris, E.; Theocharis, S. Ephrin (Eph) receptor A1, A4, A5 and A7 expression in human non-small cell lung carcinoma: Associations with clinicopathological parameters, tumor proliferative capacity and patients’ survival. BMC Clin. Pathol. 2014, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Kataoka, H.; Suzuki, M.; Sato, N.; Nakamura, R.; Tao, H.; Maruyama, K.; Isogaki, J.; Kanaoka, S.; Ihara, M.; et al. Downregulation of EphA7 by hypermethylation in colorectal cancer. Oncogene 2005, 24, 5637–5647. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, G.; Ma, H.; Bao, Y.; Wang, X.; Zhou, H.; Sheng, Z.; Sugimura, H.; Jin, J.; Zhou, X. Differential expression of EphA7 receptor tyrosine kinase in gastric carcinoma. Hum. Pathol. 2007, 38, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- Guan, M.; Xu, C.; Zhang, F.; Ye, C. Aberrant methylation of EphA7 in human prostate cancer and its relation to clinicopathologic features. Int. J. Cancer 2009, 124, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Oricchio, E.; Nanjangud, G.; Wolfe, A.L.; Schatz, J.H.; Mavrakis, K.J.; Jiang, M.; Liu, X.; Bruno, J.; Heguy, A.; Olshen, A.B. The Eph-receptor A7 is a soluble tumor suppressor for follicular lymphoma. Cell 2011, 147, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Xiang, C.; Lv, Y.; Wei, Y.; Wei, J.; Miao, S.; Mao, X.; Gu, X.; Song, K.; Jia, S. Effect of EphA7 Silencing on Proliferation, Invasion and Apoptosis in Human Laryngeal Cancer Cell Lines Hep-2 and AMC-HN-8. Cell Physiol. Biochem. 2015, 36, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-F.; Fokas, E.; Juricko, J.; You, A.; Rose, F.; Pagenstecher, A.; Engenhart-Cabillic, R.; An, H.-X. Increased expression of EphA7 correlates with adverse outcome in primary and recurrent glioblastoma multiforme patients. BMC Cancer 2008, 8, 79. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhou, K.; Huang, Y.; Cao, Y. The candidate oncogene (MCRS1) promotes the growth of human lung cancer cells via the miR–155–Rb1 pathway. J. Exp. Clin. Cancer Res. 2015, 34, 121. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Wang, H.; Bian, L.; Shao, W.; Tang, R.; Wang, Q.; Jin, K. Establishment and characterization of lung adenocarcinoma cell line XLA-07. Zhonghua Bing Li Xue Za Zhi 2012, 41, 335–339. [Google Scholar] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Log2(Fold Change) | Cytoband | ||
|---|---|---|---|---|
| EPLC-32M1 vs. 16HBE | 801D vs. 16HBE | A549 vs. 16HBE | ||
| Up-Regulated miRNAs | ||||
| hsa-miR-10a-3p | 4.446 | 6.715 | 4.289 | 17q21.32 |
| hsa-miR-10a-5p | 5.816 | 8.581 | 4.626 | 17q21.32 |
| hsa-miR-10b-5p | 8.912 | 6.686 | 8.627 | 17q21.32 |
| hsa-miR-137 | 8.775 | 9.571 | 8.524 | 1p22 |
| hsa-miR-140-3p | 4.144 | 4.659 | 3.415 | 16q22.1 |
| hsa-miR-193a-5p | 4.103 | 6.348 | 4.054 | 17q11.2 |
| hsa-miR-224-5p | 13.982 | 9.493 | 12.369 | Xq28 |
| hsa-miR-2682-5p | 8.822 | 10.672 | 8.441 | Chromosome1: (98510798-98510907) |
| hsa-miR-31-3p | 3.050 | 4.505 | 5.289 | 9p21.3 |
| hsa-miR-3179 | 7.568 | 8.893 | 7.469 | 16p13.11 |
| hsa-miR-320d | 5.557 | 3.675 | 3.404 | 13q14.11 |
| hsa-miR-365b-5p | 5.031 | 7.127 | 6.003 | 17q11.2 |
| hsa-miR-452-5p | 13.175 | 7.857 | 10.770 | Xq28 |
| hsa-miR-935 | 7.900 | 10.634 | 7.664 | 19q13.42 |
| Down-Regulated miRNAs | ||||
| hsa-miR-129-1-3p | 9.110 | 3.802 | 6.340 | 7q32 |
| hsa-miR-141-3p | 6.884 | 11.752 | 7.398 | 12p13.31 |
| hsa-miR-200c-3p | 7.068 | 9.662 | 7.691 | 12p13.31 |
| hsa-miR-203a | 4.007 | 3.724 | 9.962 | 14q32.33 |
| hsa-miR-205-3p | 6.766 | 6.766 | 6.766 | 1q32.2 |
| hsa-miR-544b | 6.650 | 6.650 | 6.6501 | Chromosome3: (124451286-124451363) |
| hsa-miR-934 | 8.313 | 8.313 | 8.313 | Xq26.3 |
| hsa-miR-936 | 7.650 | 7.650 | 7.650 | 10q25.1 |
| hsa-miR-944 | 3.597 | 7.235 | 3.4975 | 3q28 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Zhou, K.; Cao, Y. MicroRNA-944 Affects Cell Growth by Targeting EPHA7 in Non-Small Cell Lung Cancer. Int. J. Mol. Sci. 2016, 17, 1493. https://doi.org/10.3390/ijms17101493
Liu M, Zhou K, Cao Y. MicroRNA-944 Affects Cell Growth by Targeting EPHA7 in Non-Small Cell Lung Cancer. International Journal of Molecular Sciences. 2016; 17(10):1493. https://doi.org/10.3390/ijms17101493
Chicago/Turabian StyleLiu, Minxia, Kecheng Zhou, and Yi Cao. 2016. "MicroRNA-944 Affects Cell Growth by Targeting EPHA7 in Non-Small Cell Lung Cancer" International Journal of Molecular Sciences 17, no. 10: 1493. https://doi.org/10.3390/ijms17101493