
Waste Soybean Oil and Corn Steep Liquor as Economic Substrates for Bioemulsifier and Biodiesel Production by Candida lipolytica UCP 0998

Abstract
:
1. Introduction
2. Results
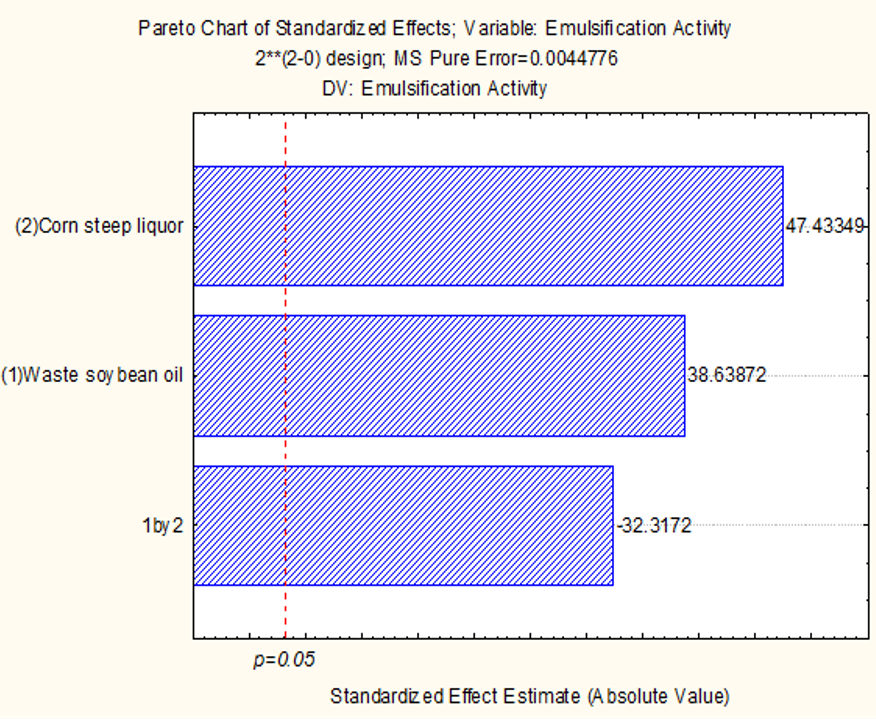
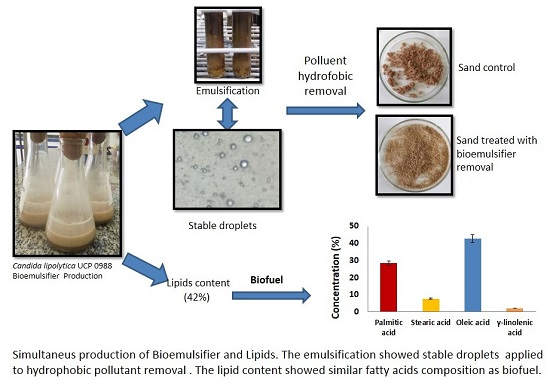
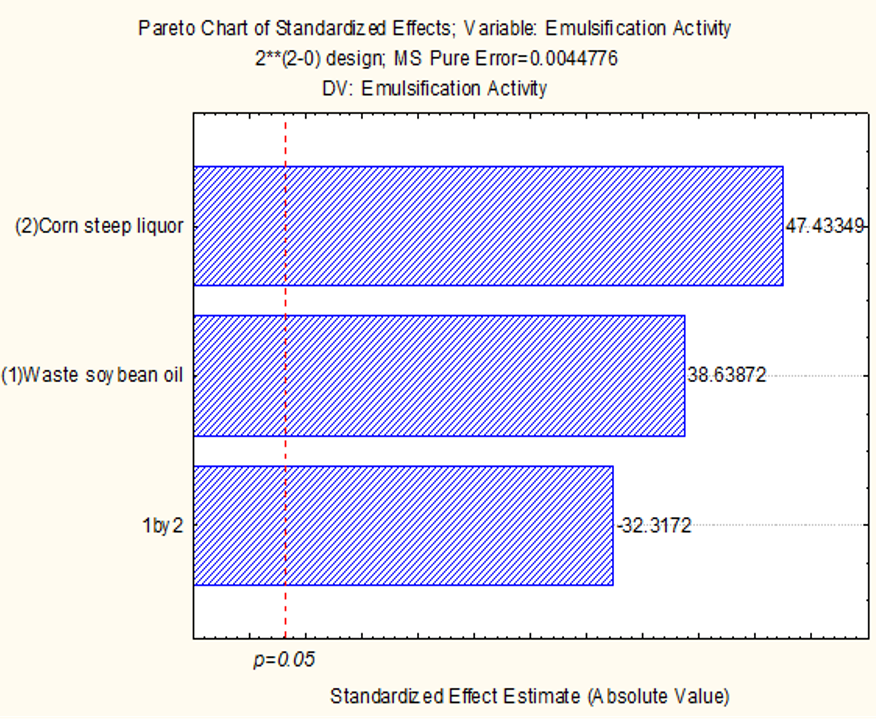
2.1. Production of Bio-Emulsifier by Candida lipolytica UCP 0988 Using Agro-Industrial Waste
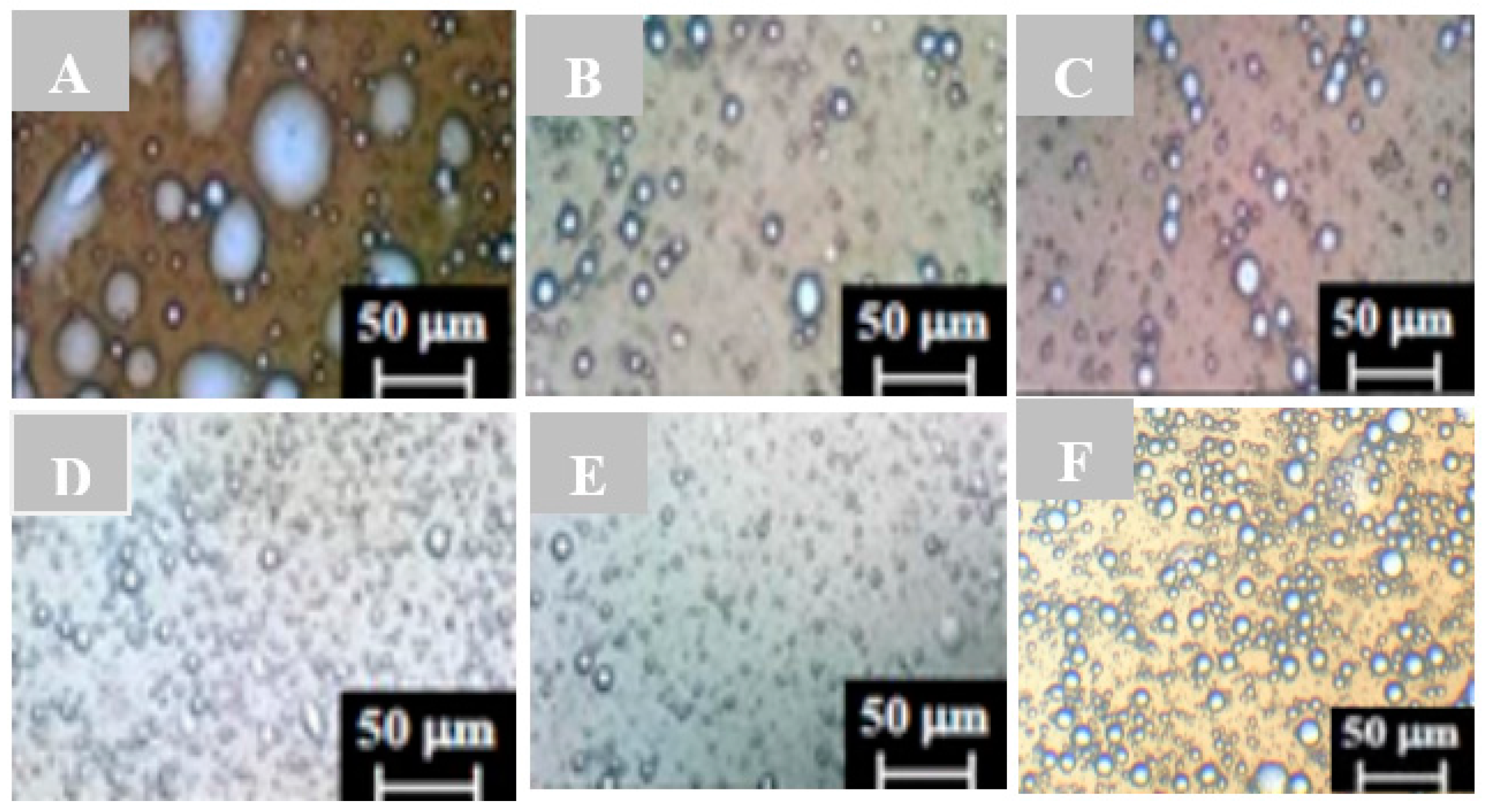
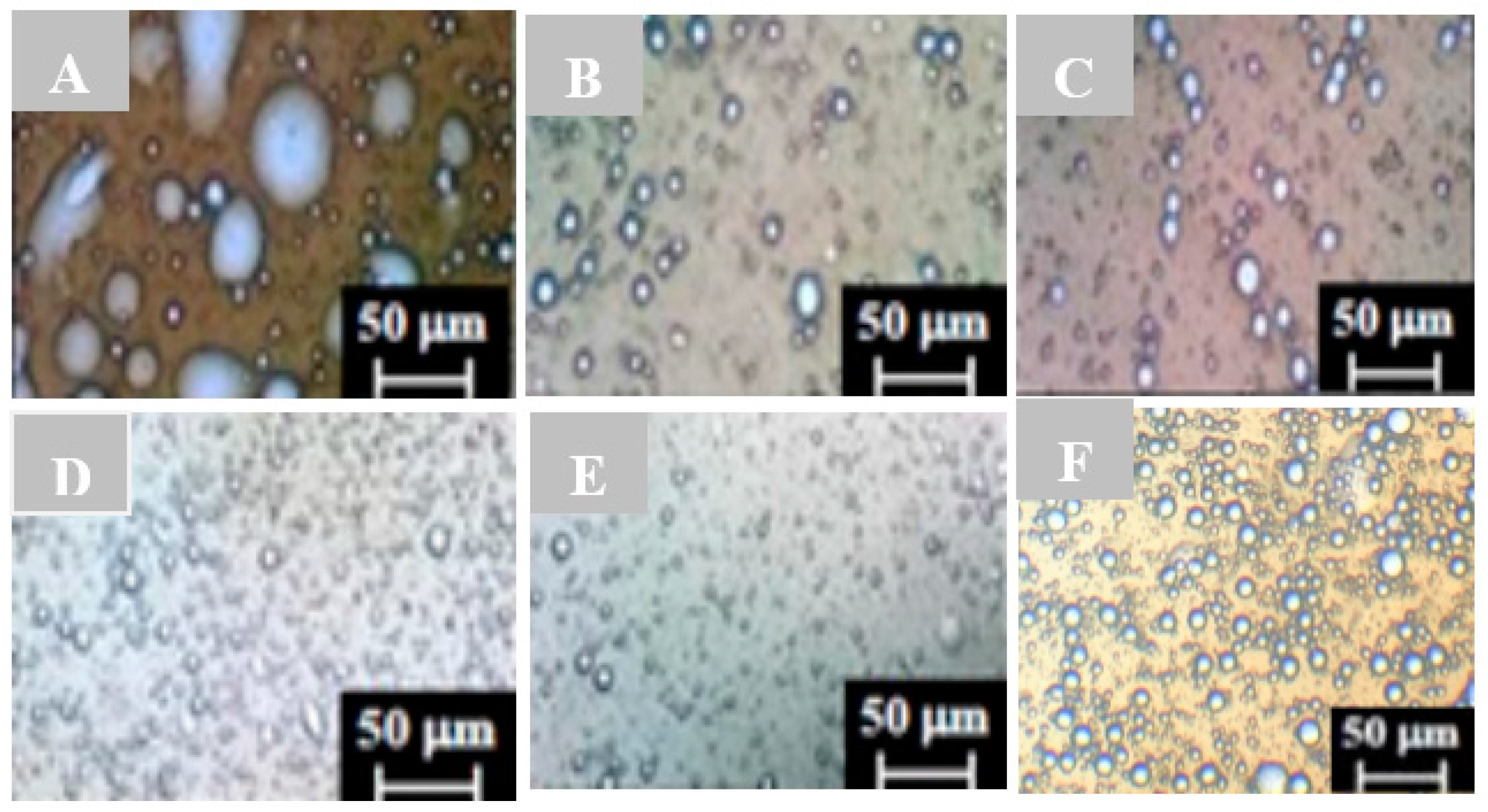
2.2. Characterization of Emulsion Droplets by Light Microscopy
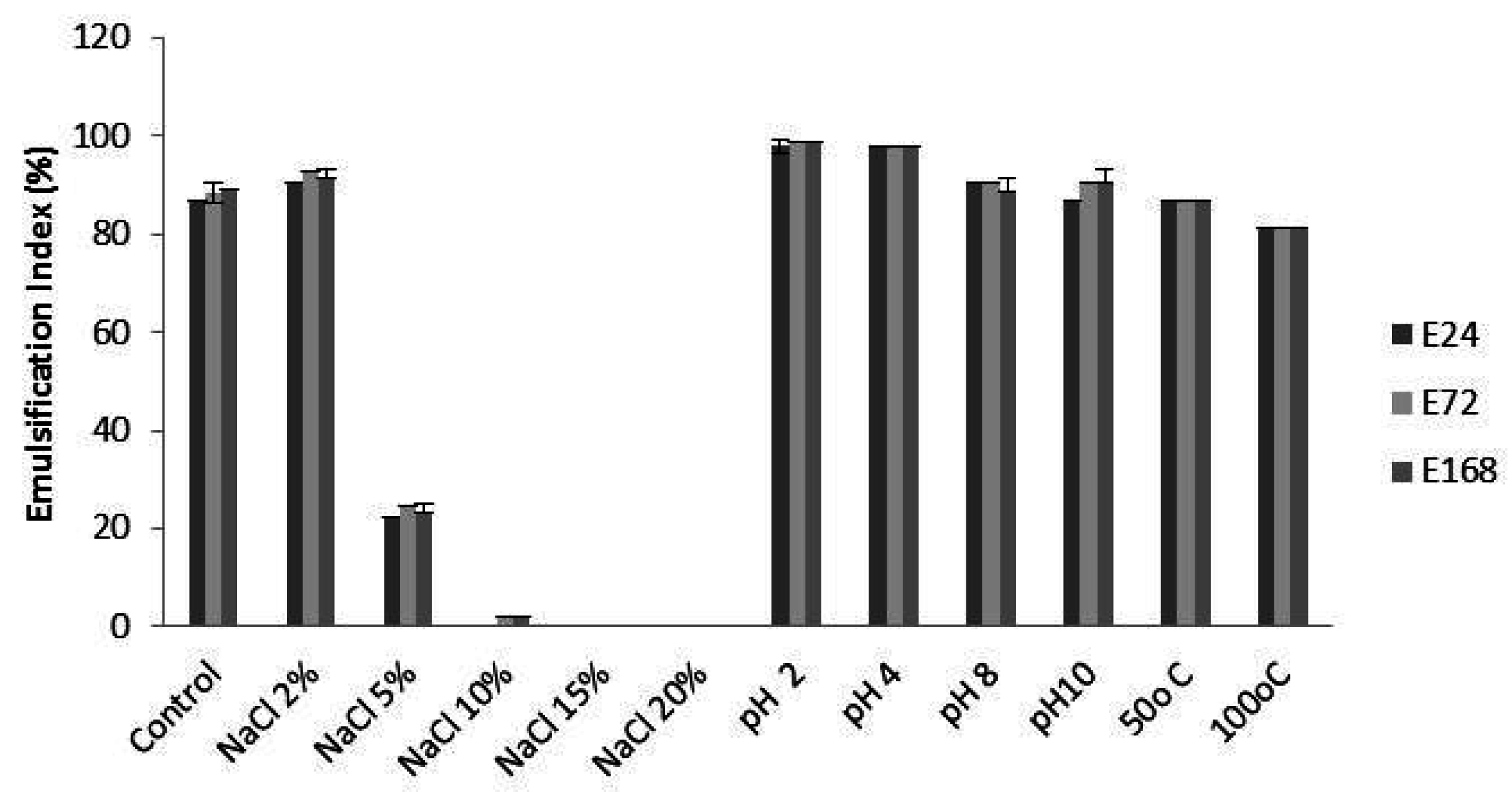
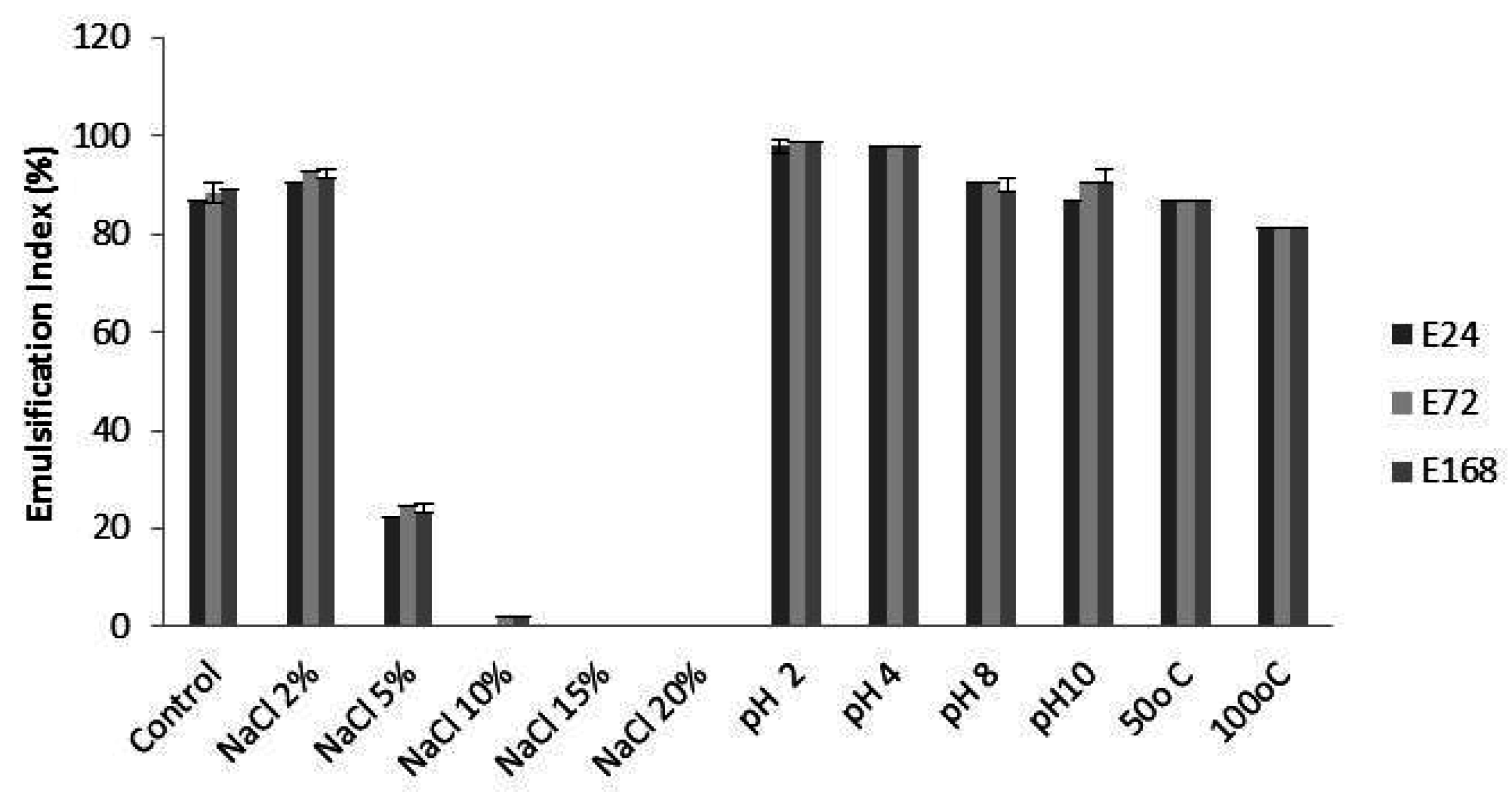
2.3. Stability of Bioemulsifier
2.4. Potential of Bioemulsifier on Displacement of Hydrophobic Compound in Seawater
2.5. Application of Bioemulsifier to Remove a Hydrophobic Compound
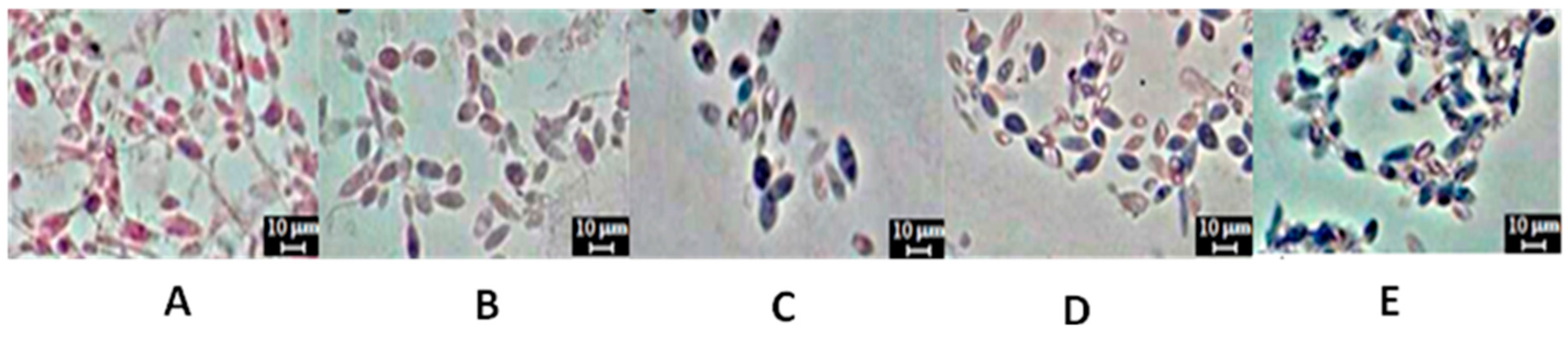
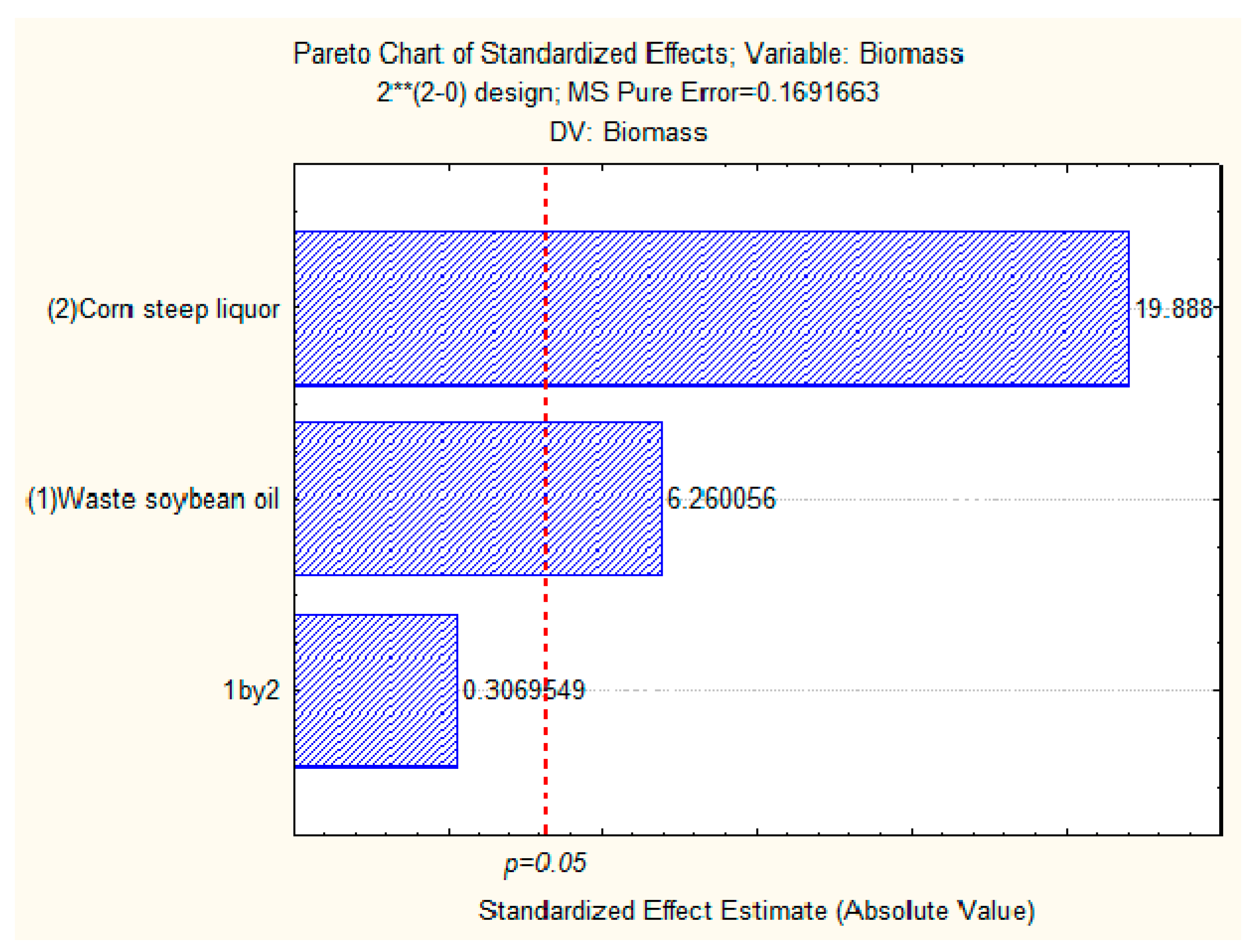
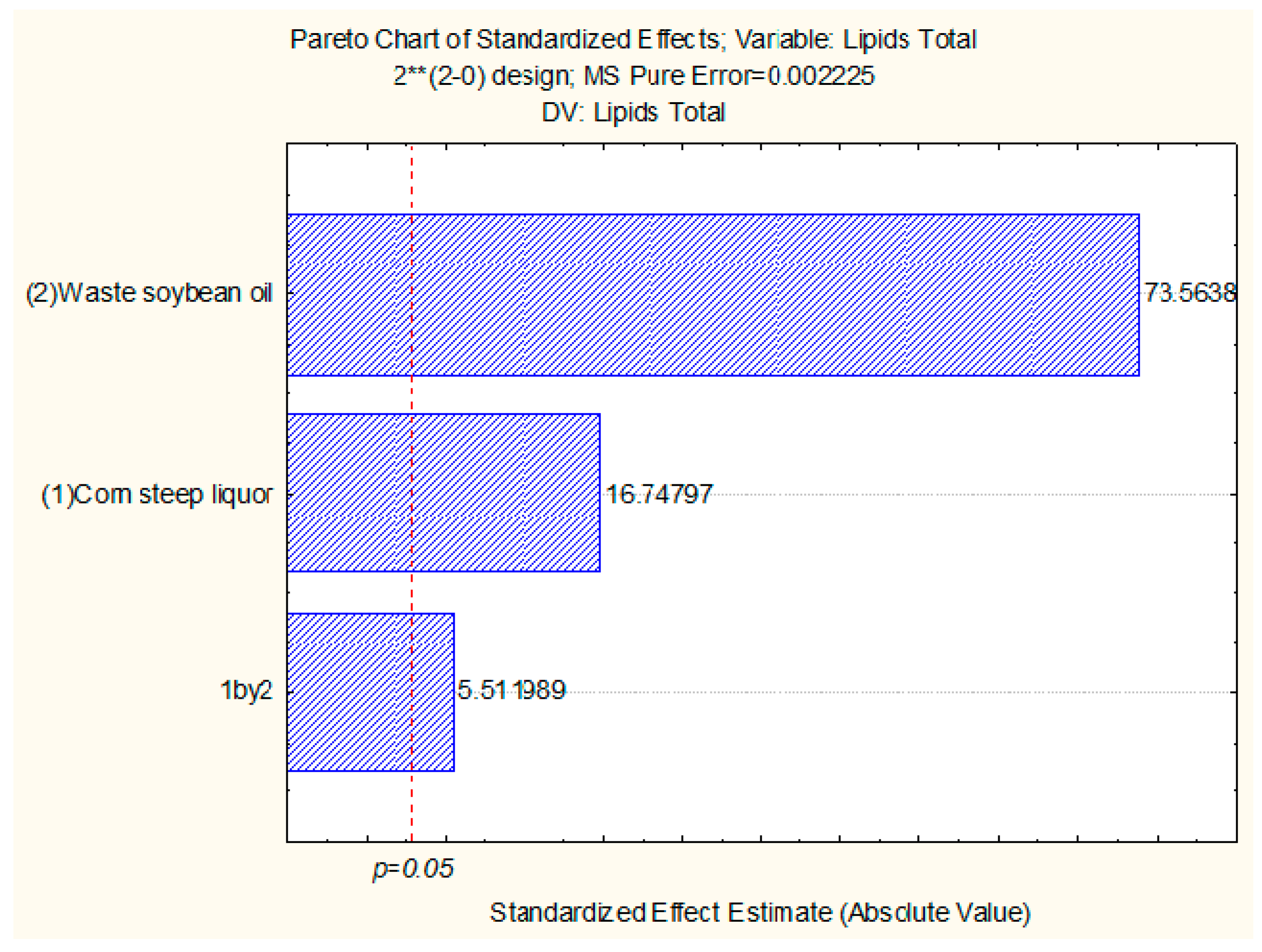
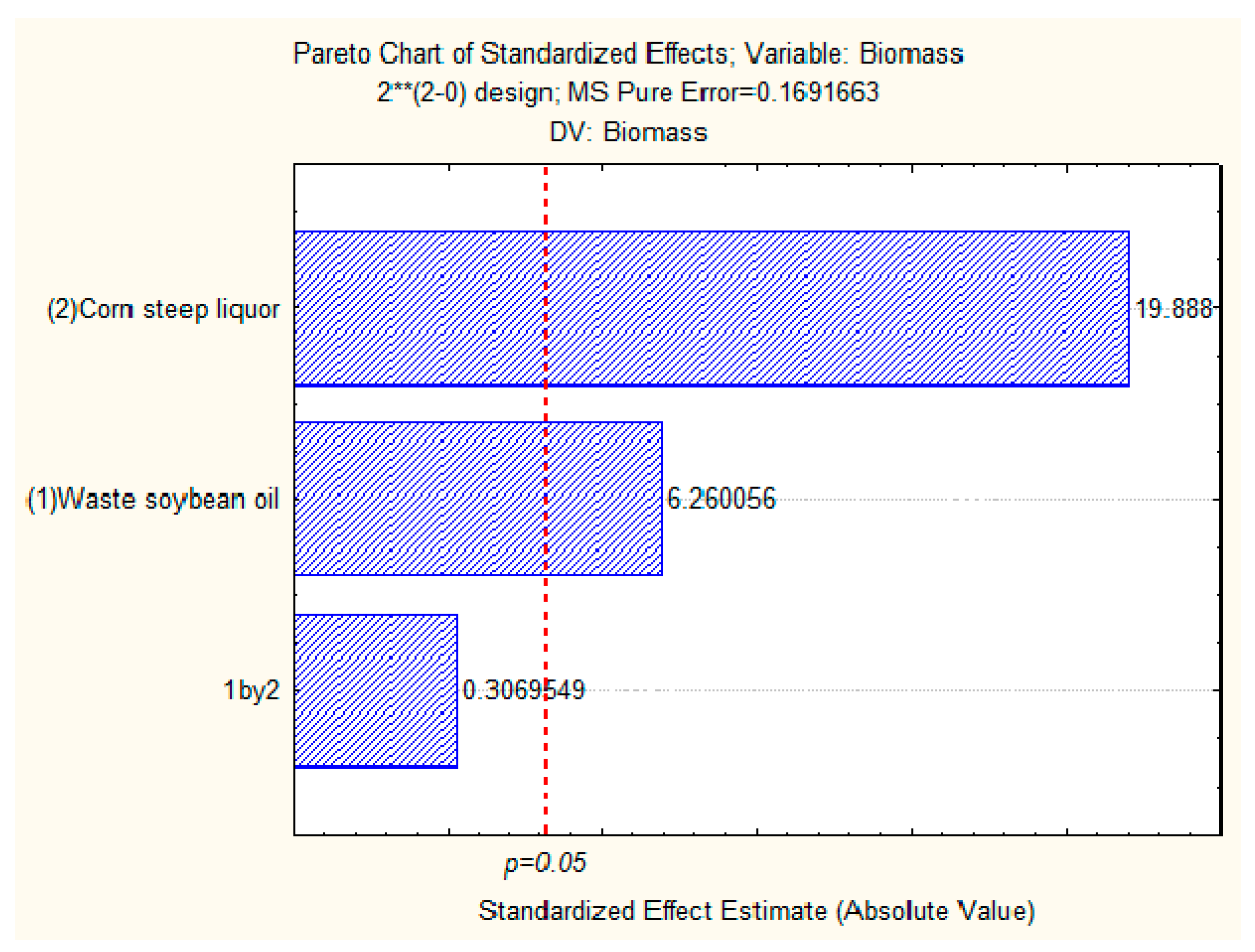
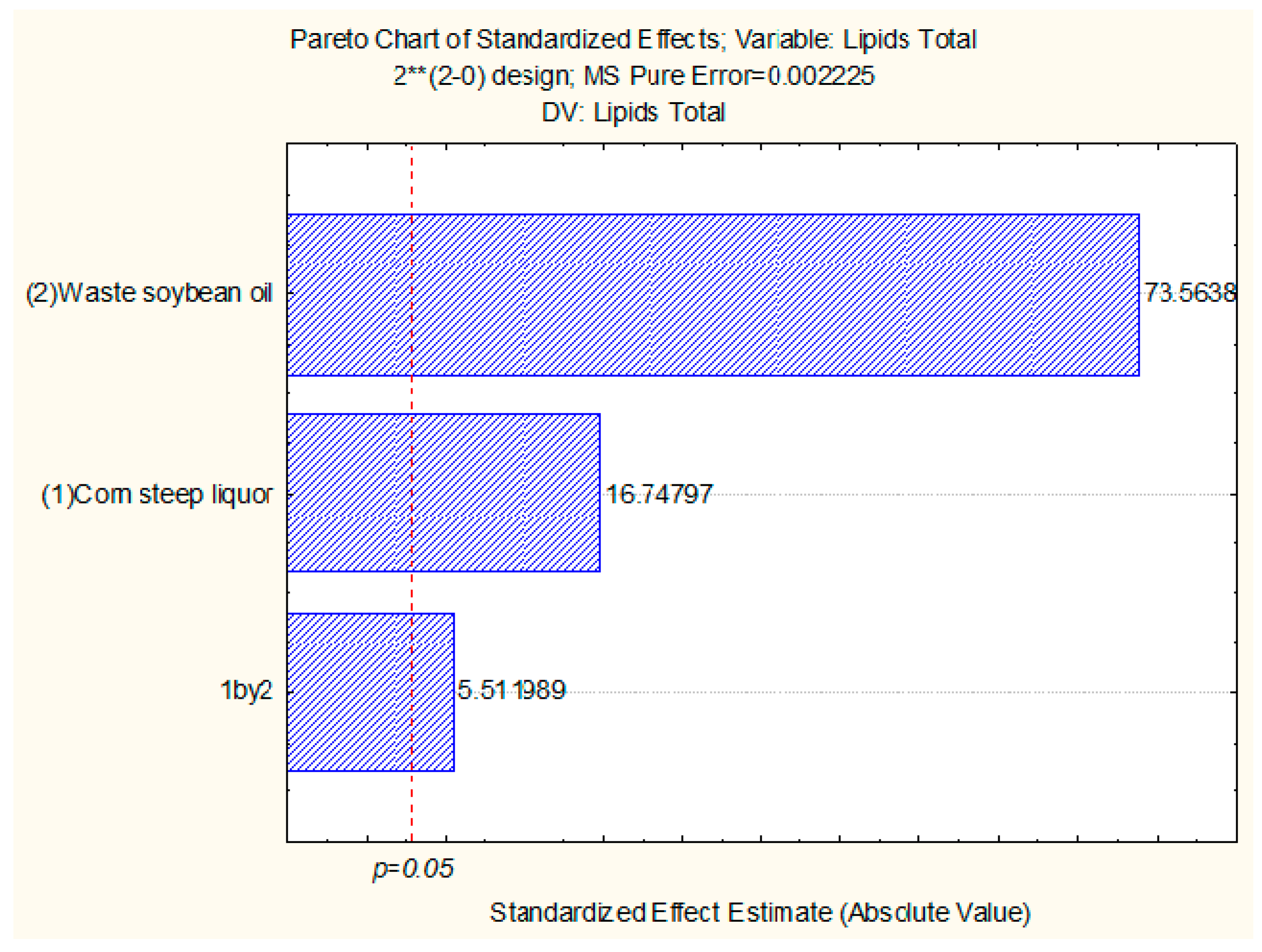
2.6. Production of Biomass and Total Lipids by Candida lipolytica UCP 0988 Using Agro-Industrial Waste
2.7. Fatty Acid Profile of Total Lipids
2.8. Transesterification/Direct Esterification of Biomass
3. Discussion
3.1. Production of Bioemulsifier by Candida lipolytica UCP 0988 Using Agro-Industrial Waste
3.2. Characterizing Droplets with Regard to Stable Emulsions
3.3. Stability of Bioemulsifier
3.4. Potential of Bioemulsifier on Displacement of Hydrophobic Compound in Seawater
3.5. Application of Bioemulsifier to Remove a Hydrophobic Compound
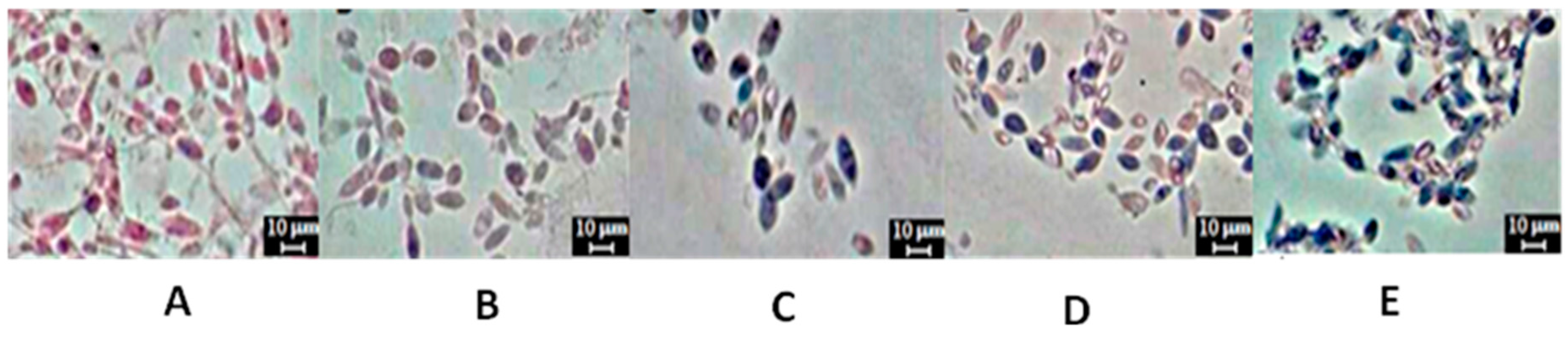
3.6. Production of Biomass and Total Lipids by Candida lipolytica UCP 0988 Using Agro-Industrial Waste
3.7. Fatty Acid Profile of Total Lipids
3.8. Transesterification/Direct Esterification of Biomass
4. Materials and Methods
4.1. Microorganism
4.2. Substrates
4.3. Growth Conditions
4.4. Determining the Production of Bioemulsifier
4.5. Isolating the Bioemulsifier
4.6. Characterizing the Bioemulsifier
4.7. Determining the Ionic Character of the Bioemulsifier
4.8. Measuring the Droplets
4.9. Oil Spreading Test
4.10. Stability Studies
4.11. Applying Bioemulsifier to Remove Diesel Oil from Contaminated Sand
4.12. Determining the Biomass
4.13. Cytochemical Analysis of Lipids
4.14. Lipid Extraction
4.15. Identifying Fatty Acids from Biomass
4.16. Transesterification/Esterification
4.17. Factorial Design
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Janek, T.; Marcin, Ł.; Anna, K. Antiadhesive activity of the biosurfactant pseudofactin II secreted by the Arctic bacterium Pseudomonas fluorescens BD5. BMC Microbial. 2012, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Uzoigwe, C.; Burgess, J.G.; Ennis, C.J.; Rahman, P.K. Bioemulsifiers are not biosurfactants and require different screening approaches. Front. Microbiol. 2015, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sekhon-Randhawa, K.K. Biosurfactants Produced by Genetically Manipulated Micro-Organisms: Challenges and Opportunities; Kosaric, N., Sukan, F.V., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 49–67. [Google Scholar]
- Franzetti, A.; Gandolfi, I.; Raimondi, C.; Bestetti, G.; Banat, I.M.; Smyth, T.J.; Fracchia, L. Environmental fate, toxicity, characteristics and potential applications of novel bioemulsifiers produced by Variovorax paradoxus 7bCT5. Biol. Technnol. 2012, 108, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Beopoulos, A.; Nicaud, J.M.; Gaillardin, C. An overview of lipid metabolism in yeasts and its impact on biotechnological processes. Appl. Microbial. Biotechnol. 2011, 4, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Cirigliano, M.C.; Carman, G.M. Isolation of a bioemulsifier from Candida lipolytica. Appl. Environ. Microb. 1984, 48, 747–750. [Google Scholar]
- Amaral, P.F.F.; da Silva, J.M.; Lehocky, M.; Barros-Timmons, A.M.V.; Coelho, M.A.Z.; Marrucho, I.M.; Coutinho, J.A.P. Production and characterization of a bioemulsifier from Yarrowia lipolytica. Process Biochem. 2006, 41, 1894–1898. [Google Scholar] [CrossRef]
- Rufino, R.D.; Luna, J.M.; Campos-Takaki, G.M.C.; Sarubbo, L. A Characterization and properties of the biosurfactant produced by Candida lipolytica UCP 0988. Electron. J. Biotechnnol. 2014, 17, 34–38. [Google Scholar] [CrossRef]
- Rossi, M.; Amaretti, A.; Raimondi, S.; Leonardi, A. Getting lipids for biodiesel production from oleaginous fungi. Biodiesel Feedstocks Process. Technol. 2011, 1, 72–74. [Google Scholar]
- Sitepu, I.; Selby, T.; Lin, T.; Zhu, S.; Boundy-Mills, K. Carbon source utilization and inhibitor tolerance of 45 oleaginous yeast species. J. Ind. Microbiol. Biotechnol. 2014, 41, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Phillips, G.O.; Williams, P.A. Introduction to food proteins. In Handbook of Food Proteins; Phillips, G.O.O., Williams, P.A., Eds.; Woodhead Publishing Ltd.: Cambridge, UK, 2001; pp. 1–12. [Google Scholar]
- Dickinson, E. Hydrocolloids as emulsifiers and emulsion stabilizers. Food Hydrocoll. 2009, 23, 1473–1482. [Google Scholar] [CrossRef]
- McClements, D.J. Food Emulsions; CRC Press: Boca Raton, FL, USA, 1999; pp. 111–124. [Google Scholar]
- Khot, M.; Kamat, S.; Zinjarde, S.; Pant, A.; Chopade, B.; RaviKumar, A. Single cell oil of oleaginous fungi from the tropical mangrove wetlands as a potential feedstock for biodiesel. Microb. Cell Factories 2012, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Willumsen, P.A.; Karlson, U. Screening of bacteria, isolated from PAH-contaminated soils, for production of biosurfactants and bioemulsifiers. Biodegradation 1996, 7, 415–423. [Google Scholar] [CrossRef]
- Rodrigues, L.R.; Teixeira, J.A.; van der Mei, H.C.; Oliveira, R. Physicochemical and functional characterization of a biosurfactant produced by Lactococcus lactis 53. Colloid Surf. B Biointerfaces 2006, 49, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobrinho, H.B.; Rufino, R.D.; Luna, J.M.; Salgueiro, A.A.; Campos-Takaki, G.M.; Leite, L.F.; Sarubbo, L.A. Utilization of two agro-industrial by-products for the production of a surfactant by Candida sphaerica UCP0995. Process Biochem. 2008, 49, 912–917. [Google Scholar] [CrossRef]
- Luna, J.M.; Rufino, R.D.; Sarubbo, L.A.; Rodrigues, L.R.; Teixeira, J.A.; Campos-Takaki, G.M. Evaluation antimicrobial and anti-adhesive properties of the biosurfactant Lunasan produced by Candida sphaerica UCP 0995. Curr. Microbial. 2011, 62, 1527–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, D.K.; Rufino, R.D.; Luna, J.M.; Santos, V.A.; Salgueiro, A.A.; Sarubbo, L.A. Synthesis and evaluation of biosurfactant produced by Candida lipolytica using animal fat and corn steep liquor. J. Petrol. Sci. Eng. 2013, 105, 43–50. [Google Scholar] [CrossRef]
- Antunes, A.A.; Silva, M.L.R.B.; Silva, C.A.A.; Campos-Takaki, G.M. Characterization of Chromobacterium violaceum isolated from Paca River, Pernambuco, Brazil. Rev. Biol. Ciencias Terra 2006, 1, 48–55. [Google Scholar]
- Albuquerque, C.D.C.; Campos-Takaki, G.M.; Fileti, A.M.F. Neural network based software sensors: Application to biosurfactant production by Candida lipolytica. In Modern Multidisciplinary Applied Microbiology: Exploiting Microbes and Their Interactions; Mendes-Vilas, A., Ed.; Wiley & Sons: New York, NY, USA, 2006; pp. 628–632. [Google Scholar]
- Cameotra, S.S.; Makkar, R.S. Synthesis of biosurfactants in extreme conditions. Appl. Microbiol. Biotechnol. 1998, 50, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Langevin, D.; Poteau, S.; Hénaut, I.; Argillier, J.F. Crude oil emulsion properties and their application to heavy oil transportation. Oil Gas Sci. Technol. 2004, 59, 511–521. [Google Scholar] [CrossRef]
- Camacho-Chab, J.C.; Guézennec, J.; Chan-Bacab, M.J.; Ríos-Leal, E.; Sinquin, C.; Muñiz-Salazar, R.; Rosa-García, S.D.; Reyes-Estebanez, M.; Ortega-Morales, O. Emulsifying Activity and Stability of a Non-Toxic Bioemulsifier Synthesized by Microbacterium sp. MC3B-10. Int. J. Mol. Sci. 2013, 14, 18959–18972. [Google Scholar] [CrossRef] [PubMed]
- Gudiña, E.O.J.; Pereira, J.F.B.; Costa, R.; Evtuguin, D.V.; Coutinho, J.A.P.; Teixeira, J.A.; Rodrigues, L.A.R. Novel bioemulsifier produced by a Paenibacillus strain isolated from crude oil. Microb. Cell Factories 2015, 14, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jillavenkatesa, A.; Dapkunas, S.J.; Lum, L.S.H. NIST Recommended Practice Guide: Particle Size Characterization; NIST Special Publication: Gaithersburg, Maryland, 2001; p. 164. [Google Scholar]
- Bungenberg de Jong, H.G.; Kruyt, H. R, Coacervation (Partial Miscibility in colloid systems). Proc. Koninkl. Med. Akad. Wetersnap. 1929, 32, 849–856. [Google Scholar]
- Thavasi, R.; Jayalakshmi, S.; Banat, I.M. Application of biosurfactant produced from peanut oil cake by Lactobacillus delbrueckii in biodegradation of crude oil. Bioresour. Technol. 2011, 102, 3366–3372. [Google Scholar] [CrossRef] [PubMed]
- Poomtien, J.; Thaniyavarn, J.; Pinphanichakarn, P.; Jindamorakot, S.; Morikawa, M. Production and Characterization of a Biosurfactant from Cyberlindnera samutprakarnensis JP52T. Biosci. Biotechnol. Biochem. 2013, 77, 2362–2370. [Google Scholar] [CrossRef] [PubMed]
- Andrade Silva, N.R.; Luna, M.A.; Santiago, A.L.; Franco, L.O.; Silva, G.K.; Souza, P.M.; Campos-Takaki, G.M. Biosurfactant-and-bioemulsifier produced by a promising Cunninghamella echinulata isolated from caatinga soil in the northeast of Brazil. Int. J. Mol. Sci. 2014, 15, 15377–15395. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, C.N.; Yong, R.N.; Gibbs, B.F. Surfactant-enhanced remediation of contaminated soil. Eng. Geol. 2001, 60, 371–380. [Google Scholar] [CrossRef]
- Rufino, R.D.; Sarubbo, L.A.; Neto, B.B.; Campos-Takaki, G.M. Experimental design for the production of tensio-active agent by Candida lipolytica. J. Ind. Microbiol. Biotechnol. 2008, 35, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Andrade, R.F.; Antunes, A.A.; Lima, R.A.; Araújo, H.W.; Resende-Stoianoff, M.A.; Franco, L.O.; Campos-Takaki, G.M. Enhanced production of a glycolipid biosurfactant produced by Candida glabrata UCP/WFCC1556 for application in dispersion and removal of petroderivatives. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 563–576. [Google Scholar]
- Urum, K.; Pekdemir, T.; Çopur, M. Surfactants treatment of crude oil contaminated soils. J. Colloid Interface Sci. 2004, 276, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Bharucha, C.; Jha, S.; Yadav, S.; Nerurkar, A.; Desai, A.J. Biosurfactant production using molasses and whey under thermophilic conditions. Bioresour. Technol. 2008, 99, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Daverey, A.; Pakshirajan, K. Sophorolipids from Candida bombicola using mixed hydrophilic substrates: production, purification and characterization. Colloids Surf. B Biointerfaces 2010, 79, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Beopoulos, A.; Chardot, T.; Nicaud, J.M. Yarrowia lipolytica: A model and a tool to understand the mechanisms implicated in lipid accumulation. Biochimie 2009, 91, 692–696. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Chevalot, I.; Komaitis, M.; Marc, I.; Aggelis, G. Single cell oil production by Yarrowia lipolytica growing on an industrial derivative of animal fat in batch cultures. Appl. Microbiol. Biotechnol. 2002, 58, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Areesirisuk, A.; Chiu, C.H.; Yen, T.B.; Liu, C.H.; Guo, J.H. A novel oleaginous yeast strain with high lipid productivity and its application to alternative biodiesel production. Appl. Biochem. Microbiol. 2015, 51, 411–418. [Google Scholar] [CrossRef]
- Karatay, S.E.; Dönmez, G. Improving the lipid accumulation properties of the yeast cells for biodiesel production using molasses. Bioresour. Technol. 2010, 101, 7988–7990. [Google Scholar] [CrossRef] [PubMed]
- Saenge, C.; Cheirsilp, B.; Suksaroge, T.T.; Bourtoom, T. Potential use of oleaginous red yeast Rhodotorula glutinis for the bioconversion of crude glycerol from biodiesel plant to lipids and carotenoids. Process Biochem. 2011, 46, 210–218. [Google Scholar] [CrossRef]
- Kitcha, S.; Benjamas, C. Screening of oleaginous yeasts and optimization for lipid production using crude glycerol as a carbon source. Energy Procedia 2011, 9, 274–282. [Google Scholar] [CrossRef]
- Galafassi, S.; Cucchetti, D.; Pizza, F.; Franzosi, G.; Bianchi, D.; Compagno, C. Lipid production for second generation biodiesel by the oleaginous yeast Rhodotorula graminis. Bioresour. Technol. 2012, 111, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Shen, H.; Zhou, W.; Wang, Y.; Yang, X.; Zhao, Z.K. Efficient conversion of acetate into lipids by the oleaginous yeast Cryptococcus curvatus. Biotechnol. Biofuels 2015, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Burdon, K.L. Fatty material in bacteria and fungi revealed by staining dried, fixed slide preparations. J. Bacteriol. 1946, 52, 665–678. [Google Scholar] [PubMed]
- Lakshmi, P.D.; Chakrapani, R.; Rao, C.S.V.R. Production of poly(3-hydroxybutyrates) by Bacillus species isolated from marine. Int. J. Chem. Sci. 2011, 9, 1963–1972. [Google Scholar]
- Neema, P.M.; Kumari, A. Isolation of lipid producing yeast and fungi from secondary sewage sludge and soil. Aust. J. Basic Appl. Sci. 2013, 7, 283–288. [Google Scholar]
- André, A.; Chatzifragkou, A.; Diamantopoulou, P.; Sarris, D.; Philippoussis, A.; Galiotou-Panayotou, M.; Papanikolaou, S. Biotechnological conversions of bio-diesel-derived crude glycerol by Yarrowia lipolytica strains. Eng. Life Sci. 2009, 9, 468–478. [Google Scholar] [CrossRef]
- Bati, N.; Hammond, E.G.; Glatz, B.A. Biomodification of fats and oils: Trials with Candida lipolytica. J. Am. Oil Chem. Soc. 1984, 61, 1743–1746. [Google Scholar] [CrossRef]
- El Bialy, H.; Gomaa, O.M.; Azab, K.S. Conversion of oil waste to valuable fatty acids using oleaginous yeast. World J. Microbiol. Biotechnol. 2011, 27, 2791–2798. [Google Scholar] [CrossRef]
- Islam, M.A.; Ayoko, G.A.; Brown, R.; Stuart, D.; Heimann, K. Influence of fatty acid structure on fuel properties of algae derived biodiesel. Procedia Eng. 2013, 56, 591–596. [Google Scholar] [CrossRef]
- Knothe, G. Designer biodiesel: Optimizing fatty ester composition to improve fuel properties. Energy Fuels 2008, 22, 1358–1364. [Google Scholar] [CrossRef]
- Gerpen, J. Biodiesel processing and production. Fuel Proc. Technol. 2005, 86, 1097–1107. [Google Scholar] [CrossRef]
- Ramos, M.J.; Fernández, C.M.; Casas, A.; Rodríguez, L.; Pérez, A. Influence of fatty acid composition of raw materials on biodiesel properties. Bioresour. Technol. 2009, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Vicente, G.; Bautista, L.F.; Gutiérrez, F.J.; Rodríguez, R.; Martínez, V.; Rodríguez-Frómeta, R.A.; Garre, V. Direct transformation of fungal biomass from submerged cultures into biodiesel. Energy Fuels 2010, 24, 3173–3178. [Google Scholar] [CrossRef]
- Marchetti, J.M.; Miguel, V.U.; Errazu, A.F. Possible methods for biodiesel production. Renew. Sustain. Energy Rev. 2007, 11, 1300–1311. [Google Scholar] [CrossRef]
- Fakas, S.; Papanikolaou, S.; Batsos, A.; Galiotou-Panayotou, M.; Mallouchos, A.; Aggelis, G. Evaluating renewable carbon sources as substrates for single cell oil production by Cunninghamella echinulata and Mortierella isabellina. Biomass Bioenergy 2009, 33, 573–580. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation on the purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Cooper, B.G. Surface active agents from two Bacillus species. Appl. Environ. Microbiol. 1987, 54, 224–229. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 29, 350–356. [Google Scholar] [CrossRef]
- Manocha, M.S.; San-Blas, G.; Centeno, S. Lipid composition of Paracoccidioides brasiliensis: Possible correlation with virulence of different strains. Microbiology 1980, 117, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Techaoei, S.; Lumyong, S.; Prathumpai, W.; Santiarwarn, D.; Leelapornpisid, P. Screening characterization and stability of biosurfactant produced by Pseudomonas aeruginosa SCMU106 isolated from soil in northern Thailand. Asian J. Biol. Sci. 2011, 3, 340–351. [Google Scholar] [CrossRef]
- Sheehan, H.L.; Storey, G.W. An improved method of staining leukocyte granules with Sudan Black B. J. Pathol. Bacteriol. 1947, 59, 336. [Google Scholar] [CrossRef] [PubMed]
- Durham, D.R.; Kloos, W.E. Comparative study of the total cellular fatty acids of Staphylococcus species of human origin. Int. J. Syst. Bacteriol. 1978, 28, 223–228. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assays | Emulsification Index (%) | Emulsification Activity (UEA) | ||
|---|---|---|---|---|
| Burned Engine Oil | n-Hexadecane | Corn Oil | n-Hexadecane | |
| 1 | 64.00 | Negative | 35.00 | 0.945 |
| 2 | 91.40 | 14.70 | 50.00 | 5.693 |
| 3 | 93.33 | 60.00 | 55.00 | 6.281 |
| 4 | 96.66 | 63.63 | 58.00 | 6.704 |
| 5 | 66.66 | Negative | 50.00 | 4.233 |
| 6 | 70.96 | Negative | 47.00 | 4.335 |
| 7 | 72.85 | Negative | 50.00 | 4.370 |
| 8 | 74.28 | Negative | 50.00 | 4.379 |
| Time of Growth (h) | Droplet Size Measurement (µm) | Emulsification Index (E24 (%)) | Units of Emulsification Activity (UEA) | Emulsification Properties |
|---|---|---|---|---|
| Liquid Medium (Control) | 1–30 | Not formed | 0.018 | No emulsification. Drops without globular form and large size. Droplets unstable. |
| 24 | 0.5–10 | 28.57 | 0.891 | Turbid with sedimentation, globular droplets and large flocculation. Droplets unstable. |
| 48 | 0.3–10 | 47.06 | 5.648 | Homogeneous with smaller and large oil droplets. Droplets unstable. |
| 72 | 0.1–10 | 76.47 | 5.812 | Droplets were homogeneous, smaller and stable. |
| 96 | 0.1–5 | 96.66 | 6.101 | Droplets were homogeneous, compact, smaller and stable. |
| SDS (Control) | 2.5–12 | 100 | 6.933 | Droplets were homogeneous, compact smaller and stable. |
| Assays | pH | Biomass (g/L) | Lipids (g/L Biomass) | Lipids (g/g Biomass) |
|---|---|---|---|---|
| 1 | 4.1 | 1.96 | 0.696 | 0.355 |
| 2 | 4.2 | 4.41 | 1.87 | 0.425 |
| 3 | 4.1 | 10.02 | 3.15 | 0.314 |
| 4 | 4.2 | 12.71 | 4.96 | 0.390 |
| 5 | 4.1 | 6.71 | 2.17 | 0.323 |
| 6 | 4.1 | 6.25 | 2.25 | 0.360 |
| 7 | 4.2 | 6.73 | 2.47 | 0.367 |
| 8 | 4.2 | 7.16 | 2.20 | 0.307 |
| Fatty Acid Methyl Esters (%) | |
|---|---|
| Capric acid (C10:0) | – |
| Myristic acid (C14:0) | – |
| Palmitic acid (C16:0) | 28.4 |
| Palmitoleic acid (C16:1) | – |
| Stearic acid (C18:0) | 7.7 |
| Oleic acid (C18:1) | 42.8 |
| Linoleic acid (C18:2) | 19.0 |
| α-Linolenic acid (C18:3–ω-3) | – |
| γ-Linolenic acid (C18:3–ω-6) | 2.1 |
| Arachidic acid (C20:0) | – |
| Gadoleic acid (C20:1) | – |
| Behenic acid (C22:0) | – |
| Microorganism | Carbon Source (Substrates) | Lipids (%) | References |
|---|---|---|---|
| Candida lipolytica | Waste soybean oil (8%) | 42.5 | Present study |
| Candida lipolytica | Molasses at 8% | 59.9 | Karatay and Donmez [40] |
| Yarrowia lipolytica | Industrial glycerol at 50% | 43.0 | Papanikolaou and Aggelis [38] |
| Cryptococcus curvatus | Acetate (15.9%) | 73.4 | Gong et al. [44] |
| Rhodotorula glutinis | Crude glycerol 10% and Tween 20 at 1% | 35.2 | Saenge et al. [41] |
| Rhodotorula graminis | Corn steep solids 2% and Yeast extract (0.1%) | 52.1 | Galafassi et al. [43] |
| Trichosporonoides spathulata | Crude glycerol at 10% | 40.6 | Kitcha and Cheirsilp [42] |
| Pseudozyma parantarctica | Glucose (40%) | 43.5 | Areesirisuk et al. [39] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, A.F.; Rodriguez, D.M.; Ribeaux, D.R.; Luna, M.A.C.; Lima e Silva, T.A.; Andrade, R.F.S.; Gusmão, N.B.; Campos-Takaki, G.M. Waste Soybean Oil and Corn Steep Liquor as Economic Substrates for Bioemulsifier and Biodiesel Production by Candida lipolytica UCP 0998. Int. J. Mol. Sci. 2016, 17, 1608. https://doi.org/10.3390/ijms17101608
Souza AF, Rodriguez DM, Ribeaux DR, Luna MAC, Lima e Silva TA, Andrade RFS, Gusmão NB, Campos-Takaki GM. Waste Soybean Oil and Corn Steep Liquor as Economic Substrates for Bioemulsifier and Biodiesel Production by Candida lipolytica UCP 0998. International Journal of Molecular Sciences. 2016; 17(10):1608. https://doi.org/10.3390/ijms17101608
Chicago/Turabian StyleSouza, Adriana Ferreira, Dayana M. Rodriguez, Daylin R. Ribeaux, Marcos A. C. Luna, Thayse A. Lima e Silva, Rosileide F. Silva Andrade, Norma B. Gusmão, and Galba M. Campos-Takaki. 2016. "Waste Soybean Oil and Corn Steep Liquor as Economic Substrates for Bioemulsifier and Biodiesel Production by Candida lipolytica UCP 0998" International Journal of Molecular Sciences 17, no. 10: 1608. https://doi.org/10.3390/ijms17101608